
Abstract
Escherichia coli O157:H7 has been the causative agent of many outbreaks associated with leafy green produce consumption. Elucidating the mechanism by which contamination occurs requires monitoring interactions between the pathogen and the plant under typical production conditions. Intentional introduction of virulent strains into fields is not an acceptable practice. As an alternative, attenuated strains of natural isolates have been used as surrogates of the virulent strains; however, the attachment properties and environmental stabilities of these attenuated isolates may differ from the unattenuated outbreak strains. In this study, the Shiga toxin (stx 1, stx 2, and/or stx 2c) genes as well as the eae gene encoding intimin of two E. coli O157:H7 outbreak isolates, F4546 (1997 alfalfa sprout) and K4492 (2006 lettuce), were deleted. Individual gene deletions were confirmed by polymerase chain reaction (PCR) and DNA sequencing. The mutant strains did not produce Shiga toxin. The growth kinetics of these mutant strains under nutrient-rich and minimal conditions were identical to those of their wild-type strains. Attachment to the surface of lettuce leaves was comparable between wild-type/mutant pairs F4546/MD46 and K4492/MD47. Adherence to soil particles was also comparable between the virulent and surrogate pairs, although the F4546/MD46 pair exhibited statistically greater attachment than the K4492/MD47 pair (p≤0.05). Wild-type and mutant pairs F4546/MD46 and K4492/MD47 inoculated into wet or dry soils had statistically similar survival rates over the 7-day storage period at 20°C. A plasmid, pGFPuv, containing green fluorescent protein was transformed into each of the mutant strains, allowing for ease of identification and detection of surrogate strains on plant material or soil. These pGFPuv-containing surrogate strains will enable the investigation of pathogen interaction with plants and soil in the farm production environment where the virulent pathogen cannot be used.
Introduction
S
O157 strains contain a 4.1-Mb gene backbone that is conserved in all E. coli stains, with an additional 1.4 Mb obtained from horizontal transfer of foreign DNA from prophage or prophage -like elements (Perna et al., 2001; Wick et al., 2005). The production of Shiga toxin and the ability to attach to and efface intestinal epithelial cells are two of the major virulence factors of the pathogen. Similar in structure and function to toxin produced by Shigella dysenteriae type 1 (O'Brien et al., 1983), Shiga toxin genes were horizontally transferred to E. coli by bacteriophage (O'Brien et al., 1984). Two major types of Shiga toxin are produced by STEC strains, Shiga toxin 1 (Stx1) and Shiga toxin 2 (Stx2) (Strockbine et al., 1986) with Stx2c being the most common variant of Stx2 found in O157 strains (Schmitt et al., 1991; Beutin et al., 2002). In addition to Shiga toxin, the locus of enterocyte effacement (LEE) encodes a number of virulence factors, including intimin and its receptor, Tir (Jerse et al., 1990; Deibel et al., 1998). Intimin is a product of the eae gene, which is responsible for the adherence to Tir that has been translocated into enterocytes in the membrane of the small intestine. Intimin-producing strains of enterohemorrhagic E. coli (EHEC) that were Shiga-toxin negative have been associated with causing bloody diarrhea in patients (Bielaszewska et al., 2008).
There are two primary approaches to studying the interaction of O157 with fresh produce. In one approach, defined environmental conditions may be applied in laboratory growth chambers, greenhouses, or hydroponic systems to gain an understanding of how O157 interacts with plants or soil. In the second approach, the interaction of O157 with plants or soils occurs in field studies where variable environmental conditions are encountered. A limitation of field studies is that virulent O157 cannot be intentionally introduced into the environment to contaminate fields. Hence, environmental E. coli O157:H7 Shiga toxin–deficient isolates were used in a field study (Erickson et al., 2010); however, the physiological response of such strains may be different from outbreak-causing virulent strains.
The purpose of this study was to delete the Shiga toxin genes and the eae gene producing intimin in F4546 (1997 alfalfa sprout) and K4492 (2006 lettuce) O157 outbreak strains to create O157 strains that could be used as surrogates for field studies. The mutant strains were labeled with a plasmid-containing green fluorescent protein (pGFPuv) for ease of detection during field- and growth-chamber studies. Finally, the mutant strains were compared to their wild-type, virulent strains for growth, Shiga toxin production, plasmid stability, attachment to leaf surfaces and soil particles, and survival in soil.
