
Abstract
This study evaluated the influence of starvation on the radiation resistance and injury in Escherichia coli O157:H7, following electron beam irradiation in 0.85% (wt/vol) saline and in apple juice. Washed exponential-phase cells of E. coli O157:H7 that were grown in tryptic soy broth (TSB) at 35°C were starved in 0.85% saline (25°C) for 10 days. Exponential- or stationary-phase cells grown in TSB (35°C) served as controls. Samples of 0.85% saline or pasteurized apple juice, inoculated with control cells or cells starved for 8 days, were exposed to electron beam irradiation at doses ranging from 0.0 (control) to 0.70 kGy. The E. coli survivors were enumerated by plating diluted samples on tryptic soy agar or on Sorbitol McConkey agar and counting bacterial colonies after incubation (35°C) for 24 h. Starved cells consistently exhibited higher irradiation D-values than controls (p<0.05). The D-values for control and starved E. coli O157:H7 in 0.85% saline were 0.11 and 0.26 kGy, respectively; D-values in apple juice were 0.16, 0.19, and 0.33 kGy for exponential, stationary, and starved cells, respectively. Irradiation (0.70 kGy) of E. coli O157:H7 in apple juice reduced numbers of exponential- and stationary-phase cells by ∼4.32 and 3.74 logs, respectively, whereas starved cells were reduced by only 2.20 logs. Exponential-phase cells exhibited the lowest resistance to irradiation, and sublethal injury in survivors of this group was higher than that of stationary-phase or starved cells irradiated at 0.50 or 0.70 kGy (p<0.05). The results of this study indicate that starvation-induced stress cross-protects E. coli O157:H7 from ionizing radiation and should be considered an important factor when determining irradiation D-values for this pathogen.
Introduction
I
As demonstrated with other bacteria, starvation in E. coli O157:H7 can result in the organism synthesizing protective proteins coded for by the rpoS gene (Cheville et al., 1996). Protective proteins are believed to be associated with cellular resistance to a range of chemical and physical challenges (Rowe et al., 1999). Such increased microbial resistance to chemical and physical challenges can decrease the lethal effect of food preservation processes including acidification, heating, and irradiation.
Food irradiation is an effective technology for killing E. coli O157:H7 and other vegetative pathogens. Its antimicrobial mode of action involves disruption of DNA by causing single- and/or double-strand breaks. The inability of microorganisms to repair the damaged DNA results in death (Olson, 1998). The antimicrobial effectiveness of irradiation is altered by factors such as temperature, radiation dose, food composition, and gaseous atmosphere (Mendonca, 2002). Also, changes in the physiological state of microorganisms due to prior stress may increase microbial resistance to irradiation. For example, Buchanan et al. (1998) demonstrated that induced acid resistance in E. coli O157:H7 cross-protected this pathogen against gamma radiation in clarified apple juice. Also, starved Listeria monocytogenes exhibited substantially increased tolerance to electron beam radiation in ground pork compared to exponential- and stationary-phase cells (Mendonca et al., 2004).
Starvation-induced stress in foodborne pathogens is relevant to the food industry because water, used for cleaning or rinsing food contact surfaces or even added directly to food, is generally of a low nutrient status (Rowe et al., 2000). Also, pathogens may survive in water (without nutrients) on food contact surfaces long enough to induce the stress response, which may cross-protect them from subsequent food preservation methods (Rowe et al., 2000).
To date, published research on the extent to which starvation can cross-protect E. coli O157:H7 against ionizing radiation is lacking. Accordingly, one objective of this study was to determine the effect of starvation on radiation resistance of E. coli O157:H7 that are exposed to electron-beam irradiation in 0.85% saline and in apple juice. A second objective was to determine the extent of sublethal injury in exponential phase, stationary phase and starved E. coli O157:H7 following electron-beam irradiation in apple juice.
Materials and Methods
Microorganisms and culture conditions
Four strains of E. coli O157:H7 (ATCC 43895, ATCC 43894, FRIK 125, and C 467) were used in the study. Strains ATCC 43895, ATCC 43894, and C 467 were obtained from the culture collection of the Microbial Food Safety Laboratory of Iowa State University. Strain FRIK 125, an isolate from an apple cider outbreak, was provided by Dr. Charles Kaspar, University of Wisconsin. Stock cultures were stored at −70°C in brain heart infusion broth (Difco Laboratories, Detroit, MI) supplemented with 10% glycerol until used. Each stock culture was transferred at least twice in 10 mL tryptic soy broth (TSB) (Difco) and incubated overnight at 35°C prior to each experiment.
