
Abstract
The emergence of antibiotic-resistant foodborne Salmonella has become a major public health problem. Consumption of undercooked poultry contaminated with Salmonella can induce food poisoning in humans. In this study, we investigated the occurrence and antibiotic resistance patterns of Salmonella spp. isolated from 120 chicken carcasses produced in 6 poultry slaughterhouses in South Korea. A total of 11 samples (9.2%) were found contaminated with Salmonella: 5 isolates were serotyped as Salmonella Bellevue strain (slaughterhouse C) and 6 isolates were serotyped as Salmonella Enteritidis strain (slaughterhouse E). Salmonella Bellevue isolates were resistant to five antibiotics (ampicillin, chloramphenicol, nalidixic acid, tetracycline, and trimethoprim/sulfamethoxazole), while Salmonella Enteritidis isolates were resistant to nine antibiotics (ampicillin, cefotaxime, ceftazidime, cefazolin, cephalothin, amikacin, nalidixic acid, streptomycin, and tetracycline). All cephalosporin-resistant Salmonella Enteritidis isolates exhibited the extended-spectrum β-lactamase (ESBL) phenotype and carried the gene encoding CTX-M-15, the most prevalent ESBL enzyme worldwide. Based on molecular subtyping performed using the automated rep–polymerase chain reaction (PCR) system (DiversiLab), the isolates showing ≥95 similarity in their rep-PCR banding patterns were classified into 5 pulsotypes. Given that cephalosporins are the drugs of choice for invasive Salmonella infections, the high incidence of ESBL-producing strains in chicken should emphasize the necessity of regular monitoring of the occurrence of antibiotic-resistant ESBL-positive Salmonella strains in poultry meat.
Introduction
S
The emergence of antibiotic-resistant strains among various pathogenic bacteria has become an important health concern throughout the world (Miranda et al., 2009). Extensive use of antibiotics in animal husbandry has been suggested as the one of the important causes of antibiotic resistance observed among pathogenic bacteria isolated from humans, which makes human salmonellosis more difficult to treat (Fluit, 2005; Hyeon et al., 2011). The overuse and abuse of diverse antimicrobial agents underlie the emergence of multidrug resistant (MDR) Salmonella reported in many countries (Fluit, 2005; Kim et al., 2012; Wu et al., 2013). Therefore, the patterns of antibiotic resistance in Salmonella strains from poultry should be monitored periodically.
Extended-spectrum β-lactamases (ESBLs), which are β-lactamases with the ability to hydrolyze penicillins and most of the cephalosporins, are synthesized by an increasing number of Enterobacteriaceae (Zhao et al., 2009; Wu et al., 2013). ESBLs are less prevalent in Salmonella than in other Gram-negative Enterobacteriaceae members, including Escherichia coli and Klebsiella and Proteus spp. (Zhao et al., 2009). The resistance to cephalosporins and fluoroquinolones, which are the drugs of choice to treat clinical salmonellosis, limits available therapeutic options (Bertrand et al., 2006; Hyeon et al., 2011). Furthermore, extended-spectrum cephalosporins are preferentially used to treat children because of potential secondary effects of fluoroquinolones on cartilage development (Fey et al., 2000; Lee et al., 2003; Bertrand et al., 2006). The ESBL-producing Salmonella might be transmitted to humans from animals via contact or consumption of contaminated food, thus complicating the treatment (Carattoli, 2008; Liebana et al., 2013; Clemente et al., 2013). Although the prevalence and antimicrobial resistance of Salmonella from poultry samples in South Korea have been reported in previous studies (Woo, 2007; Hyeon et al., 2011; Kim et al., 2012), limited data are available for their ESBL characteristics.
The aim of this study was to investigate Salmonella contamination in chicken carcasses collected from six poultry slaughterhouses located in different regions of South Korea. Salmonella isolates were analyzed based on their antibiotic-resistance spectra, and cephalosporin-resistant strains were further characterized by ESBL genotypic and phenotypic patterns. Additional molecular subtyping of Salmonella isolates was performed using an automated rep–polymerase chain reaction (PCR) system (DiversiLab) to identify the relatedness among the isolates.
