
Abstract
Three Bacillus cereus strains isolated from an outbreak of food poisoning caused by the consumption of fermented black beans (douchi) containing B. cereus is described. The outbreak involved 139 persons who had nausea, vomiting, and diarrhea. The strains were isolated from vomit and the unprepared douchi. Two of the strains produced the emetic toxin cereulide, as evidenced by polymerase chain reaction analysis for the presence of the nonribosomal synthetase cluster responsible for the synthesis of cereulide and by chemical analysis by high-performance liquid chromatography–mass spectrometry. These two strains belong to genetic group III of B. cereus, and multiple locus sequence typing revealed that the type was ST26, as a major part of B. cereus emetic strains. One of these strains produced significantly more cereulide at 37°C than the type cereulide producer (F4810/72), and it was also able to produce the toxin at 40°C and 42°C. The third strain belongs to genetic group IV, and it is a new multiple locus sequence type closely related to strains that are cytotoxic and enterotoxigenic. It possesses genes for hemolysin BL, nonhemolytic enterotoxin, and cytotoxin K2; however, it varies from the majority of strains possessing genes for hemolysin BL by not being hemolytic. Thus, two B. cereus strains producing the emetic toxin cereulide and a strain producing enterotoxins might have been involved in this food-poisoning incident caused by the consumption of a natural fermented food. The ability of one of the strains to produce cereulide at ≥37°C makes it possible that it is produced in the human gut in addition to occurring in the food.
Introduction
B
The aim of this article is to characterize two emetic and one enterotoxic B. cereus strains that were involved in an outbreak causing emesis and diarrhea due to the consumption of Chinese fermented black beans, douchi. Douchi is used to flavor fish or stir-fried vegetables in the Chinese kitchen. It is produced by two fermentation processes. The process starts with washing, soaking, steaming, and cooling of the black beans, and inoculation with fungi (Aspergillus oryzae, or Mucor spp.) or bacteria (Bacillus subtilis) and the first fermentation. This is followed by addition of salts including ferrosulfate, and a second natural anaerobic fermentation for 27 d, after which the product is dried. The process and the product are similar to African fermented bean products such as ogiri and iru (Zhao, 2003).
Materials and Methods
Three B. cereus strains (KM007-1, KM007-2, and KM008) were isolated during an outbreak. They originated from a vomit sample (KM007-1; KM007-2) and from a douchi sample (KM008) (Table 1). The B. cereus strains were isolated on mannitol–egg yolk–polymyxin agar after homogenization of 25 g vomit or food samples in 0.9% saline (National Standards of the People's Republic of China for examination of B. cereus [GB/T 4789.14-2003]). They were identified by glucose anaerobic fermentation, nitrate reduction, gelatin liquefaction, mobility, β-hemolysis,
CDC, Centers for Disease Control and Prevention.
The outbreak of food poisoning occurred in a canteen of an enterprise in Kunming, Province of Yunnan, Republic of China on March 2, 2006. At 12:15, 250 workers had lunch. From 1:20 to 8:30, 139 of these searched for medical care, 92 were heavily attacked, with nausea, vomiting with at least 2 bringups, and diarrhea with at least 3 evacuations. Most patients recovered approximately 24 h after presenting the acute symptoms. The dishes served existed of a soup, fried green peppers with douchi, stir-fried liver, steamed pumpkin, fried Chinese cabbage, and fried pork with green beans. It appeared that it was people who had consumed the fried green peppers with douchi who experienced the acute symptoms. For enumeration of bacteria in food residues and vomits, 20 samples were collected and analyzed by the Center for Disease Control and Prevention (CDC) of Kunming, China. In the fried green peppers with douchi and in uncooked douchi, B. cereus were identified in densities of 2.3×107 and 1.6×107 colony-forming units/g, respectively. No other food pathogens were identified in the douchi, in leftovers of the other served dishes, or in the rice (National Standard of the People's Republic of China GB/T 4789.2.3.4.5.10.11.15–2003). B. cereus was also identified from vomits. It was concluded that the douchi, produced by a local factory, was the source of the B. cereus causing the outbreak. Three strains were kept by the physicians and were available for further analysis.