Starvation of E. coli O157:H7
A portion (1.0 mL) of E. coli O157:H7 culture was transferred to TSB (100 mL) in a screw-capped 250-mL Erlenmeyer flask. The inoculated medium was incubated at 35°C with shaking at 150 rpm in a gyrorotary shaker incubator (New Brunswick Scientific Co. Inc., Edison, NJ). Growth was monitored with a spectrophotometer (Spectronic 1201; Milton Roy Co., Rochester, NY) via optical density measurements at 600 nm. When E. coli O157:H7 grew to an optical density of 0.10–0.15, exponential-phase cells were harvested by centrifugation (10,000×g, 10 min, 4°C) and washed once in 0.85% (wt/vol) NaCl. The cell pellets were suspended in fresh saline to give a final cell concentration of approximately 2.0×107 colony-forming units (CFU)/mL as determined by plate counts. The cells were starved by holding the cell suspension statically in a screw-capped 250-mL flask at 25°C for 10 days.
Determination of cell viability and radiation resistance
During starvation, samples of E. coli O157:H7 cell suspension were removed for determining cell viability and resistance to electron-beam irradiation. Serial dilutions of the cell suspension were prepared in 0.1% peptone water, and 0.1-mL aliquots of appropriate dilutions were surface plated on tryptic soy agar (TSA) and Sorbitol MacConkey agar (SMA) (Difco). All inoculated agar plates were incubated aerobically at 35°C and bacterial colonies were counted at 24 h. Samples (10-mL) of cell suspension were placed into sterile screw-capped test tubes and held in crushed ice before treatment with electron-beam irradiation as described in the section on irradiation and dosimetry. Numbers of survivors in irradiated samples were determined as previously described for determination of cell viability.
Preparation and inoculation of test samples
Pasteurized, clarified apple juice (pH 3.97) was purchased from a local supermarket in Ames, IA and stored in a laboratory refrigerator (4°C) before inoculation. The level of background microflora in the apple juice was determined by surface-plating serial dilutions (0.1% peptone) on TSA. Washed exponential-phase cells of E. coli O157:H7 in saline were prepared from a TSB culture as previously described. Stationary-phase cells from an overnight (18 h) culture in TSB (35°C) were harvested by centrifugation (Sorvall® Super T21; Sorvall Product, L.P., Newtown, CT) at 10,000×g for 10 min at 4°C, washed once in saline to remove residual growth medium, then suspended in fresh saline (4°C). Cells starved for 8 days in saline (25°C) were harvested by centrifugation, washed, and suspended in fresh saline (4°C). Suspensions of starved or nonstarved cells were adjusted to give approximately 108 CFU/mL based on plate counts. Tubes of sterile saline or apple juice (10 mL per tube) were each inoculated with 0.1 mL of E. coli O157:H7 cells (exponential, stationary, or starved) to give approximately 106 CFU/mL. Tubes of saline were inoculated with only exponential and starved cells.
For the tests performed in apple juice, a cocktail made of equal volumes of all four pathogens was used. Screw-capped tubes of inoculated samples were held in a 50:50 ice/water mixture prior to irradiation.
Irradiation treatment and dosimetry
Inoculated tubes of saline or apple juice were irradiated at the Iowa State University Linear Accelerator Facility, which has a MeV CIRCE III Linear Electron Accelerator (MeV Industrie S.A., Jouy-en-Josas, France). All samples were irradiated in duplicate at five target doses (0, 0.15, 0.30, 0.50, and 0.70 kGy) in the electron beam mode at an energy level of 10 MeV. Dose rates were 12, 20, and 45 kGy/min, for 0.15 and 0.30, 0.50, and 0.70 kGy, respectively. Dose rates differed because the linear accelerator had to be operated at a low power to apply the lower doses (0.15 and 0.30 kGy). Each target average dose represents an arithmetic average of doses measured at the top and bottom surfaces of the test samples.
Dosimeter alanine pellets, 5 mm (length) by 5 mm (diameter) (Bruder Analytische Messtechnik, Rheinstetten, Germany), were used to determine the maximum and minimum absorbed radiation doses. Alanine pellets were placed at the top and bottom surfaces of one of the duplicate tubes of saline or apple juice. Immediately after irradiation, the pellets were placed in a Bruker EMS 104 EPR Analyzer to measure absorbed doses by electron paramagnetic resonance. The dose maximum/minimum ratios for 0.15, 0.30, 0.50, and 0.70 kGy were 1.13, 1.20, 1.16, and 1.07, respectively. The average absorbed dose was derived from the arithmetic average of the top and bottom surface readings.
