
Abstract
Nonthermal technologies are being investigated as viable alternatives to, or supplemental utilization, with thermal pasteurization in the food-processing industry. In this study, the effect of ultraviolet (UV)-C light on the inactivation of seven milkborne pathogens (Listeria monocytogenes, Serratia marcescens, Salmonella Senftenberg, Yersinia enterocolitica, Aeromonas hydrophila, Escherichia coli, and Staphylococcus aureus) was evaluated. The pathogens were suspended in ultra-high-temperature whole milk and treated at UV doses between 0 and 5000 J/L at a flow rate of 4300 L/h in a thin-film turbulent flow-through pilot system. Of the seven milkborne pathogens tested, L. monocytogenes was the most UV resistant, requiring 2000 J/L of UV-C exposure to reach a 5-log reduction. The most sensitive bacterium was S. aureus, requiring only 1450 J/L to reach a 5-log reduction. This study demonstrated that the survival curves were nonlinear. Sigmoidal inactivation curves were observed for all tested bacterial strains. Nonlinear modeling of the inactivation data was a better fit than the traditional log-linear approach. Results obtained from this study indicate that UV illumination has the potential to be used as a nonthermal method to reduce microorganism populations in milk.
Introduction
S
Thermal pasteurization constitutes the most extensively used and approved method for the inactivation of milkborne pathogens. Thermal pasteurization used to make milk safe for consumption has associated with it high energy costs, can denature proteins, and has associated issues with flavor degradation. Many nonthermal technologies, such as the germicidal portion of ultraviolet light (UV-C) ranging from 200 to 280 nm, are now being investigated as an alternative to thermal treatment of milk to inactivate pathogens and improve shelf life (Bintsis et al., 2000; Sastry et al., 2000; Smith et al., 2002; Sudhirk, 2004; Matak et al., 2005; Krishnamurthy et al., 2007; van Wyk and Gouws, 2011; Rossitto et al., 2012). UV technology has the advantage of being chemical free, producing less waste, and being more energy efficient than the traditional thermal pasteurization process. UV-C treatment using low-pressure mercury lamps with the wavelength of 253.7 nm has been approved by the U.S. Food and Drug Administration (USFDA) for water and juices (Anonymous, 2000; USFDA, 2000). UV-C light at 253.7-nm wavelength has optimal germicidal properties for the inactivation of microorganisms including bacteria, protozoa, yeasts, molds, algae, fungi, and viruses. UV-C light is absorbed by the DNA, causing a cross-linkage of neighboring pyrimidine nucleoside bases (thymine and cytosine) in the DNA strand. Dimerized nucleotide damage in DNA renders the strand unreplicable, leading to cell death (Bolton, 2010).
One disadvantage of UV light as a microbial inactivation method is that organisms must be directly exposed to the UV-C to absorb UV photons. When a liquid is opaque or turbid, such as milk, UV-C photons cannot penetrate deeply into the liquid. The effective penetration depth for milk at 254 nm is approximately 0.003 cm (Koutchma, 2009b). A UV system that is designed to bring every particle of milk to the surface for UV exposure will improve the efficacy of UV microbial inactivation.
UV-C light treatment is a potential nonthermal alternative or enhancement to the high-temperature short-time pasteurization process. FDA has a 5-log pathogen reduction requirement in 21 CFR 120.24 (USFDA, 2011) that describes their requirement for the minimum level of pathogen killing required for a technology to achieve in making a food product nonhazardous. The first objective of this study was to determine processing conditions and the dose of UV-C light necessary to achieve a 5-log reduction of milkborne pathogens in experimentally inoculated ultrahigh-temperature (UHT) whole milk. The second objective was to use nonlinear modeling methods to assess parameters of pathogen UV inactivation.
Materials and Methods
Bacterial strains
All seven cultures were obtained from the American Type Culture Collection (ATCC, Manassas, VA). E. coli O157:H7 ATCC# 43888, S. enterica subsp. enterica serovar Senftenberg ATCC# 43845, S. aureus subsp. aureus ATCC# 29213, Y. enterocolitica subsp. enterocolitica ATCC# 9610, S. marcescens subsp. marcescens ATCC# 13880, and A. hydrophila ATCC# 7966 were reconstituted and grown at 37°C in tryptic soy broth (TSB; Hardy Diagnostics, Santa Maria, CA). L. monocytogenes ATCC# 43256 was reconstituted and grown at 30°C in TSB. All cultures were stored in glycerol (20% vol/vol) and TSB at −80°C until use.