Materials and Methods
Bacterial strains
Bacterial strains and plasmids used in this study are listed in Table 1. Strains were streaked from frozen stock onto tryptic soy agar (TSA) (Neogen, Lansing, MI) and incubated at 37°C for 18 to 21 h. Single colonies from TSA plates were used to inoculate Luria Bertani broth for mutant construction or tryptic soy broth (TSB) for use in experimental studies to compare growth, attachment, and survival of wild-type and mutant strains. Growth media for strains containing pGFPuv were supplemented with 100 μg/mL ampicillin (Amp) (Roche Diagnostics, Indianapolis, IN).
pGFPuv, plasmid-containing green fluorescent protein; UGA CFS, Center for Food Safety, The University of Georgia.
Construction of mutant strains
The genes deleted in strain F4546 were stx 1, stx 2, and eae. The genes deleted in strain K4492 were stx 2, stx 2c, and eae (strain K4492 did not contain the stx 1 gene). Nucleotide deletions in the stx 1, stx 2, stx 2c, and eae genes were constructed using the Quick and Easy Gene Deletion kit (Gene Bridges, Heidelberg, Germany) by identifying 50 nucleotides adjacent to the upstream and downstream insertion sites of each gene to be deleted as well as nucleotides to amplify a neomycin-resistance cassette flanked by a flipase recognition target, FRT (Table 2). The PCR products generated were integrated into the strains by Red/ET recombination directed from plasmid pRedET(tet). Neomycin-resistance cassettes were resolved from the chromosome via flipase recombinase to generate nonpolar mutations (Buchholz et al., 1996; Zhang et al., 1998). EDL933 sequence, Genbank accession number AE005174, was used to determine the stx 1, stx 2, and eae regions for deletion in strain F4546 (Perna et al., 2001). The sequence of stx 1A, stx 1B, and the intergenic sequence between nucleotide 2996866 and 2995836 (complement) were deleted. The sequence between nucleotide 1352451 and 1353380 was deleted for stx 2A and stx 2B. Finally, the eae gene was deleted between nucleotide 4667711 and 4665776 (complement) to create strain MD42. TW14359 sequence, Genbank accession number CP001368, was used to determine the stx 2, stx 2c, and eae regions for deletion in strain K4492 (Kulasekara et al., 2009). The stx 2 A and B subunits, and the intergenic sequence between nucleotides 3226028 and 3226957 (complement), were deleted. The sequence between nucleotides 2695335 and 2696262 (complement) was deleted for stx 2c A and B subunits, and the intergenic space between. The eae gene was deleted between nucleotide 4657873 and 4655938 (complement) to construct strain MD41. A CaCl2 heat-shock method (Sambrook et al., 1989) was used to transform plasmid pGFPuv (Clonetech, Palo Alto, CA) containing genes to produce green fluorescent protein (GFPuv) and ampicillin resistance to each recombinant strain. The resulting strains MD46 (F4546 Δstx 1, Δstx 2, Δeae pGFPuv) and MD47 (K4492 Δstx 2, Δstx 2c, Δeae pGFPuv) produced bright green fluorescence under ultraviolet light (365 nm).
Underlined sequence is homologous to neomycin cassette template.
Detection of Shiga toxin
Wild-type and mutant pairs, F4546/MD42 and K4492/MD41, were cultured in Gram-negative broth (Thermo Fisher Scientific, Lenexa, KS) at 37°C for 18 to 24 h. In three replicate trials, diluted cultures of these isolates along with positive/negative controls were applied to The ImmunoCard STAT!® EHEC (Meridian Biosciences, Inc., Cincinnati, OH) to detect Shiga toxin 1 and 2.
Stability of pGFPuv in O157 strains
Mutant O157 strains MD46 and MD47 containing pGFPuv were grown in TSB with and without ampicillin (9 mL) at 37°C for 25 (trial two) or 30 days (trial one). After overnight culture, 10-μL portions were removed and either inoculated into fresh culture medium or applied to TSA-Amp plates for overnight incubation (37°C) and observation of fluorescent colonies under ultraviolet light (365 nm).