The strains KM007-1, KM007-2, and KM008 were analyzed for salicin fermentation, starch hydrolysis, and hemolytic activity as described by Apetroaie et al. (2005). Growth at 6°C, 48°C, and 50°C on T3 agar plates (Travers et al., 1987) was recorded after 21 and 3–5 d, respectively. DNA was isolated according to the protocol of Hansen and Hendriksen (2001). The primers and polymerase chain reactions (PCR) for detection of hblA, hblC, hblD, nheA, nheB, and nheC were essentially performed as described by Hansen and Hendriksen (2001); ces and cesB by Ehling-Schulz et al. (2004, 2005); cytK1 and cytK2 by Guinebretiére et al. (2006); repX by Hu et al. (2009); and 16S/23S rDNA ITS by Willumsen et al. (2005). Amplification and sequencing of the panC, and the consequent determination of phylogenetic affiliation and food poisoning potential was performed according to the protocols and recommendations of Guinebretiere et al. (2008, 2010). Sequencing of the panC was performed by Macrogen, Netherlands. Multiple locus sequence typing (MLST) was performed as described by Priest et al. (2004). Sequencing, using both forward and reverse primers, was accomplished by Macrogen, Netherlands. The sequence between the primers EM1F1 and EM1R was determined using both forward and reverse primers by Macrogen, Netherlands. For high-performance liquid chromatography–mass spectrometry (HPLC-MS) analysis of cereulide, the bacteria were grown on tryptic soy or heart brain infusion agars (Oxoid) at 10°C for 10 d, 15°C and 20°C for 5 d, or at 30°C, 37°C, 40°C, 42°C, and 45°C for 3 d. After growth, the bacterial lawn from half a petri dish (9-cm diameter) was scraped off with the aid of an inoculation loop. The biomasses (0.127–0.552 mg fresh weight) were collected into 2.0-mL eppendorf tubes, and cereulide was extracted with 96% ethanol as described by Thorsen et al. (2009). The supernatant was collected and centrifuged at 15,000 ×g for 15 min.
HPLC-MS analysis was performed using an API 4000 (AB Sciex, Framingham, MA) after HPLC separation. The MS was used with electrospray ionization in positive mode. Full scans from 500 to 1300 atomic mass units (amu) were recorded.
The HPLC system consisted of an Ultimate 3000 dual-gradient low-pressure-mixing HPLC system (Dionex, Sunnyvale, CA). The separation was performed using a ternary gradient consisting of water, methanol, and acetonitrile on a Synergy Polar-RP column (L=150 mm, ID=2 mm, particles=4 μm; Phenomenex, Torrance, CA).
Cereulide was quantified on mass fragment 1170 (1169.5–1170.5) amu at retention time 8.50 min while valinomycin was quantified using mass fragment 1112 (1111.5–1112.5) amu at retention time 8.40 min. The cereulide was identified using the extract of F4810/72 as a standard. As no cereulide in pure form is available, the HPLC-MS was calibrated with valinomycin as an internal standard (Sigma-Aldrich, Brøndby, Denmark) (Häggblom et al., 2002). The calibration was then used to quantify cereulide leading toward data with some offset, which are, however, precise in respect to relative comparisons.
Results
Three B. cereus strains from the outbreak described above were analyzed. Two of them differing in colony morphology originated from the vomit of one person, while the third originated from unprepared douchi. The isolates were initially identified as being B. cereus by conventional phenotypic methods. This identification was confirmed by 16S rDNA sequencing and 16S-23S rDNA internal transcribed spacer profiling (Hansen and Hendriksen, 2001). Partial sequencing of panC revealed that the isolates KM007-2 and KM008 belong to B. cereus genetic group III and KM007-1 to genetic group IV. MLST after the scheme of Priest et al. (2004) assigned (
PCR analysis for genes encoding for the emetic toxin synthetase and three enterotoxins revealed that KM007-1 possess genes for the HBL, the NHE complexes, and cytotoxin K2, but not for the nonribosomal cereulide peptide synthetase, while KM007-2 and KM008 possess genes for the nonribosomal cereulide peptide synthetase and NHE. The sequences between the primers EM1F and EM1R of KM007-2 and KM008, which is a partial sequence of the nonribosomal cereulide peptide synthetase, were identical and they were also identical to the sequence of other ST26 members. Also, KM007-2 and KM008 possessed the gene for replicon X of the B. anthracis pXO1.
KM007-2 and KM008 were able to grow at 48°C, but not at 6°C and 50°C; KM007-1 was not able to grow at any of these three temperatures. KM007-1 was able to degrade salicin and starch. This was in contrast to KM007-2 and KM008. KM008 is hemolytic, KM007-2 weakly hemolytic, and KM007-1 nonhemolytic. HPLC-MS analysis for the emetic-toxin cereulide revealed that KM007-2 and KM008 also produced the toxin on tryptic soy as well as brain-heart-infusion agars. KM007-1 did not, as expected, produce this toxin. The concentrations produced varied for KM007-2 and KM008 between 87 and 177 μg/g fresh biomass at temperatures between 20°C and 37°C at these two agar media, but did not vary much at these three temperatures. This is in contrast to the strain F4810-72, which is considered as a type strain for cereulide production (Thorsen et al., 2009). This strain produced the most cereulide at 20°C in both media and limited amounts at 37°C. To analyze this discrepancy in more detail, a comparative study with triplicate incubations of KM007-2 and F4810-72 on BHI agar at 30°C–45°C was conducted (Fig. 1). It appears from Figure 1 that the two strains produced equal amounts of cereulide at 30°C, while KM007-2 produced statistically more cereulide than F4810/72 at 37°C, 40°C, and 42°C, while no cereulide was produced at 45°C.