Microbiological analysis
All samples were held at 4°C and analyzed within 2 h after irradiation. Serial dilutions of saline or apple juice in 0.1% peptone were prepared and 0.1-mL aliquots of appropriate dilutions were surface-plated, in duplicate, onto plates of TSA (for saline samples) and on both TSA and SMA (for apple juice samples). In instances when numbers of E. coli O157:H7 survivors decreased to very low levels (for example, <10 CFU/mL), 1.0-mL samples of undiluted saline or apple juice were plated directly onto appropriate agar plates. All inoculated plates were incubated aerobically at 35°C and bacterial colonies were counted at 24 h.
Calculation of D-values
The D-values (radiation dose that produces 90% reduction in viable cells) expressed in kGy were determined by plotting the log10 number of survivors per mL versus radiation dose (kGy) using Microsoft Excel 2010 Software (Microsoft Inc., Redmond, WA). Linear regression analysis (Ostle et al., 1975) was used to determine the line of best fit for the data. The D-value was determined by calculating the negative reciprocal of the slope of the regression curve.
Determination of sublethal injury
For each replicate experiment, survivor curves based on recovery of bacterial colonies on TSA and SMA were prepared. The percent sublethal injury for each strain at each radiation dose used (0.0 to 0.70 kGy) was determined by dividing the difference in numbers of E. coli O157:H7 survivors on nonselective (TSA) and selective (SMA) agar by the numbers of survivors on TSA followed by multiplication by 100.
Statistical analysis
All experiments were replicated three times and results are reported as averages. The D-value data were compared using the Tukey method (SAS Institute, Cary, NC).
Results and Discussion
Cell viability during starvation
Figure 1 shows the viability of E. coli O157:H7 (ATCC 43895, ATCC 43894, FRIK 125, and C 467) during starvation in 0.85% saline (25°C) based on enumeration of bacterial colonies on TSA. Initial populations of E. coli ATCC 43895, ATCC 43894, and FRIK 125 decreased by about 0.5 logs during the first 2 days of starvation. Initial populations of strain C 467 decreased by about 0.3 logs by day 4 of starvation. Numbers of culturable cells from all four strains remained relatively constant during days 6–10. These observations are consistent with those of Jouper-Jaan et al. (Jouper-Jaan et al., 1992), who reported that after 9 days of multiple (energy and nutrient) starvation, only about 77% of the initial populations of E. coli and Vibrio spp. were culturable.