Materials and Methods
Sample collection
A total of 120 broilers were collected between February and April 2014, from 6 poultry slaughterhouses (20 samples each) each located in a different province of South Korea (Kyunggi, Chungbuk, Seoul, Chungnam, Cheonbuk, Kyungbuk: slaughterhouses A–F). The chicken carcasses were collected on the same day in each plant. The fully processed broiler samples were collected from the storage step. The broilers were individually packaged; they were transported to the laboratory on ice within 1 day and stored at 4°C. All samples were used for experiment within 24 h.
Isolation of Salmonella from chicken carcasses
All experimental procedures for the isolation of Salmonella were performed according to our previous study (Hyeon et al., 2012) with minor modification. Briefly, whole chicken carcasses were rinsed with 400 mL of buffered peptone water (BPW; Oxoid, Hampshire, UK) with gentle shaking for 1 min; 25 mL of the rinsate was combined with the same volume of BPW and incubated at 37°C for 24 h. For selective enrichment, 0.1 mL of enrichment was transferred to 10 mL of Rappaport-Vassiliadis (RV) broth (bioMérieux, Marcy l'Etoile, France) and incubated at 42°C for 24 h. A loopful of RV broth culture was seeded onto xylose lysine deoxycholate agar (BD Difco, Detroit, MI) and incubated at 37°C for 24 h. A maximum of three presumptive colonies were subcultured onto nutrient agar (BD). The suspected colonies were screened by the urease test and confirmed by the Vitek 2 GN kit (bioMérieux).
Salmonella serotyping
Serotyping was performed using a commercial antisera kit (BD). The somatic O antigen was identified by the slide agglutination test. The flagella antigen was detected by inoculation of the tested rinsates into GI motility agar (BD) followed by inoculation into veal infusion broth (BD); overnight cultures were fixed with 0.6% formalin and analyzed by the tube agglutination test.
Antibiotic susceptibility testing
The disc-diffusion method was used to determine antibiotic susceptibility of the isolated strains. A single colony of each strain grown on nutrient agar at 37°C for 24 h was inoculated into Mueller-Hinton broth (Oxoid) and incubated for 5 h at 37°C. The culture suspension was adjusted to 0.5 MacFarland turbidity and spread on Mueller-Hinton agar with a sterile cotton swab. Antibiotic-containing discs (from Oxoid and BD) were placed on MHA with dispenser (Oxoid) followed by incubation at 37°C for 18 h. The following antibiotics were tested: ampicillin (10 μg), amoxicillin–clavulanic acid (30 μg), amikacin (30 μg), gentamicin (10 μg), streptomycin (10 μg), tetracycline (30 μg), chloramphenicol (30 μg), cefoxitin (30 μg), cephalothin (30 μg), cefazolin (30 μg), cefotaxime (30 μg), ceftazidime (30 μg), sulfamethoxazole-trimethoprim (25 μg), nalidixic acid (30 μg), ciprofloxacin (5 μg), norfloxacin (10 μg), and imipenem (10 μg). Escherichia coli ATCC 25922 was used as the reference strain in all the cases. According to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2013), the isolates were considered susceptible, intermediate, or resistant.
ESBL detection and antimicrobial susceptibility
ESBL production was determined by the phenotypic confirmatory disc-diffusion method recommended by the CLSI. Briefly, ceftazidime and cefotaxime discs with or without clavulanic acid (BD) were placed on the same MHA plate. ESBL production was determined by comparison of the inhibition zone diameters: on the clavulanic acid–containing discs, ESBL-positive strains exhibited the inhibition zone 5 mm wider than that on the corresponding disc without clavulanic acid (Wu et al., 2013).
ESBL production was also examined by determining the minimum inhibitory concentration (MIC) for 11 β-lactam antibiotics: ampicillin (AMP), amoxicillin/clavulanic acid (AMC), piperacillin/tazobactam (TZP), cefazolin (KZ), cefoxitin (FOX), cefotaxime (CTX), ceftazidime (CAZ), cefepime (FEP), aztreonam (AZT), ertapenem (ETP), and imipenem (IMP), using the Vitek AST-N224 kit (bioMérieux).