Growth of bacterial strains and milk inoculation
Each strain culture was twice transferred to TSB and sheep blood agar (Hardy Diagnostics). Plates for each strain were incubated at 37°C for 18–24 h with the exception of Listeria, which was incubated at 30°C for 48 h. Bacterial growth was scraped from the agar surface into Butterfield's phosphate-buffered water (BB) (USFDA, 2014). Bacterial levels were determined using turbidity measurements at 620 nm with a spectrophotometer (UV-1201S; Shimadzu, Kyoto, Japan) and a conversion value of 0.02 optical densities being equal to 1×107 colony-forming units (CFU)/mL (7 logs CFU/mL). The suspension was adjusted to a concentration of 109 CFU/mL (9 logs CFU/mL) in BB. For inoculation into the 20 L of UHT pasteurized whole milk (purchased from Advanced Food Products, Visalia, CA) at a final concentration of approximately 106–107 CFU/mL (6–7 logs CFU/mL), the culture was added directly to milk via syringe into an in-line aseptic sampling port. Prior to UV treatment, the UV unit was run for 5 min to allow adequate mixing of the bacteria in the milk. Background populations of microorganisms were not detected in any of the test UHT milk prior to inoculation.
UV-C processing of UHT milk
Milk was exposed to UV-C light using a thin-film turbulent flow system designed and manufactured by SurePure, Milnerton, South Africa (Keyser et al., 2008; Simmons et al., 2012) (Table 1). The UV system consisted of four sequential UV-C turbulator chambers. A turbulator chamber was a 316-grade stainless steel corrugated spiral tube, which contained a quartz sleeve encasing a single germicidal low-pressure mercury UV lamp (Spec #2364, First Light, Poultney, VT) (100-W lamp wattage, 29W UV-C output, 90% at 254 nm) (Fig. 1). The flow rate of 4300 L/h achieved a turbulent flow (Reynolds Value (Re)>7500). The 4 lamps delivered a UV-C dose of 100 J/L (Keyser et al., 2008) to the milk being treated after 1 pass through the system. A closed-loop piping system was created for processing (Fig. 2) multiple passes to achieve higher doses of UV energy.
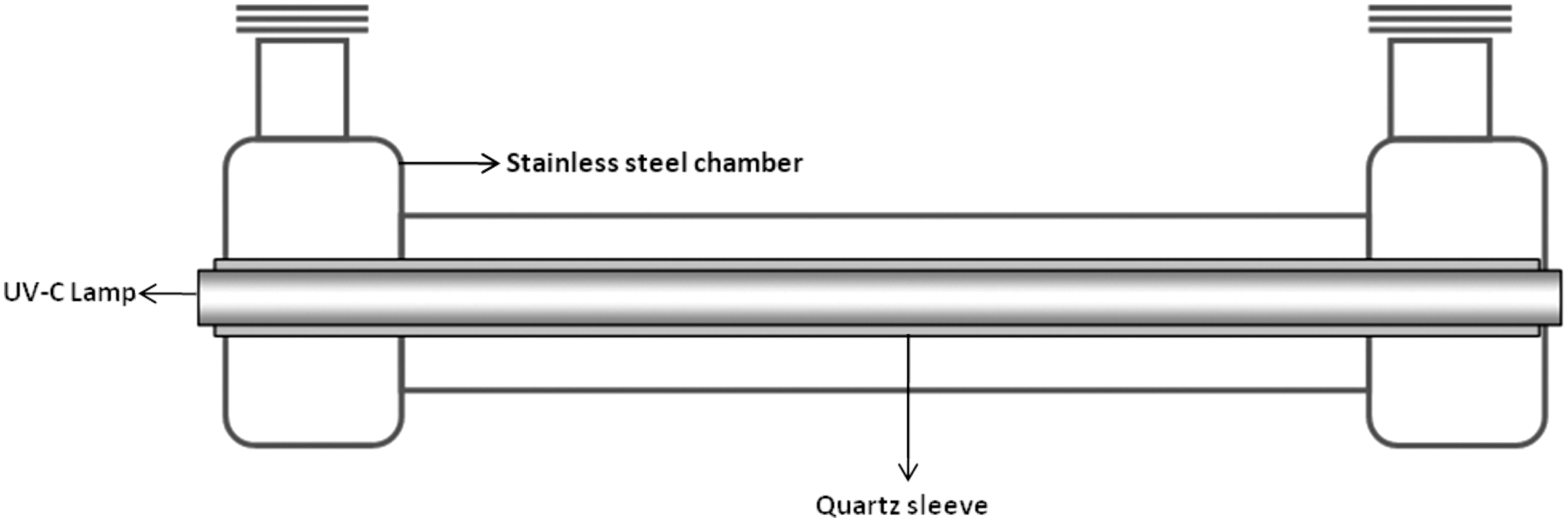
Cross section of a SurePure ultraviolet (UV) treatment system turbulator™ (used with the permission of SurePure Inc.).