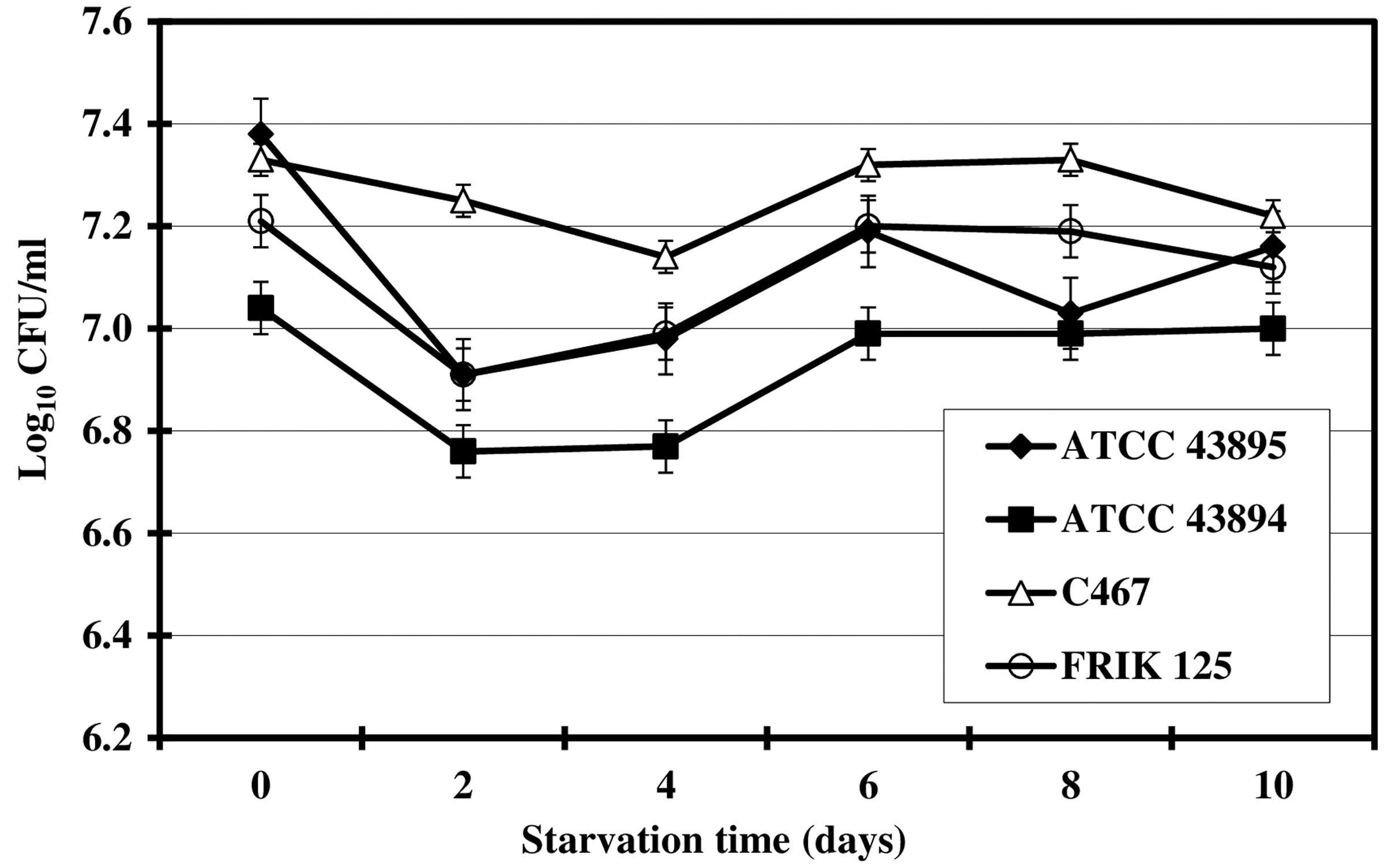
Viability of four strains of Escherichia coli O157:H7 (ATCC 43895, ATCC 43894, FRIK 125, and C467) during multiple nutrient starvation for 10 days in 0.85% saline at 25°C. Populations are the means of three values±SDs. Error bars represent SDs (n=3, i.e., three independent experiments). CFU, colony-forming units.
Not all cells in a starving bacterial population remain viable based on plate counts. Starved bacteria have been reported to actively transport and metabolize nutrients but fail to multiply and form colonies under standard plating conditions (Roszak et al., 1987). Kurath et al. (1983) studied the physiology of a marine Pseudomonas sp. during starvation-survival of this organism. They demonstrated that within the population of starved cells, numbers of respiring cells (based on active reduction of 2-[p-indophenyl]-3-[p-nitrophenyl]-5-phenyl tetrazolium chloride [INT]) were 10-fold higher than the viable counts (based on plate counts). This difference between plate counts and INT-reducing counts indicates the presence of a subpopulation of nonculturable but respiring cells that is about 1.0 log higher than culturable cells. Therefore, decreases in viability of E. coli O157:H7 reported in the present study and that of Vibrio spp. reported by Jouper-Jaan et al. (1992) may not reflect actual death of a part of the initial population. Further research is needed to determine whether starvation induces a viable but nonculturable state in E. coli O157:H7.
Radiation resistance in saline
The radiation resistance (D-value) of E. coli O157:H7 during starvation in 0.85% saline at 25°C is shown in Figure 2. The D-value for each of the four strains at day 0 of starvation was approximately 0.11 kGy. All D-values increased as starvation time increased and reached a maximum at about day 8 of starvation. Maximum D-values for starved strains ATCC 43895, ATCC 43894, C 467, and FRIK 125 were 0.35, 0.23, 0.19, and 0.26 kGy, respectively. These results indicate that starvation increases the resistance of E. coli O157:H7 to electron-beam irradiation.