Characterization of the β-lactamase-encoding genes
The presence of genes encoding different types of β-lactamases (Temoniera [TEM], sulphydryl variable [SHV], and cefotaxime [CTX-M]) was confirmed by PCR and analyzed by DNA sequencing as described previously (Batchelor et al., 2005; Rayamajhi et al., 2008). Among the CTX-M-positive isolates, the bla CTX-M gene group was identified using the primers specific for bla CTX-M-1 and bla CTX-M-9. Sequence analyses and comparisons with known sequences were performed using the BLAST programs at the National Center for Biotechnology Information website (NCBI, 2014).
Rep-PCR DNA fingerprinting with DiversiLab
Salmonella strains were incubated on nutrient agar overnight. Genomic DNA was extracted using the UltraClean Microbial DNA Isolation kit (MoBio Laboratories, Solana Beach, CA) and DNA concentration was quantified using a NanoDrop 2000 UV spectrophotometer (Thermo Scientific, Wilmington, DE). The isolated genomic DNA was used as a template for rep-PCR amplification with the DiversiLab Salmonella kit (bioMérieux); the amplified products were placed in a microfluidics chip and analyzed using a 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA). Dendrograms and subtyping patterns were created by determination of phylogenetic distance by using the unweighted-pair group method with arithmetic mean. Subtyping patterns were analyzed and statistically compared by Pearson correlation using the DiversiLab software. With reference to Healy et al. (2008) and Hyeon et al. (2011), genetically similar isolates were defined as having >95% similarity.
Results and Discussion
Salmonella contamination in chicken carcasses
Salmonella incidence in chicken carcasses is presented in Table 1. Salmonella spp. were found in two of six poultry slaughterhouses (C and E; 33.3%) and isolated from 11 of 120 chicken carcass samples (9.2%). Six strains from slaughterhouse E were identified as serovar Enteritidis and five strains from slaughterhouse C were identified as serovar Bellevue (Table 1).
The Salmonella occurrence in poultry (9.2%) observed in this study was lower than the previously reported rates of 15.5–63.3% (Woo, 2007; Bae et al., 2013; Choi et al., 2014). This discrepancy might have resulted from the differences in detection methods, investigation time, and country-specific hygiene and sanitation standards (Kim et al., 2012; Choi et al., 2014). Furthermore, the reduced incidence of Salmonella may be related to the policy of individual packaging of every poultry carcass institutionalized by the Korean government since January 2011 to reduce contamination of pathogens on poultry meat (Kim et al., 2012).
Previous studies have found that Salmonella Enteritidis is the most frequently reported Salmonella serovar detected in retail chicken meat and poultry slaughterhouses in South Korea (Chung et al., 2003; Woo, 2007; Kim et al., 2012), which is consistent with our findings.
Antibiotic resistance
The antibiotic susceptibility patterns of Salmonella isolates are summarized in Table 2. Among the 11 isolates, 6 strains from slaughterhouse E were resistant to 9 antibiotics (ampicillin, cefotaxime, ceftazidime, cefazolin, cephalothin, amikacin, nalidixic acid, streptomycin, and tetracycline), while the other 5 strains were resistant to 5 antibiotics (ampicillin, chloramphenicol, nalidixic acid, tetracycline, and trimethoprim-sulfamethoxazole). The highest resistance rate was observed to ampicillin (100%), nalidixic acid (100%), and tetracycline (100%).
AMP, ampicillin; C, chloramphenicol; CAZ, ceftazidime; CTX, cefotaxime; CN, amikacin; KF, cephalothin; KZ, cefazolin; NA, nalidixic acid; S, streptomycin; SXT, trimethoprim/sulfamethoxazole; TE, tetracycline.