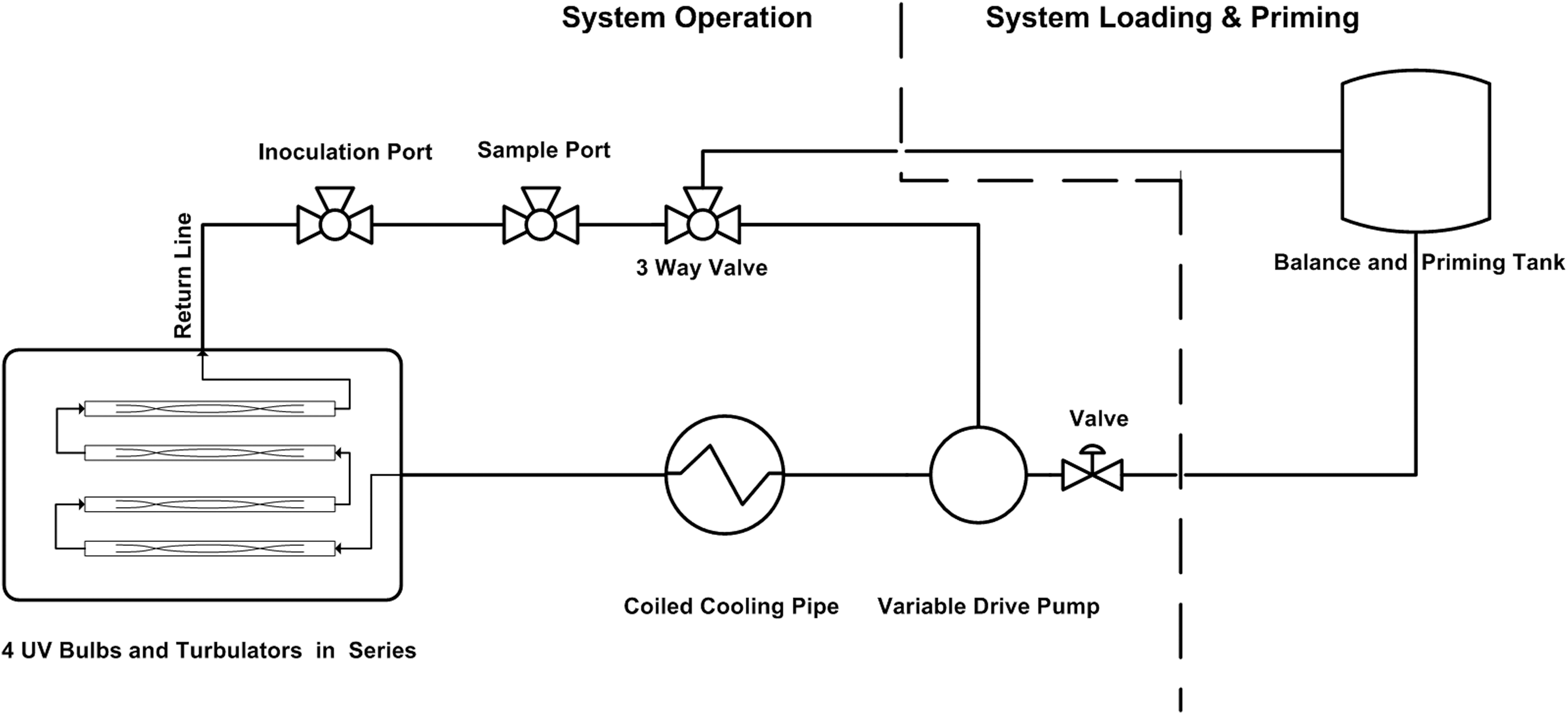
Schematic drawing of the pilot-scale SurePure ultraviolet (UV) treatment system containing four UV-C lamps.