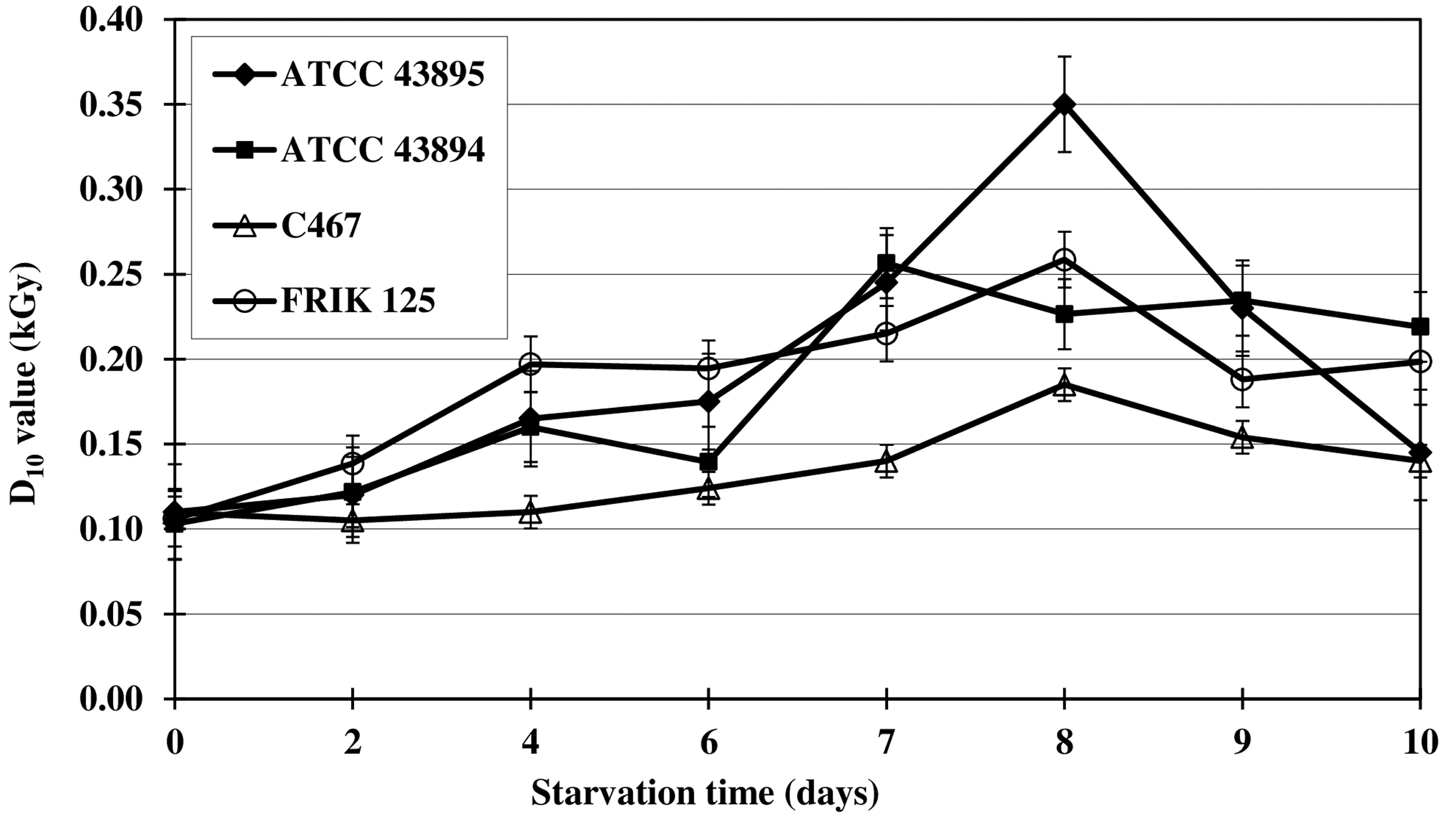
Radiation resistance (D-values) of four strains of Escherichia coli O157:H7 (ATCC 43895, ATCC 43894, FRIK 125, and C467) during multiple nutrient starvation for 10 days in 0.85% saline at 25°C. Populations are the means of three values±SDs. Error bars represent SDs (n=3, i.e., three independent experiments).
Radiation resistance in apple juice
Survivors of a four-strain mixture of E. coli O157:H7 (starved and non-starved exponential- and stationary-phase cells) following irradiation in pasteurized apple juice (pH 3.97) are shown in Figure 3A and B. Populations of survivors following irradiation in apple juice are based on bacterial colony counts on TSA (Fig. 3A) and SMA (Fig. 3B). Populations of E. coli O157:H7 survivors on TSA were generally higher than those on SMA. Populations of the organism in apple juice decreased with increasing radiation dose irrespective of the plating medium used. The higher numbers of survivors on TSA compared to SMA suggest that a portion of the surviving population may have been sublethally injured by irradiation. Depending on the extent of metabolic injury, injured cells can become sensitive to chemical agents in selective media (Ray et al., 1992). The colonies that developed on TSA represented both injured and noninjured cells, whereas only noninjured cells formed colonies on SMA. The ability to recover injured pathogens in foods during culturing procedures is important to the food industry because injured pathogens may recover in foods and regain their pathogenicity (Ray et al., 1992). Therefore, in the present study, the use of TSA counts for estimating E. coli O157:H7 survivors is justified considering the fact that injured pathogens could potentially have the same health impact as noninjured ones. Based on TSA counts, 0.7 kGy reduced initial populations of exponential- and stationary-phase (control) cells by 4.32 and 3.74 log cycles, respectively, whereas starved cells were reduced by only 2.2 log cycles (Fig. 3A). These results indicate that the starved cells have a greater ability to survive the lethal effects of ionizing radiation compared to exponential- or stationary-phase cells.