Comparison of growth curves for wild-type and mutant O157
Single colonies of wild-type/mutant pairs F4546/MD46 and K4492/MD47 (each strain contained pGFPuv) were inoculated individually into 3-mL TSB-Amp or M9 minimal salts broth (M9B) (Theofel et al., 2009) supplemented with 100 μg/mL ampicillin and incubated overnight at 37°C. Overnight cultures were diluted and approximately 100 colony-forming units (CFU) were added to 50 mL TSB-Amp or M9B-Amp. Cultures were incubated at 37°C with agitation (150 rpm), and sampled at 0, 1, 2, 4, 6, 8, 10, 12, and 24 h (TSB-Amp) or 0, 4, 6, 8, 12, 15, 24, 27, and 30 h (M9B-Amp). Samples were diluted accordingly and 100 μL was applied to TSA-Amp plates for enumeration. Three (TSB-Amp) and two (M9B-Amp) replicate trials were performed at separate times using strains reconstituted from the frozen state.
Culture of wild-type and mutant O157 for attachment and survival studies
Individual inocula were prepared for each isolate (F4546 pGFPuv, MD46, K4492 pGFPuv, and MD47) by thawing the isolates, streaking onto TSA-Amp, and incubating at 37°C for 21–24 h. Individual colonies from these plates were streaked onto a second TSA-Amp plate, incubated overnight, and single colonies from this second plate were used to inoculate 50 mL TSB-Amp that was incubated at 37°C with agitation (150 rpm) for 21–24 h. Cultures were sedimented by centrifugation (4193×g, 15 min, 4°C), washed twice in 0.1% peptone water, and resuspended in 0.1% peptone water to give≈9 log CFU/mL.
Comparison of wild-type and mutant O157 for leaf attachment
Green leaf lettuce leaves were aseptically removed from live plants (≈14-days post-transplantation) and held in 2 L of each isolate (6 log CFU/mL in sterile deionized water [SDW]) for 24 h at 4°C (Xicohtencatl-Cortes et al., 2009) with the stalk-end above the inoculum surface. Leaf samples were dried (30 min/side) prior to either analyzing immediately for total O157 on nonrinsed leaves or placing in a Whirl-Pak bag, swirling with 100 mL SDW for 1 min, and then enumerating for firmly attached O157 on rinsed leaves. Both nonrinsed and rinsed leaves (n=10 each) were macerated in a mortar and pestle with 0.1% peptone water (1:9, wt:vol). O157 were enumerated on TSA-Amp after 24-h incubation at 37°C. Three replicates were performed at separate times for this experiment using freshly prepared cultures.
Comparison of wild-type and mutant O157 for soil attachment
A Tifton Sandy Loam soil was obtained from produce fields in Tifton, GA. Moisture content of the soil was determined following drying at 100°C for 24 h and then water was added to give moisture contents ranging from 8.6% to 10.6%. Soil (5 g) was added to a glass funnel lined with a sterile #5 Whatman filter (GE Healthcare, Piscataway, NJ). Working inocula of each isolate (1 mL of 5 log CFU/mL in SDW) was applied to the center of the soil sample. After 30 min, SDW (25 mL) was applied four times to the contaminated soil to remove unattached organisms. Enumeration of attached O157 was determined by shaking filter and soil from each inoculated sample with 0.1% peptone water (1:9 wt:vol) for 30 s. Tenfold dilutions of this soil dispersion were made and aliquots applied to TSA-Amp plates for subsequent enumeration of fluorescent colonies. Four replicates were performed at separate times using freshly prepared cultures.