Chilled milk (4°–6°C) was pumped from a feed tank using a multistage centrifugal feed pump (Inoxpa, Banyoles, Spain) into the system. The time needed to pass 20 L of product through the system once was 16.4 s. Samples were taken via syringe from an aseptic sampling port at specific time points throughout the UV treatment run correlating to 0, 33, 82, 115, 164, 197, 247, 329, 411, 493, 575, and 822 s or a calculated cumulative UV dose of 0, 200, 500, 700, 1000, 1200, 1500, 2000, 2500, 3000, 3500, and 5000 J/L, respectively (Table 2).
Microbial analysis
Milk samples were 10-fold diluted and 0.10-mL spread plated to tryptic soy agar (TSA) and incubated at appropriate time and temperature for each bacterial strain as stated above. For later time points, where samples were projected to be <100 CFU/mL, undiluted samples were spread plated (1 mL across 4 plates) on TSA for more precise counts. The colonies were counted to determine the colony-forming units per milliliter. The untreated (control) sample consisted of milk recirculated in the UV unit for 5 min before inoculation of target organism.
Modeling analysis
The surviving microbial counts (colony-forming units per milliliter) were log-transformed and plotted as a function of cumulative UV dose. Modeling of the data was performed with GInaFiT (version 1.5), the freeware add-in for Microsoft® Excel (Geeraerd et al., 2005). Experimental points were the means of three experiments conducted on independently grown cultures. Eight inactivation models were tested for each pathogen (Table 3). The root-mean-square errors (RMSE) and R
2 values were used to evaluate the performance of each model (McKellar and Lu, 2004; Ratkowsky, 2004; Oteiza et al., 2010). The model with the smallest RMSE was considered the best fit. Using the RMSE as the selection criteria, the Mixed Weibull model (Coroller et al., 2006) was selected for final modeling of the data. The mixed Weibull model is:
where N represents the microbial cell density (colony-forming units per milliliter) after treatment, N0 is the initial microbial cell density (colony-forming units per milliliter), t is the cumulative UV dose at a specific time, p is the shape parameter, α is the log proportion between the sensitive fraction (f) and the resistant one (1-f), and δ1 and δ2 are the UV dose for the first decimal reduction of the subpopulation 1 and subpopulation 2, respectively. When p>1 a convex curve is observed; when p<1 a concave curve is observed, and when p=1 a linear curve is observed.
Results
Background populations of microorganisms were not detected in any of the test UHT milk prior to inoculation. The average initial populations of the tested pathogens inoculated into the bovine UHT milk ranged from 5.64 to 7.34 logs CFU/mL. Using the RMSE values calculated for each model as an indicator of goodness-of-fit, the mixed Weibull model was the best fit for the UV-C inactivation data (Table 3). The coefficients of goodness-of-fit determination (R 2) and the RMSE values for the mixed Weibull model ranged between 0.9902–0.9994 and 0.0579–0.2331, respectively (Table 4). The 4-log and 5-log reduction values for the mixed Weibull model were determined from the identified log10(N) and its corresponding UV dose generated by the GInaFiT tool (Table 4).
p, shape parameter; δ1, ultraviolet dose for the first decimal reduction of the subpopulation 1; R 2, coefficient of determination; RMSE, root-mean-square error.
The fitted mixed Weibull model inactivation curves are shown in Figure 3a–g. Microbial counts of bacteria decreased as the cumulative dose of UV-C light increased. The shape of the inactivation curves varied depending on the UV-C light resistance of each bacterial species tested. The general inactivation pattern displayed for the pathogens tested showed a sigmoidal inactivation curve. For L. monocytogenes (Fig. 3d), S. marcescens (Fig. 3f), Salmonella Senftenberg (Fig. 3e), and E. coli (Fig. 3c), there was an initial shoulder as evidenced by the Weibull function δ1, the energy required to create the first decimal reduction of bacteria, which ranged from 736 J/L to 546 J/L. This was followed by exponential cell death followed by a shallower sloping-tail where the inactivation rate drastically decreases. The 3 organisms with the least shouldering and the lowest δ1 were A. hydrophila (Fig. 3a), S. aureus (Fig. 3g), and Y. enterocolitica (Fig. 3b), which were 53%, 58%, and 44% relative to L. monocytogenes (Fig. 3d).