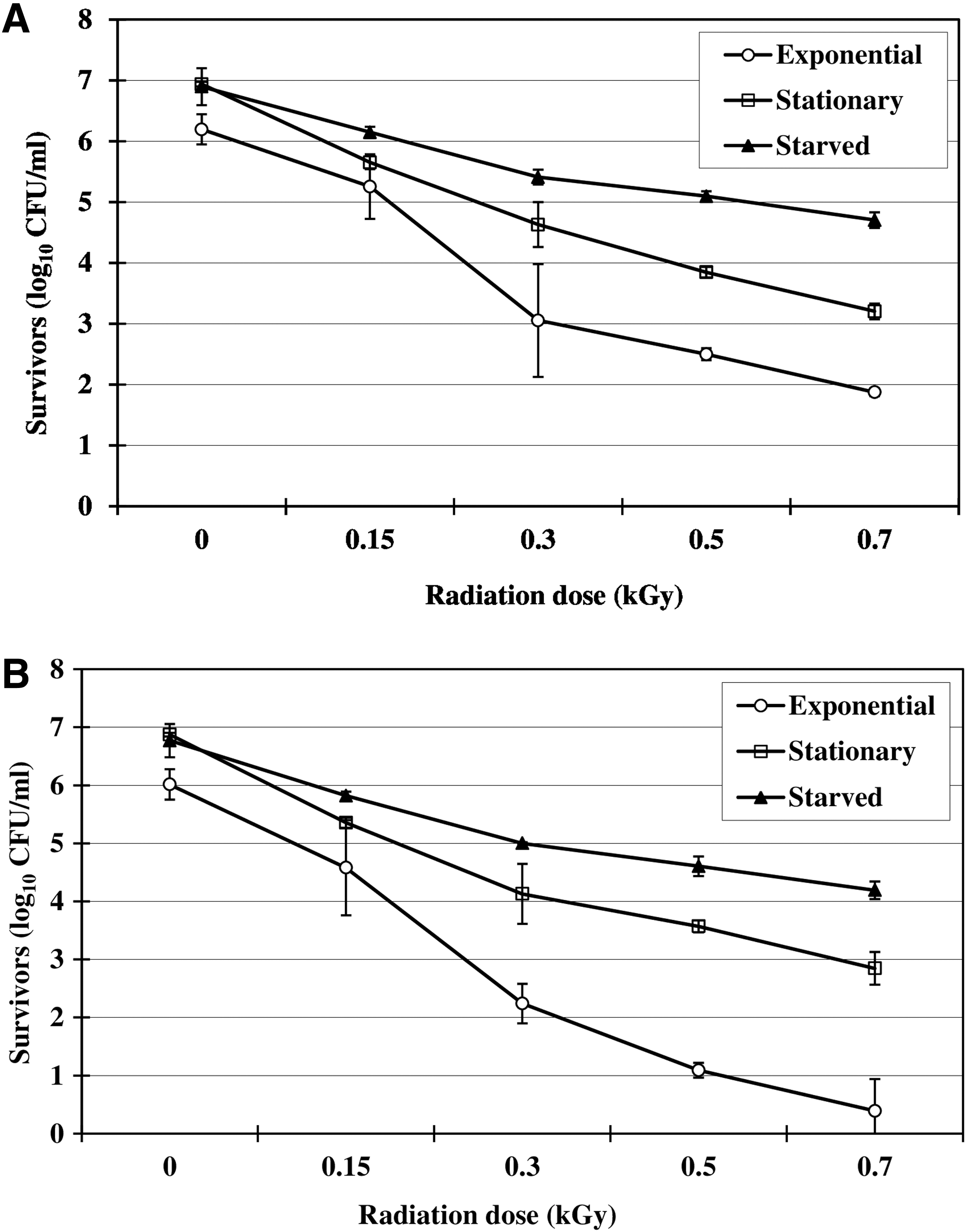
Survival of starved and nonstarved (exponential- and stationary-phase) Escherichia coli O157:H7 following irradiation in apple juice at 4°C irradiation in apple juice at 4°C tryptic soy agar. Survivors were recovered on tryptic soy agar
The radiation resistance (D-value) for each of the four E. coli O157:H7 strains (exponential-phase or starved) in 0.85% saline and for the four-strain mixture (exponential-phase, stationary-phase, or starved) in apple juice is shown in Table 1. Escherichia coli O157:H7 in three physiological states (exponential, stationary, and starved) were used in the present study to determine whether there were marked differences in radiation resistance among these states. Stationary-phase cells from overnight (18–24 h) laboratory cultures are typically used in experiments on pathogen inactivation by food processing methods; however, bacteria that contaminate foods may come from natural environments where nutrient limitation or starvation is the norm rather than the exception (Koch, 1971; Roszak and Colwell, 1987). From a practical perspective, it is therefore important to determine the radiation resistance of starved bacteria that may contaminate foods from nutrient-depleted environmental sources. To determine whether starvation altered the irradiation resistance of E. coli O157:H7 strains, exponential cells were used as a control. D-values for each of the four strains and the four-strain mixture were significantly higher in starved cells than in control (exponential- or stationary-phase) cells (p<0.05). It has long been recognized that stationary-phase cells are generally more resistant to ionizing radiation than exponential-phase cells. Previous studies have demonstrated that E. coli O157:H7 cells in the stationary growth phase, and also those that endure starvation, synthesize specific sets of stress proteins. These stress proteins increase the cells' resistance to a range of chemical and physical challenges (Arnold et al., 1995).
