
Abstract
The oxidation of formate constitutes a main energy source for Campylobacter jejuni, an important foodborne pathogen. Formate occurs in the hosts' gut as a byproduct of fermentation, which may contribute to C. jejuni's adaptation to that niche. C. jejuni possesses a periplasmic formate dehydrogenase (Fdh) that facilitates the metabolism of formate. In C. jejuni NCTC-11168, a poorly invasive strain, Fdh localizes to the periplasm via the twin arginine translocation (Tat) system, and the fdhA encodes the signature Tat signal motif. However, screening the genome of C. jejuni 81-176, a highly invasive strain, showed that the Tat signal motif was encoded on a separate gene (CJJ81176_1504) directly upstream of the fdhA (CJJ81176_1503). This possibly suggested a difference in formate metabolism between these two strains. Therefore, we investigated whether the Fdh was Tat dependent in C. jejuni 81-176. For this purpose, we measured formate respiration in C. jejuni 81-176 (wild-type), a Tat mutant (ΔtatC-81-176), and a complementation strain (C-ΔtatC-81-176) using BIOLOG-AN plates. We also used a viologen-based assay to specifically measure the Fdh enzymatic activity in whole cells and periplasmic preparations of the wild-type and mutant strains. Collectively, our data indicated that the Fdh activity and localization in C. jejuni 81-176 were dependent on a functional Tat system. The proper maturation and maintenance of Fdh are essential for cognate energy production, which might explain the association of the Fdh with the Tat system across different strains. The latter highlights the potential of the Tat system and Fdh as appealing targets for novel anti-C. jejuni therapeutics.
Introduction
C
In wild-type C. jejuni NCTC-11168, Fdh is encoded by fdhABCD (Cj1511c–Cj1508c) and the maturation, periplasmic localization, and subsequent activity of this RP are dependent on a functional twin arginine translocation (Tat) system (Hitchcock et al., 2010). In this strain, a typical Tat signal motif (TRRSFLK) is located in the N-terminus of the FdhA subunit. In comparison, the genome of C. jejuni 81-176 shows that the FdhA (CJJ81176_1503) does not possess a Tat signal motif (Fig. 1A), while cognate Fdh activity and localization were not previously experimentally shown to be Tat dependent. This is important, because it might mark a principal divergence in energy metabolism and host adaptation potential of this highly invasive strain (Oelschlaeger et al., 1993). Here, we investigated the activity and localization of the Fdh and its relationship to the Tat system in C. jejuni 81-176.
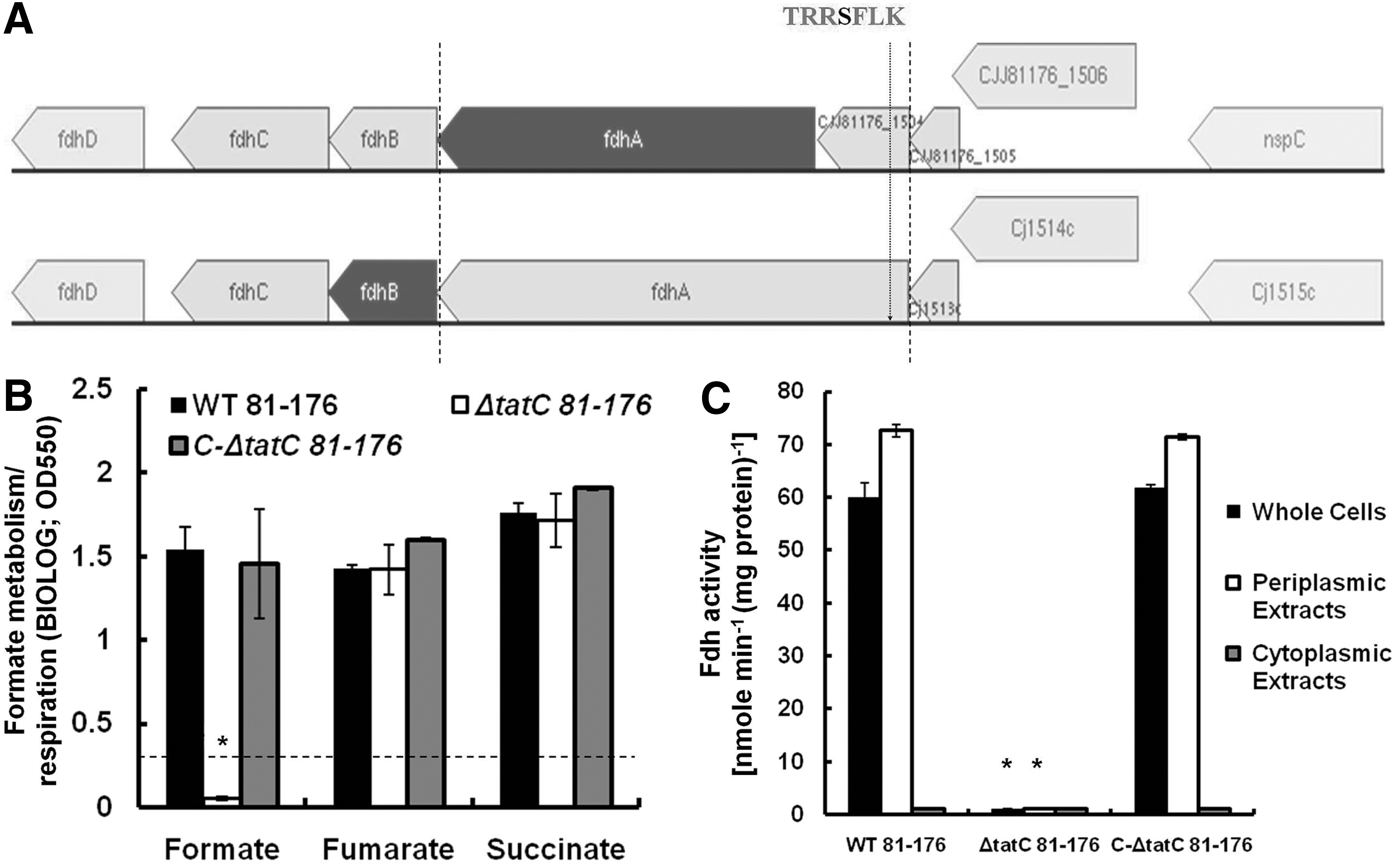
The arrangement of the formate dehydrogenase (Fdh) operon, formate respiration, and Fdh activity in Campylobacter jejuni 81-176.