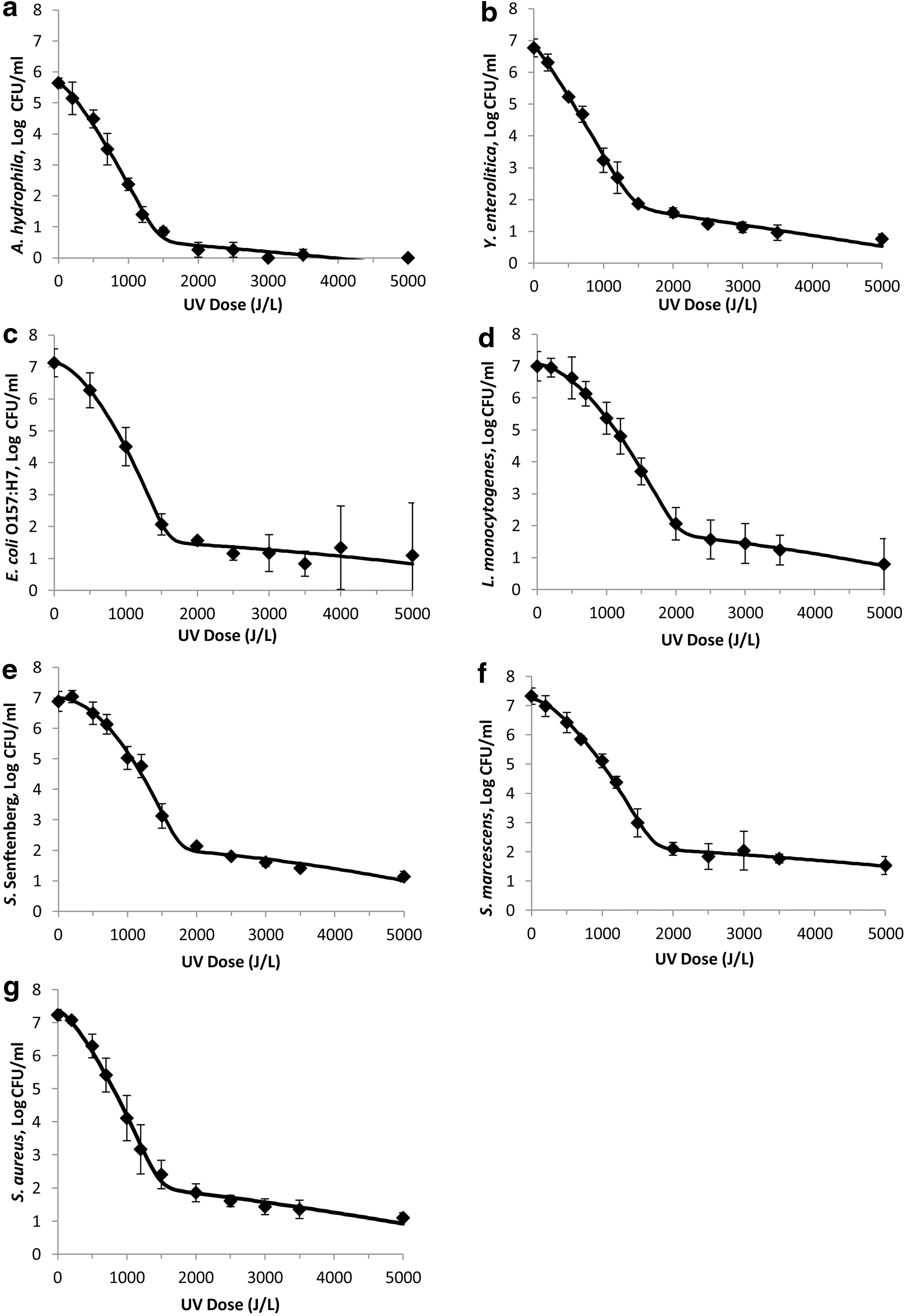
Survival curves of pathogens fitted with the mixed Weibull model
The 5-log UV-C light-inactivation values for the targeted bacterial pathogens displayed a wide range. L. monocytogenes was the most resistant to the UV treatment among seven pathogenic organisms tested (Table 4). It achieved a 5-log reduction at 2000 J/L of energy. S. aureus was the most sensitive to the UV treatment, by achieving a reduction of 5-logs with 1450 J/L of energy. All of the models had a p parameter value >1 making the shape of the curves convex.