The level of antibiotic resistance reported in this study was considerably higher than that reported by previous studies in South Korea. Thus, Cheong et al. (2007) observed that about 50% of isolates from broiler chickens were resistant to ampicillin, tetracycline, and nalidixic acid; another study showed that 90% of Salmonella isolates from retail chickens were resistant to 1 or 2 antibiotics (Hyeon et al., 2011). However, Kim et al. (2012) found a high occurrence of MDR Salmonella in chicken meats: among 47 strains isolated between January and May 2011, 41 isolates (87.2%) were resistant to at least 3 antibiotics. In South Korea, tetracyclines, cephems, quinolones, and aminoglycosides are the most frequently used antibiotics in the broiler industry (Chung et al., 2003; Cheong et al., 2007; Kim et al., 2012). Although the Korean government has banned the use of antibiotic feed additives as chicken growth promoters since July 2011 (Kim et al., 2012), antibiotic resistance does not seem to decrease dramatically.
ESBL-producing Salmonella
The six strains from slaughterhouse E showed resistance to tested cephalosporin antibiotics such as cephalothin, cefazolin, cefotaxime, and ceftazidime. All these strains were positive for ESBL production in the disc test. The resistance of ESBL-positive strains to cephalosporins was further confirmed using the Vitek n224 kit. The susceptibility of the ESBL-positive isolates to broad-spectrum cephalosporins is presented in Table 3. All isolates were resistant to cefotaxime, ceftazidime, cefepime, and aztreonam, as indicated by the high MICs (Table 3). However, these strains were susceptible to carbapenems, including ertapenem (MIC ≤0.5 mg/L) and imipenem (MIC ≤0.25 mg/L). These six ESBL-producing Salmonella strains harbored bla CTX-M genes (Table 3) of the CTX-M-1 cluster; the genes encoding TEM and SHV enzymes were not detected. All of the bla CTX-M genes were identified by sequencing as bla CTX-M-15 (Table 3), which is one of the most prevalent ESBL gene worldwide (Ewers et al., 2012).
All isolates had positive Vitek2 test results. AMP, ampicillin; CTX, cefotaxime; KZ, cefazolin; CAZ, ceftazidime; FEP, cefepime; AZT, aztreonam; FOX, cefoxitin; AMC, amoxicillin/clavulanic acid; TZP, piperacillin/tazobactam; SHV, sulphydryl variable; TEM, Temoniera. All strains were susceptible to imipenem (minimum inhibitory concentration [MIC] ≤0.25 μg/mL) and ertapenem (MIC ≤0.5 μg/mL).
The occurrence of ESBL-producing Salmonella shown in this study (6 of 11 samples, 54.5%) is higher than that reported by any other studies (0–17%) conducted in such countries as the United States (0% in retail meats; Zhao et al., 2009), Portugal (0.45% in retail meats; Clemente et al., 2013), China (8.6% in chicken meats; Wu et al., 2013), and Kuwait and United Arab Emirates (17% in clinical isolates; Rotimi et al., 2008). In a previous study conducted in South Korea, Tamang et al. (2011) reported that 5.8% of Salmonella isolates (13 of 225) from poultry samples collected from 1995 to 2009 were ESBL positive; all of them expressed CTX-M-15 but not TEM or SHV enzymes, which is consistent with our findings. Rayamajhi et al. (2010) found that 16.5% of Salmonella isolates (15 of 91) from chicken waste collected in the farms between 2006 and 2007 harbored the bla TEM-1 or bla DHA-1 genes. However, the information generated from our study is limited to compare with other previously conducted nationwide studies. The number of tested Salmonella strains was lower than that in previous studies. Furthermore, potential duplicate strains may be identified with our sampling method.
Salmonella infections caused by CTX-M-15-producing strains have been previously reported in South Korea. Salmonella enterica serotype Enteritidis isolated from the stool of a 3-year-old boy with gastroenteritis was found to produce the CTX-M-15 enzyme (Lee et al., 2009). Kim et al. (2011) investigated 49 clinical Salmonella strains isolated from 15 hospitals located in 11 cities, and found 6 ESBL-positive strains of the CTX-M-15 type (12.2%). The present study did not investigate the relationship between human and poultry isolates; however, Cheong et al. (2007) reported a clonal relationship between Salmonella strains isolated from human samples and broiler chickens in South Korea, suggesting a possibility of Salmonella transmission from livestock to humans.