Survivors were recovered on tryptic soy agar.
Means (±SDs) of three replicate experiments.
Means with different superscript letters in the same column are significantly different (p<0.05).
N/A, not applicable; stationary phase cells were not used for tests with saline.
A four-strain mixture of the pathogen.
The synthesis of a substantial amount of the stress proteins in starved or stationary-phase cells is controlled by the rpoS gene (Cheville et al., 1996). By using an rpoS mutant, Rowe and Kirk (Rowe and Kirk, 2000) were able to demonstrate a central role for the rpoS gene in the induction of heat resistance following starvation stress in E. coli O157:H7. Even though the rpoS gene mediates the synthesis of protective proteins in stationary-phase and starved E. coli O157:H7, it is surprising that the starved cultures in the present study were much more resistant to radiation in apple juice than the stationary-phase cultures (Table 1). Further research is needed to fully explain the relatively high radiation resistance of starved E. coli O157:H7.
Radiation-induced sublethal injury
Figure 4 shows the levels of sublethal radiation injury in surviving populations of E. coli O157:H7 at each absorbed dose. Injury of exponential-phase cells was consistently higher than that of stationary-phase or starved cells at all doses. However, only at the higher doses (0.5 and 0.7 kGy) was the extent of this injury significantly greater than that of starved or stationary-phase cells (p<0.05). In this study, sublethally injured cells represent cells that, at the time of analysis, were unable to form colonies on SMA. However, they were able to form colonies on TSA. These cells evidently endured severe damage. Severely injured cells require more intricate repair mechanisms that may need special environmental conditions to become fully functional. Stationary-phase cells or starved cells might have endured less sublethal injury or developed very efficient mechanism for repair of their lesions.
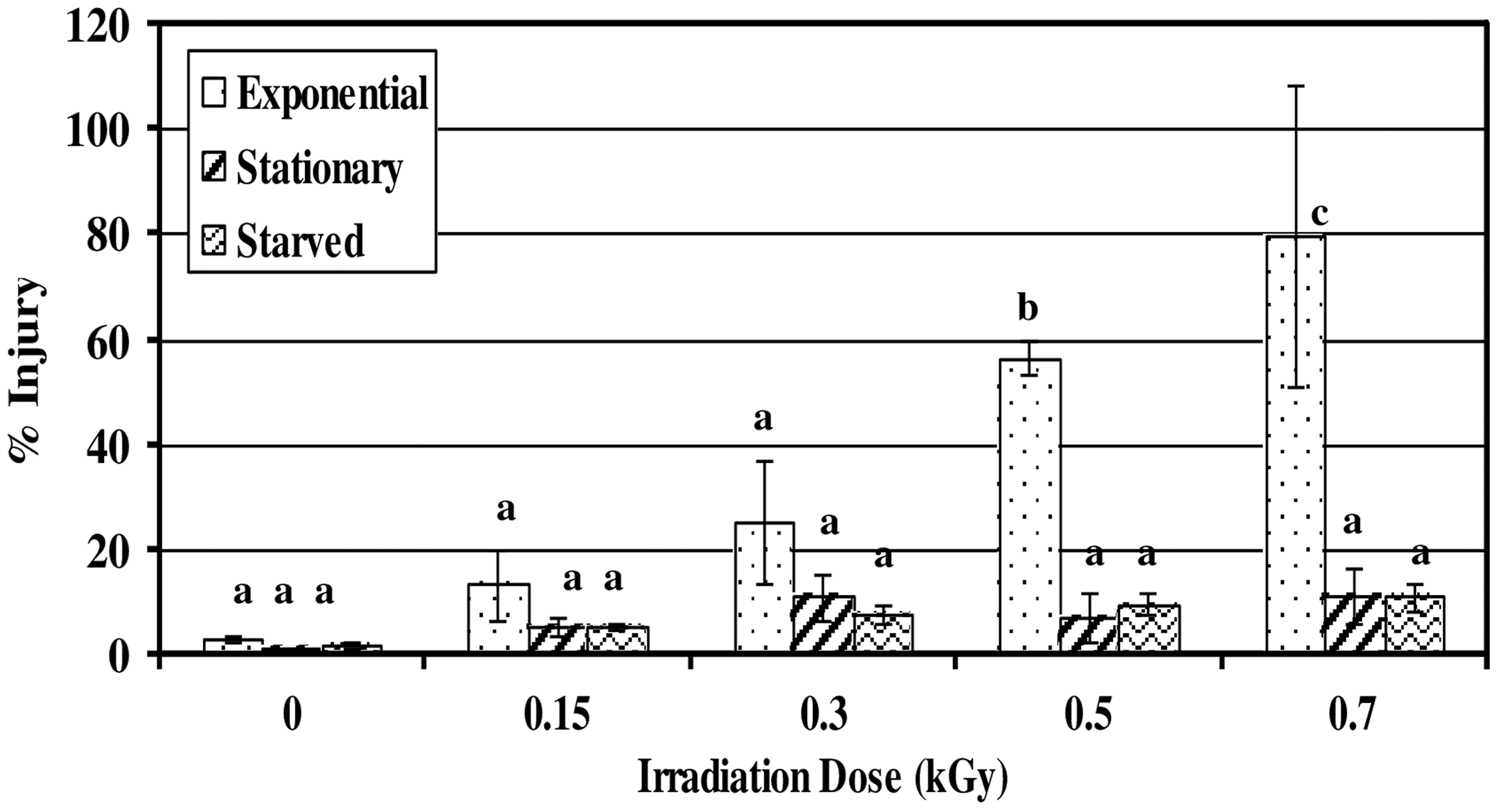
Sublethal injury of a four-strain mixture (ATCC 43895, ATCC 43894, FRIK 125, and C467) of Escherichia coli O157:H7 following electron-beam irradiation in apple juice. Error bars represent SDs (n=3, i.e., three independent experiments). Bars that do not share a common letter are significantly different.