Discussion
UV light is a promising technology for the destruction of pathogenic microorganisms in milk. This study was able to achieve 5-logs of inactivation in all of the pathogens tested (Table 4), which meets the criteria set forth by the U.S. Food and Drug Administration. The results of this work show that the sensitivity to UV-C of milkborne pathogens belonging to different genera varied significantly. Among the tested organisms, L. monocytogenes proved to be the most resistant to UV-C, whereas S. aureus was the most sensitive (Table 4). For A. hydrophila, which had a lower initial microbial load (Fig. 3a), the UV inactivation dose may be higher than reported here. In looking at the effect of atmospheric plasma inactivation of Bacillus subtilis spores on membrane surfaces, Deng et al. (2005) demonstrated that an 80% increase in time was required for a 3-log reduction for 109 spores as compared to 106. There was no clear trend in UV resistance between Gram-positive and Gram-negative organisms, which is in contrast to other studies (Shama, 1999; Sastry et al., 2000). Differences in UV equipment designs, bacterial species and strains, and processing conditions make it difficult to compare results across the literature (Smith et al., 2002; Matak et al., 2005; Krishnamurthy et al., 2007; Engin and Karagul-Yuceer, 2012). All of these studies do agree that turbidity of the liquid, UV penetration depth, UV treatment time, UV dose, and turbulent flow are important parameters that can influence the reduction of microorganisms. Important parameters of product characteristics that could influence UV effectiveness include solids content, transparency, homogeneity of the product, surface geometry, and depth of fluid column (National Advisory Committee on Microbiological Criteria for Foods, 2006).
Despite the growing interest in applications of nonthermal technologies in dairy processing, there has been very limited research of UV treatment of bovine milk. Using the CiderSure 3500 (continuous UV-C light; turbulent flow-through system) UV apparatus, Matak et al. (2005), was able to achieve a 5-log reduction of L. monocytogenes in goat's milk with a UV dose of 15.8 mJ/cm2. The study performed by Reinemann et al. (2006), using the PureUV system (PureUV, Milnerton, South Africa) (continuous UV light, turbulent flow-through system), achieved a 3-log or greater reduction of total bacteria, coliforms, psychrotrophic bacteria, and thermoduric bacteria in raw cow's milk at a UV dose of 1.5 kJ/L. Using a static, pulsed excimer UV-laser system (COMPEX-110, Lambda Physics, Grenoble, Germany), Smith et al. (2002) achieved a 2-log reduction of S. marcescens in raw bovine milk with pulsed UV at 248 nm using a dose of 12.6 J/cm2. Bandla et al. (2012) reported that UV-C treatment of raw cow's milk with 16.82 mJ/cm2 in the coiled tube UV reactor was capable of reducing standard plate count by 2.3-log CFU/mL. Innocente et al. (2014) achieved a 3.2-log reduction of total microbial counts in raw milk using a static, pulsed-light xenon lamp system (Clarinor, Rouaine, France) at a dose of 26.25 J/cm2.
The UV-C test unit used in this study had a flow rate of 4300 L/h, a flow rate that can be found in commercial processing systems. UV treatment of milk, in its initial phases of industrial implementation, could be used as an adjunct to thermal pasteurization at the point of bottling to eliminate any microorganisms that survived heat treatment and postpasteurization growth that might occur. UV-C treatment postpasteurization has been shown to increase shelf life of fluid 2%-fat and 4%-fat milk from 7 to 35 days (Rossitto et al., 2012). Utilization of UV treatment as a replacement to thermal pasteurization still has many regulatory steps to address regarding its application, but if utilized would be a green technology with significant energy savings.