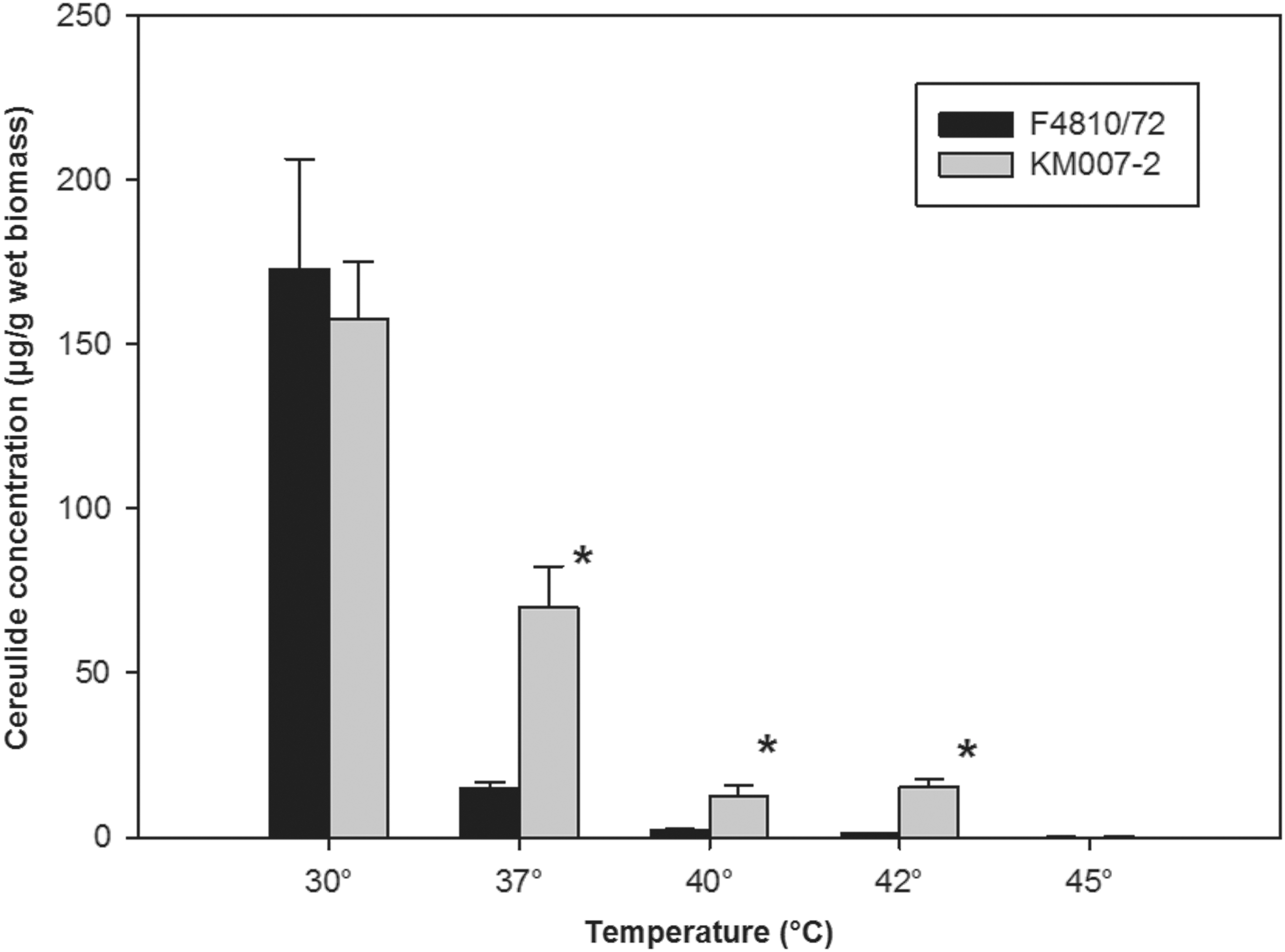
Cereulide production of Bacillus cereus F4810/72 and KM007-2 at 30°C, 37°C, 40°C, 42°C, and 45°C. Columns marked with an asterisk are the amount of cereulide produced by F4810/72 and KM007-2 that are statistically significantly different (p<0.05) (t-test).