Comparison of wild-type and mutant O157 for survival in soil
Tifton soil (380 g) was spread over the bottom of four 3-L Gladware containers (Clorox Company, Oakland, CA) and sprayed with a mixture of either the virulent (F4546, K4492) or mutant (MD46, MD47) isolates (3.8 mL of 7 log CFU/mL in 0.1% peptone water). After mixing, the inoculated soil was acclimated for 24 h (20°C, 70% humidity, dark). Acclimated soil (17 g) was mixed with 153 g of noninoculated soil prior to storage in 532-mL plastic cups (Pactiv Corporation, Lake Forest, IL). Five cups of inoculated soil did not receive water (dry soil), whereas 80 mL of SDW was added to another five inoculated cups to maintain 5 mm of water on the surface of the soil (wet soil) prior to storing dry and wet soil samples at 20°C in the dark. Samples (10 g) were removed from each of the cups on days 0, 1, 2, 3, 4, and 7 and dispersed in 90 mL of 0.1% peptone water. After removing aliquots for enumeration of O157, 90 mL of 2x-TSB-Amp was added to the homogenized soil sample for overnight enrichment at 37°C. Aliquots of these enriched samples were streaked on TSA-Amp plates for overnight incubation at 37°C, after which the presence of fluorescent colonies revealed a positive result for the presence of O157 in the soil sample. Only one replicate trial was performed.
Statistical analysis
Enumeration data were log transformed prior to conducting statistical analyses (StatGraphics Centurion XVI, StatPoint, Inc., Herndon, VA). Growth rates from each of the wild-type/mutant pair trials were calculated from the linear portion of the logarithmic scale-curve and then subjected to the t-test. In addition, both enumeration and percent attachment data for each isogenic mutant pair were subjected to a t-test, whereas the survival in soil of wild-type and mutant isolates was subjected to analysis of variance (p≤0.05).
Results and Discussion
Construction of strains and detection of Shiga toxin
Surrogate strains of O157 associated with produce outbreaks and that had been traced to a production source were constructed in this study. The Gene Bridges system developed by Muyrers et al. (2001) was used to delete Shiga toxin and intimin genes from outbreak strains, F4546 and K4492. Two rounds of mutagenesis using the same oligonucleotides (Table 2) were performed on strain K4492 to delete stx 2 and stx 2c genes, since the sequence for stx 2 and stx 2c were 98% identical based on Blast alignment (Altschul et al., 1990). Individual deletions as specified above were confirmed by PCR and DNA sequencing of each strain (data not shown). Though intermediate steps during construction of MD42 and MD41 employed the use of antibiotic-resistant markers, antibiotic resistance was removed (based on sequencing data), resulting in triple mutant (stx1 and /or stx2, stx2c, eae) strains that did not retain these antibiotic resistance markers as evidenced by their inability to grow on agar containing the antibiotic (data not shown).
The wild-type/mutant pairs F4546/MD42 and K4492/MD41 were assayed for Shiga toxin 1 and 2 production using the Immunocard STAT!® EHEC. The wild-type strain, F4546, produced both Shiga toxin 1 and 2, whereas the mutant, MD42, produced neither (Fig. 1). The lettuce outbreak–associated stain, K4492, which lacks stx1 , did not produce a band corresponding to Shiga toxin 1 but did produce one corresponding to Shiga toxin 2. The mutant MD41, however, did not produce a band corresponding to Shiga toxin 2 (Fig. 1). Strain MD36, in which stx 2 was deleted from K4492 but stx 2c was not, did produce a band corresponding to Shiga toxin 2 according to the Immunocard STAT!® EHEC test, indicating that the test system detected both Shiga toxin 2 and 2c (data not shown).

Detection of Shiga toxin in wild-type and mutant O157.
Evaluation of plasmid stability in mutant O157 strains
Recombinant strains MD42 and MD41 were transformed with pGFPuv to create strains MD46 and MD47 to enhance the detection of these strains when inoculated into the rhizosphere and/or phyllosphere of fresh produce. As a measure of plasmid stability, pGFPuv in strain MD46 was maintained for 12 days in the absence of antibiotic, whereas pGFPuv in strain MD47 was maintained for 30 days. In the presence of ampicillin, pGFPuv was maintained in MD46 and MD47 for the duration of the trial (30 days). The difference in plasmid stability of these two isolates may be due to inherent physiological differences between the two outbreak isolates used in this study, similar to the variable stability observed for a broad host range plasmid, IncP-1, which had been incorporated into different microbial strains from the same genus or species (De Gelder et al., 2007) and to the wide range of stability shown by pGFPuv incorporated into different isolates of E. coli O157:H7 (Ma et al., 2011b). Confirmation that the plasmid remained in these recipient strains was validated in two field studies wherein strains MD46 and MD47 continued to be detected in and on lettuce plants 14 days after application to field-grown cutback lettuce shoots (Erickson et al., 2013) and in soil 39 days after application of contaminated compost (Erickson et al., 2014).