As mentioned above, cephalosporins and fluoroquinolones are the drugs of choice for invasive Salmonella infections. ESBL-positive strains also carry the genetic determinants for the resistance to aminoglycosides, fluoroquinolones, chloramphenicol, and tetracycline (Shahada et al., 2006; Rodriguez et al., 2009; Wu et al., 2013). In this study, six ESBL-producing strains were found resistant to aminoglycosides and tetracycline as well as cephalosporins. None of the strains was resistant to ciprofloxacin and norfloxacin. However, all ESBL-producing strains were highly resistant to nalidixic acid (Table 2), and one strain, KI_11, exhibited reduced susceptibility to ciprofloxacin (MIC 0.5 μg/mL, data not shown) in Vitek 2 assay. Previous studied reported that nalidixic acid resistant Salmonella spp. with reduced susceptibility to fluoroquinolone (MIC 0.25–1 μg/mL) may lead to treatment failure (Valdezate et al., 2007; Lee et al., 2011).
Molecular subtyping of isolates
To evaluate the genetic similarity of 11 antibiotic-resistant Salmonella isolates, their chromosomal DNA was characterized using automated rep-PCR. The dendrogram and computer-generated virtual gel images generated by the DiversiLab are presented in Figure 1. We classified the strains into five pulsotypes, A–E, based on the rep-PCR banding patterns with ≥95% similarity. The banding patterns of 4 Salmonella Enteritidis isolates (KI_11, 13, 14, and 15) from slaughterhouse E included in Class C indicated that they were genetically identical or closely related (Fig. 1); however, the other two Salmonella Enteritidis isolates (KI_3 and 4) showed lower similarity with the rest of the group and were placed in Class B and D. Five Salmonella Bellevue isolates were classified into 3 pulsotypes with ≥95% similarity (Fig. 1).
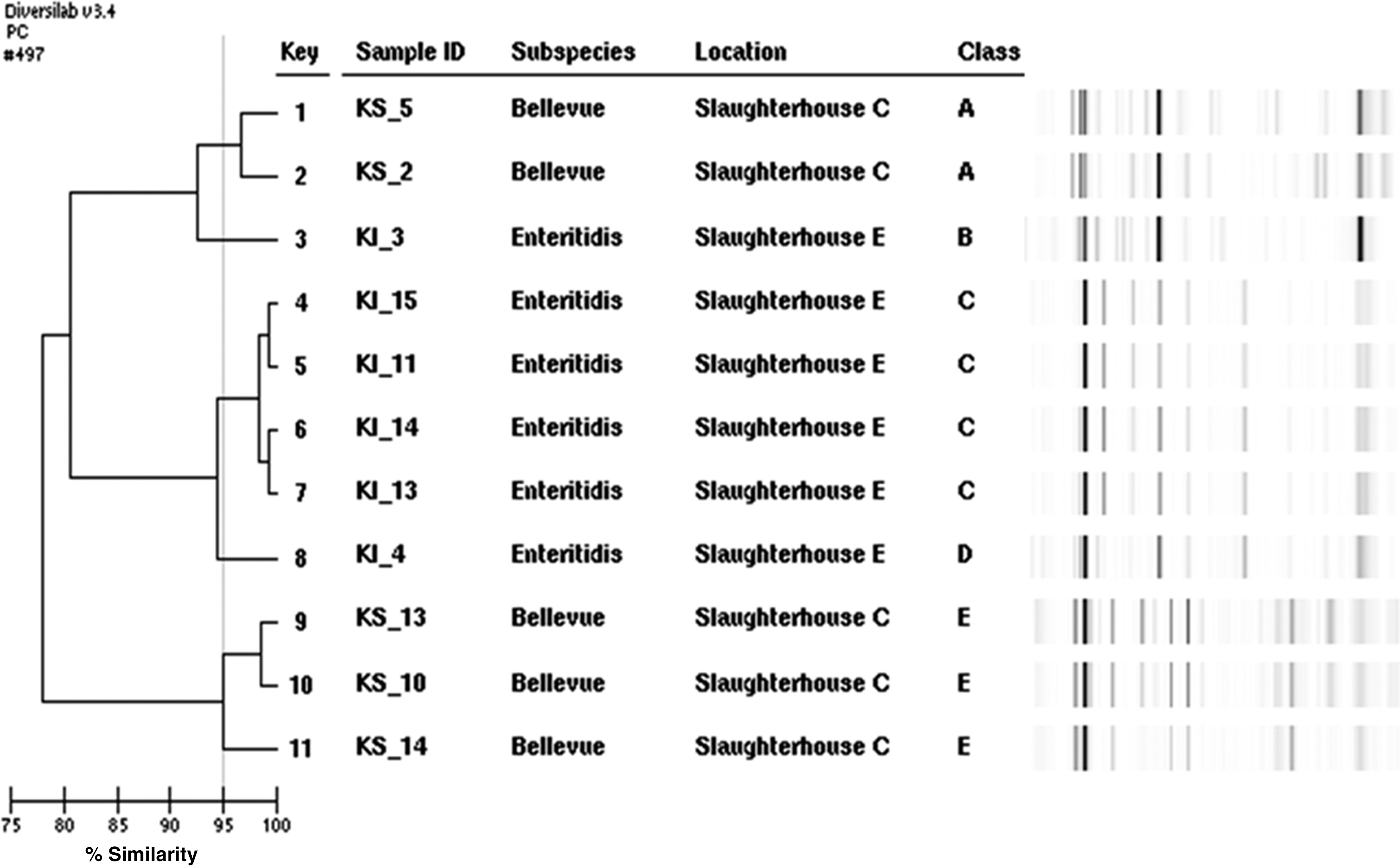
Dendrogram analysis and virtual gel image of DiversiLab™ rep-polymerase chain reaction (PCR) fingerprinting analysis of 11 Salmonella isolates from slaughterhouses C and E. The isolates showing ≥95 similarity in their rep-PCR banding patterns were classified into 5 pulsotypes (A–E).