Materials and Methods
Bacterial strains and growth conditions
The wild-type C. jejuni 81-176, a Tat (ΔtatC-81-176) mutant, and a complementation (C-ΔtatC-81-176) strain (Rajasehkara et al., 2009) were used in this study. The strains were cultured on Mueller-Hinton (MH) agar under microaerobic conditions (85% N2, 10% CO2, 5% O2) at 42°C. Laked horse blood (5%, Oxoid), antibiotics (chloramphenicol: 20 μg mL−1, kanamycin: 50 μg mL−1), and the Campylobacter selective supplement (SR155E, Oxoid) were added to the MH medium when necessary.
Assessment of formate respiration
Formate metabolism/ respiration was assessed using BIOLOG-AN plates (BIOLOG Inc., California) (Brandl et al., 2004), which contain substrates that can be metabolized by bacteria, resulting in the production of a purple formazan. The latter can be quantified spectrophotometrically at 550 nm. The C. jejuni strains were grown on blood agar for 24 h. The cultures were then suspended in 10 mM phosphate buffer (pH 7.0), centrifuged, washed twice, and resuspended to achieve an optical density (OD)600 of 0.7. The suspensions were dispensed into BIOLOG-AN plates (100 μL/well) that were then incubated microaerobically at 42°C for 48 h before measuring the OD550.
Quantification of Fdh activity
The Fdh activity was measured as described in Hitchcock et al. (2010). C. jejuni cells (OD600 0.2) were suspended in an oxygen-restricted solution containing 25 mM sodium phosphate buffer (pH 7) with 1 mM benzyl-viologen and 10 mM sodium formate. The mixture was placed in sealed cuvettes and the absorbance was measured spectrophotometrically; an absorption coefficient of 8.6 mM−1/cm−1 at 578 nm was used for benzyl-viologen. Additionally, periplasmic (100 ng/μL) and cytoplasmic extracts from each strain were isolated using the Peripreps™ Periplasting kit (Epicentre). The extracts were assayed for Fdh activity as described earlier but the buffer was replaced with 10 mM Tris/HCl (pH 7.5).
Results and Discussion
Analysis using BIOLOG-AN plates showed that the C. jejuni 81-176 and C-ΔtatC-81-176 strains were capable of metabolizing formate (Fig. 1B). However, ΔtatC-81-176 was unable to use formate (Fig. 1B). To ensure that the latter was due to the impairment of the Tat system, we investigated whether ΔtatC-81-176 was able to metabolize fumarate and succinate. The metabolism of these two substrates is mainly facilitated by the Tat-independent fumarate reductase (Cj0408-Cj0410) (Weingarten et al., 2009). Our results showed that the metabolism of fumarate and succinate by ΔtatC-81-176 and the wild-type was statistically similar (p>0.05) (Fig. 1B). Using a viologen-based assay, we showed that whole cells and the periplasmic extract of ΔtatC-81-176 lacked Fdh activity (Fig. 1C). Both the wild-type and C-ΔtatC-81-176 displayed statistically similar Fdh activity (P>0.05) (Fig. 1C). When the cells of the latter strains were fractionated, Fdh activity was only noted in the periplasmic extracts but not in the cytoplasmic fractions (Fig. 1C). Taken together, our data show that the Fdh activity and periplasmic localization in C. jejuni 81-176 are dependent on a functional Tat system. This was also confirmed using sodium dodecyl sulfate gel analysis of the periplasmic protein fractions followed by protease digestion and Nano/liquid chromatography coupled to tandem mass spectrometry analysis of the peptides. The analysis led to the identification of an Fdh peptide that belonged to the periplasm of the wild-type but was absent in the ΔtatC-81-176 profiles (data not shown). These findings corroborate those reported for C. jejuni NCTC-11168 (Hitchcock et al., 2010) and establish the association of the Tat system with Fdh in these different strains. After comparing the fdh operons and sequences from C. jejuni 81-176 and NCTC-11168 (Supplementary Table S1 and Supplementary Fig. S1; Supplementary Data are available online at
Footnotes
Acknowledgments
Research in the Rajashekara laboratory is supported by funds from the Agriculture and Food Research Initiative (AFRI, US Department of Agriculture), the Ohio Agricultural Research and Development Center, and the Ohio State University. We thank Dr. Malak Esseili for her help in the sodium dodecyl sulfate gel analysis and protein purification.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.