Light in the visible and UV spectrum can cause oxidation of protein and lipids in milk that lead to “light oxidation,” causing flavor defects. Lipid oxidation is distinct from and a different chemistry than fatty acid hydrolysis. Fatty acid hydrolysis leads to rancidity. Sensory analysis in Rossitto et al. (2012) demonstrated that UV-C treatment of pasteurized cow's milk at 880 and 1760 J/L could be distinguished from untreated milk in a triangle test for difference. Matak et al. (2007) reported some panelists used aroma descriptors of “barnyard” and “goaty” for raw goat milk treated with 16 J/cm2 for 18 s. Cilliers et al. (2014) reported a “tallowy” descriptor of aroma and flavor of raw cow's milk treated with a UV-C dose of 1045 J/L. These descriptors of “cardboard” and “tallowy” have also been reported for heat-treated milk (Lee and Min, 2009). Any sensorial defects that might be associated with the specific low-power UV-C equipment used in this investigation at 2000 J/L (the dose required to kill 5 logs of the most UV-resistant pathogen, L. monocytogenes) will require further studies.
Despite the long-standing use of the log-linear kinetics approach for thermal inactivation in the food industry, there has been increasing evidence that inactivation of microorganisms do not follow first-order kinetics, especially for nonthermal processes (Qualls and Johnson, 1983; Shama, 1999; Sastry et al., 2000; Wright et al., 2000; Quintero-Ramos et al., 2004; Matak et al., 2005; Peleg, 2006; Keklik et al., 2012). Nonlinear shapes can be explained by “multi-hit kinetics,” which is the result of the cumulative effect of UV dose (Heldman and Newsome, 2003; Quintero-Ramos et al., 2004; Chevrefils et al., 2006). The shoulder can be explained as bacteria that can survive low UV doses due to photorepair ability. Photoreactivation is an enzyme process that cleaves the pyrimidine dimers to reverse the damage from the exposure of UV-C light. With increasing UV-C exposure, DNA is damaged beyond the cell's capability to repair the damage, leading to cell death (Bolton, 2010). The mixed Weibull model, which was the model that best fit the inactivation curves for all the pathogens tested here (Fig. 3a–g), assumes the existence of two subpopulations, one more sensitive and one more resistant (Coroller et al., 2006). When the critical UV dose is achieved, the curve shows a straight line. As the inactivation rate approaches the initial inoculum level, the rate of killing reduces as more UV light is applied. This is the tailing portion of the dose–response curve. The resistance of each organism will influence the shape of the curve. More resistant organisms will display more pronounced shoulder and tails. The shouldering phenomenon may also be a result of DNA repair mechanisms (Koutchma et al., 2009a; Gayan et al., 2012), which was not tested for in this study.
Forcing a straight line through a nonlinear curve may result in an overestimate or underestimate of the organism resistance, which may lead to overprocessing or underprocessing food (Peleg, 1999). By overprocessing, flavor and nutritional quality could be jeopardized, and by underprocessing, safety could be jeopardized. Nonlinear survivor curves have led to alternative kinetics models for a more accurate assessment of the data observed (van Boekel, 2002).
Conclusions
In summary, UV-C light was found to be an effective nonthermal technology that can reduce microbial populations in UHT bovine whole milk. L. monocytogenes was found to be the most resistant organism among the seven organisms tested in the study. Pathogen reduction of greater than 5-logs is possible with 1450–2000 J/L of UV irradiation in combination with thin-film turbulent flow of milk, and therefore may hold promise as an alternative technology to heat pasteurization. A limitation of this study was that use of single strains of microorganisms might not reflect the variability that may exist among different strains. Further studies are warranted to consider the varying UV-C resistance in different strains of the same organism. Future studies pertaining to sensorial and chemical properties of milk exposed to UV-C are also needed.
Footnotes
Acknowledgments
This work was funded by the California Dairy Research Federation. We thank Kristina Maloy, Kandi Gist, and Jacob Koontz of the Dairy Food Safety Laboratory at the Veterinary Medicine Teaching and Research Center for their technical assistance.
Disclosure Statement
The Dairy Food Safety Laboratory (Dr. James S. Cullor) had kindly received the loan of the SP4 unit from SurePure that was used to conduct the research described in this paper. In addition, SurePure in 2009 donated an unrestricted gift to the University of California. SurePure did not provide any direct funding for the conduct of this research. In addition, SurePure did not participate in the design or conduct of the research described in this paper. These donations did not influence or bias the performance of the research described in this manuscript in any way.
No competing financial interests exist.