Discussion
Here we report on a food-poisoning outbreak caused by B. cereus involving 139 people who had consumed infected fermented black beans (douchi). Two other cases of B. cereus food poisoning related to the consumption of douchi have previously been reported from China (Shen et al., 2005; Liu et al., 2006). B. cereus food-poisoning cases reported in China have also been affected by the consumption of other fermented or soybean products, such as bean curd (Zhang, 1997), soybean paste (Li, 2000), and home-made rice wine (Zhou et al., 2009). The consumption of homemade rice wine was even fatal for an adult male and a 7-year-old girl (Li and Huang, 1997). The occurrence of B. cereus, including cereulide-producing isolates, has also been reported from different African fermented seed–based food products (Thorsen et al., 2010, 2011) without any known involvement in food poisoning.
Three different B. cereus strains from the outbreak were characterized. Two of the strains possessed the synthetase gene cluster involved in the production of cereulide, and they were shown to be able to produce cereulide. Therefore, they are able to cause the emetic syndrome as experienced by most of the 139 workers. The third strain possesses the genes for the HBL, the NHE complexes, and cytotoxin K2; therefore, it might have been involved in the diarrheal syndrome experienced by some of the workers. The occurrence of different B. cereus strains showing diversity in biochemical and toxic properties associated with a food-poisoning case was also reported by Pirhonen et al. (2005).
We assigned by MLST the two cereulide-producing strains to the sequence type ST26 (Priest et al., 2004). This sequence type consists of strains unable to degrade starch and salicin, with varying hemolytic activity, which are unable to grow at 6°C and able to grow at 48°C and possess genes for the nonhemolytic enterotoxin NHE (Ceuppens et al., 2013). These characteristics correspond to the characteristics of KM007-2 and KM008. However, KM007-2 and KM008 are not identical because they differ in their hemolytic capabilities. KM007-2 and KM008 differ from most other strains of this type by producing the same level of cereulide at 20°C, 30°C, and 37°C; their production at 37°C approximates other strains' production at lower temperatures. Furthermore, it is shown that KM007-2 produces cereulide at 40°C as well as 42°C. Most strains, including F4810/72, have been shown to downregulate their cereulide production in response to an increasing temperature, with the highest concentration produced at ≈20°C, low production at 37°C, and no production at temperatures above 37°C (Häggblom et al., 2002; Apetroaie-Constantin et al., 2008). However, the cereulide production of the strains RIVMBC00067 and RIVMBC00075 remained at temperatures between 20°C and 37°C (Apetroaie-Constantin et al., 2008). The cereulide content at 37°C has been shown to vary between biomass grown on different agar media. These included tryptic soy agar, where relatively high amounts were produced (Apetroaie-Constantin et al., 2008). The differences in cereulide production between KM007-2 and KM008 in comparison to other strains is most likely caused by variations in the regulation of the synthesis of cereulide, as the sequence of the partial ces is identical to other strains of ST26. Furthermore, it is most likely that ces of these two strains are placed on a plasmid, as in other cereulide producers (Hoton et al., 2005; Ehling-Schulz et al., 2006b), as they possess the replicon X of B. anthracis pXO1. The ability to produce cereulide at 37°C makes it likely that it can be produced in the human gut in addition to production occurring in food.
The third strain KM007-1 belongs to genetic group IV of Guinebretière et al. (2008) and is assigned to a new sequence type (ST 752) by MLST. Genetic group IV strains grow at temperatures between 10°C and 45°C; they are generally cytotoxic and some can be highly cytotoxic (Guinebretière et al., 2008). The new sequence type is phylogenetically closely related to strains that are cytotoxic, and have been isolated from blood or from food-poisoning cases. As with other strains in this group, it possesses the genes for HBL, NHE, and cytK2, and it affects salicin and degrades starch. However, it is very unusual that a strain possessing genes for HBL is nonhemolytic (Beecher et al., 1995). It is likely that KM007-1, based on its close phylogenetic relationships to cytotoxic strains and that it possesses genes for three enterotoxins, have been involved in the diarrheal syndrome experienced by some of the workers.
Conclusions
In conclusion, a case of food poisoning influencing 139 workers consuming fermented black beans (douchi) is described. The case included emesis as well as diarrhea and was caused by B. cereus. Three B. cereus strains from the case are characterized; two produced cereulide, the emetic toxin; one of these strains was unusual by being able to produce the toxin at temperatures above 37°C. Therefore, a potential for cereulide production in the gut exists. If this potential is realized, it calls for new precautions. The third strain, a new sequence type possessing genes for enterotoxins, might have been involved in the diarrheal syndrome affecting some workers.
Footnotes
Acknowledgments
This study was partly funded by the Sino-Danish bilateral cooperation and Hubei Provincial Department of Education (project No. Q20111703). The help from the Danida Fellowship Centre is greatly appreciated.
Disclosure Statement
No competing financial interests exist.