Growth kinetics of wild-type and mutant O157 strains
In this study, the isogenic pairs (K4492 pGFPuv/MD47 and F4546 pGFPuv/MD46) were subjected to growth in nutrient-rich (TSB-Amp) and minimal media (M9B-Amp). Cultures grown in TSB-Amp exhibited a lag phase as that culture was started with cells from an overnight culture that had entered the stationary phase. In contrast, no lag phase occurred for cultures grown in M9B-Amp as the cells used to start that culture were from an overnight culture that was still in the logarithmic phase. With both isogenic pairs, however, statistically similar logarithmic growth rates were observed in nutrient-rich and minimal media for the wild-type and isogenic mutant (Fig. 2, p>0.05). Hence, similar to other studies involving the loss of virulence factors or the insertion of the GFP-plasmid (Allison et al., 2007; Ma et al., 2011a), the surrogates generated in this study had fitness similar to that of the virulent isolates under nutrient-rich and nutrient-minimal growth conditions.
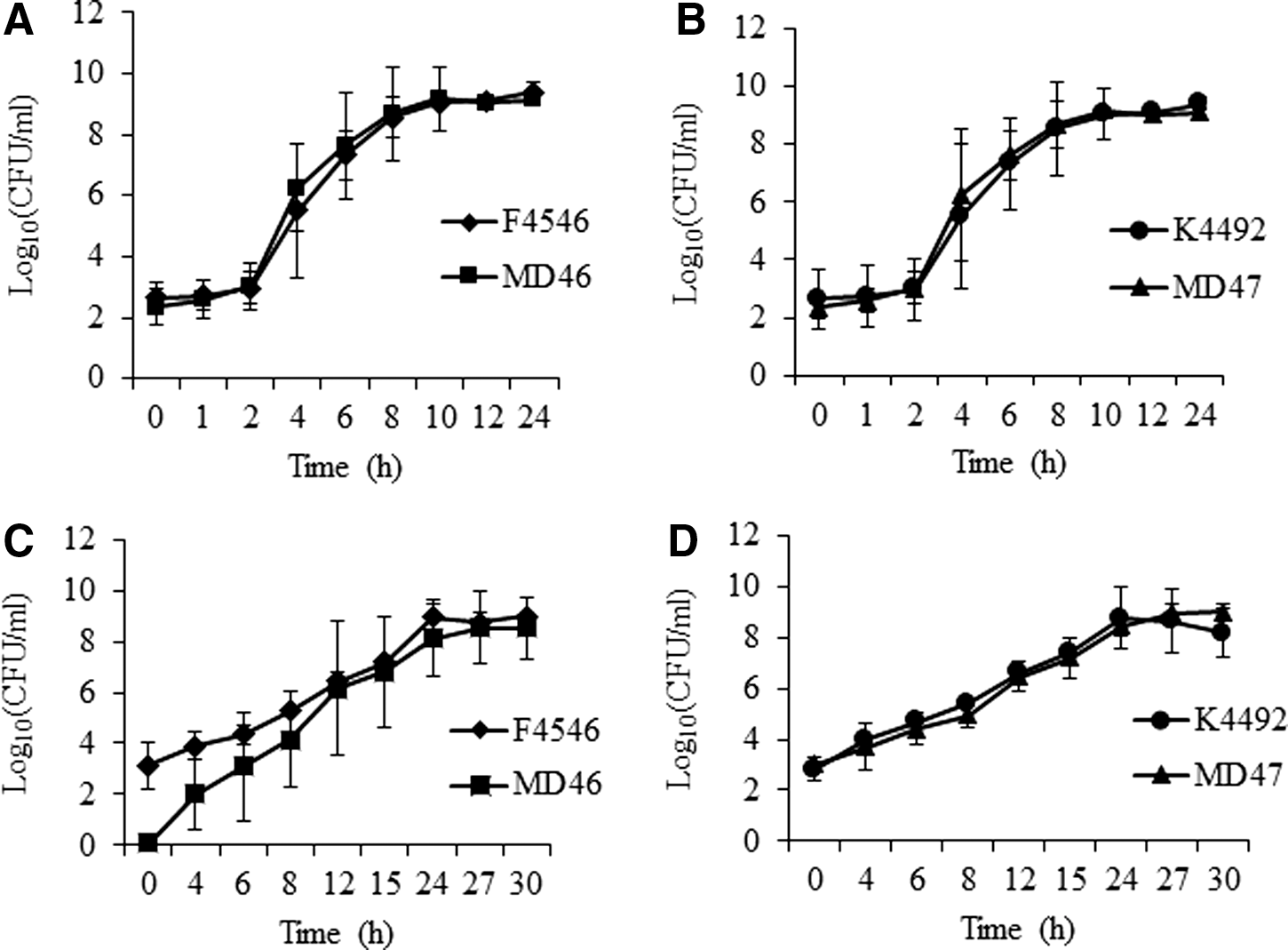
Growth curves of wild-type and mutant O157 and TSB-amp
Comparison of wild-type and mutant O157 for leaf and soil attachment
To ensure that genetic manipulation of the mutant strains did not disrupt innate processes that would affect their fate in environmental systems, studies were conducted to compare the attachment of wild-type and mutant strains to either plant tissue or soil. In the case of plant tissue, leaves were held in each inoculum type for an extended period (24 h) to ensure uniform exposure and sufficient time for attachment at a temperature (4°C) where growth would not occur. No significant differences in the total (nonrinsed leaves) or firmly attached (rinsed leaves) populations or in the percentage of total bacteria that were firmly attached to leaves were observed among the wild-type and mutant O157 strains (Table 3, p>0.05). While no differences in leaf attachment were observed between wild-type and mutant strains, it is acknowledged that intimin is important for attachment to human and animal cells (Lai et al., 2013) and therefore might potentially play a role in pathogen attachment to leaves. In any event, in the case of soil, both wild-type and mutant isolates had comparable degrees of attachment. Increased attachment by either F4596 or MD46 compared to K4492 or MD47 pair (Table 4), however, highlight that the degree of soil attachment can vary between different isolates.