At first, we assumed that six Salmonella Enteritidis isolates were genetically the same stain, because they had the same serotype and antibiotic-resistance pattern and were obtained in the same poultry slaughterhouse at same day. Although most Salmonella strains did not exhibit the identical molecular subtyping patterns (Fig. 1), these similar or identical isolates may be originated either from a farm that exclusively supplied live broilers to a specific poultry slaughterhouses or from environment of a processing plant.
Currently, pulsed-field gel electrophoresis (PFGE) is considered the “gold standard” method for molecular subtyping of Salmonella spp. (Ridley et al., 1998; Rivoal et al., 2009). However, in our previous study, we have found that rep-PCR provided better discriminatory power for phenotypically similar Salmonella Enteritidis isolated from human clinical samples and food products than did PFGE and multilocus sequencing typing (Hyeon et al., 2013). In this study, by using rep-PCR we were able to distinguish between Bellevue and Enteritidis serovars and to classify the isolates into 5 different pulsotypes based on ≥95 similarity in rep-PCR banding patterns.
Conclusions
In this study, we investigated Salmonella contamination in chicken carcasses from six poultry slaughterhouses in South Korea and found a high incidence of antibiotic-resistant strains. These strains exhibited resistance to at least 6 antibiotics, and 54.5% of them produced the ESBL enzyme. In another experiment currently conducted in our research group, however, a similar pattern of ESBL-producing Salmonella occurrence was observed in retail chickens from several chicken meat brands (unpublished report). The emergence of antibiotic-resistant Salmonella, including ESBL-producing strains, in food animals poses a potential risk to public health and should be regularly monitored to developing proper control measures.
Footnotes
Acknowledgments
This research was supported by Golden Seed Project, Ministry of Agriculture, Food and Rural Affairs (MAFRA), Ministry of Oceans and Fisheries (MOF), Rural Development Administration (RDA), and Korea Forest Service (KFS).
Disclosure Statement
No competing financial interests exist.