The starvation of E. coli O157:H7 in 0.85% saline was an attempt to simulate the survival of this pathogen in water (devoid of nutrients) on food contact surfaces for an extended period of time, sufficient to stimulate the stress response. The use of non-stressed pathogens in food safety experiments is likely to underestimate the full ability of these organisms to survive food preservation methods. Therefore, it would be practical to use D-values of pathogens in their most radiation-resistant state as a basis for establishing adequate food irradiation processes.
The present study provides evidence that starvation in E. coli O157:H7 dramatically increases the resistance of this pathogen to electron-beam irradiation. The development of irradiation processes for inactivating E. coli O157:H7 without altering the desirable sensory attributes of food products depends on accurate assessment of the organism's radiation resistance (D-value). Even small differences in D-values can have a significant effect on the antimicrobial efficacy of radiation processes because of the exponential nature of microbial destruction by irradiation. Further research on the radiation resistance of other stressed pathogens is needed to provide food processors with more realistic microbial resistance data. The acquisition of such data would permit the design of adequate processes that ensure the safety of irradiated foods.
Conclusions
Based on the results of the present study, starvation in E. coli O157:H7 increases this pathogen's resistance to electron-beam irradiation and reduces the extent of sublethal injury in survivors of starved cells. Starvation-induced cross-protection of pathogens against food irradiation treatment should be taken into account when determining irradiation D-values.
Footnotes
Acknowledgment
The authors thank Dr. Charles Kaspar, University of Wisconsin, Madison, for supplying one of the bacterial strains (FRIK 125) used in this experiment. The authors also thank Meijun Zhu for statistical assistance and Michael Holtzbauer, irradiator operator, for technical assistance. This project was supported by the USDA/CSREES National Research Initiative Competitive Grants Programs, Project No. 98-35502-6604.
Disclosure Statement
No competing financial interests exist.