Bacteria attached to leaf following soaking in inoculum for 24 h at 4°C, draining, and then drying.
Bacteria remaining on leaf tissue following soaking, draining, drying, and then swirling in a 10-fold volume of 0.1% peptone water.
(Number surface bacteria/number total bacteria)×100.
Mean log colony-forming units (CFU)/mL.
Mean log CFU/g after rinsing soil.
(Number attached bacteria/number bacteria applied)×100.
Comparison of survival in soil of wild-type and mutant strains
Wild-type and mutant O157 isolates had statistically similar survival rates in wet and dry soil systems over the course of a 7-day sampling period (Table 5, p>0.05). Survival of the wild-type and mutant pairs, however, was much greater in the wet soil system compared to the dry soil (Table 5). Previously, O157 was determined to alter gene expression when exposed to soil (Duffitt et al., 2011) or plant lysates (Kyle et al., 2010), implying metabolic adjustments to their environment to facilitate survival. The similar survival rates of the pairs of wild-type and mutant isolates suggests that the Shiga toxin, intimin, or GFP plasmid genes did not affect mechanisms associated with survival. This conclusion is supported by a previous study in which it was determined that a mutant of O157 EDL933, lacking stx and eae, and a pGFP harboring Pseudomonas aeruginosa GP41B had survival rates in soil systems comparable to their wild-type counterparts (Dandie et al., 2001).
(Log colony-forming units/g, mean±SD, or number positive by enrichment culture/total number samples analyzed).
The need for safe surrogates of O157 is vital to expand our understanding of how the organism survives in the environment. In this study, virulence genes were removed from two O157 outbreak isolates under the premise that such isolates may possess other attributes that facilitate their survival in fields. Given that growth and attachment to soil and plant surfaces and survival in soil was not affected by the genetic alterations, it is envisioned that these recombinant isolates would respond similarly to the virulent, wild-type strains and hence would be comparable surrogates to apply in field studies.
Footnotes
Acknowledgments
This work was supported by the United States Department of Agriculture NIFSI grant 2007-51110-03812. The authors thank Gaylen Uhlich for helpful technical discussions.
Disclosure Statement
No competing financial interests exist.