
Abstract
Zinc (Zn) is often supplemented at elevated concentrations in swine diets, particularly in piglets, to prevent enteric infections and promote growth. Previous studies from Denmark have suggested a genetic linkage and a phenotypic association between Zn resistance, encoded by czrC, and methicillin-resistance conferred by mecA in Staphylococcus aureus. Such an association has not been reported in the U.S. swine population. We conducted an analysis of the effects of Zn, supplemented as zinc oxide (ZnO), on the nasal carriage of methicillin-resistant Staphylococcus aureus (MRSA) in nursery (n=40) and finisher pigs (n=40) enrolled in a nutritional study. Nasal swabs, collected from nursery and finisher pigs, were inoculated onto MRSA CHROMagar and presumptive MRSA colonies were tested for the presence of mecA and czrC genes by polymerase chain reaction. Zinc susceptibility was determined by the agar dilution method. The prevalence of mecA-positive MRSA was 10% (4/40) and 20% (8/40) among nursery and finisher pigs, respectively. Of the 12 mecA-positive S. aureus isolates, 7 had the czrC gene (58.3%) compared to none among the 68 mecA-negative isolates. The presence of both mecA (p=0.002) and czrC (p=0.006) genes were positively associated with higher levels of Zn supplementation. The median minimum inhibitory concentrations of Zn for czrC-positive and czrC-negative isolates were 12 and 2 mM, respectively (p<0.0001). The link between czrC and mecA genes suggests the importance of elevated Zn supplementation in the co-selection and propagation of methicillin resistance among S. aureus in pigs.
Introduction
M
Materials and Methods
Animals, diet, and sampling
The use of animals and the procedures employed were approved by the Kansas State University Animal Care and Use Committee. The study was conducted with nursery (n=40) and finisher (n=40) pigs. The nursery pigs were divided into a control (n=20) and zinc (ZnO)-supplemented (n=20) groups. The control group received a basal diet consisting of zinc, included as zinc sulfate (ZnSO4), at 55 mg/kg of feed, as part of the normal trace mineral mix. In the zinc-supplemented group, nursery pigs received the basal diet (including ZnSO4 at 55 mg/kg of feed) supplemented with 1800 mg/kg of zinc, as zinc oxide (ZnO; Zinc Nacional S. A., Monterrey, Mexico). The study with finisher pigs consisted of control (n=10; no supplemental ZnO), low zinc (n=10; ZnO at 75 mg/kg of feed), moderate zinc (n=10; ZnO at 150 mg/kg of feed), and high zinc (n=10; ZnO at 225 mg/kg of feed) groups. All 4 groups received a basal diet with zinc, as ZnSO4, at 110 mg/kg of feed. A nasal swab (CultureSwab™, Becton Dickson and Company, Franklin Lakes, NJ) was collected from each pig, placed in Stuart's transport medium, and transported on ice to the laboratory.
Isolation and identification of MRSA
Nasal swabs were enriched at 37°C for 24 h in Mueller-Hinton broth (Becton and Dickson, Sparks, MD) with 6.5% sodium chloride (Sigma-Aldrich, St. Louis, MO) (Moodley et al., 2011). The enriched broth was inoculated onto CHROM™ MRSA agar plates (Hardy Diagnostics, Santa Maria, CA) and incubated at 37°C for 24–36 h for MRSA isolation. Presumptive MRSA colonies (mauve to dark pink or magenta) from CHROM agar plates were selected for species confirmation, genotypic characterization for methicillin resistance, and phenotypic and genotypic characterizations for zinc resistance. The genus (staph for genus-specific 16S rRNA) and species (nuc) confirmation were done by PCR as per the procedure described by Zhang et al. (2004). The confirmed isolates were stored on Protect beads (Cryo-Vac®, Round Rock, TX) at −80°C. S. aureus strain 9B (Dr. Henrik Hasman, National Food Institute, Technical University of Denmark) served as positive control for staph and nuc genes. The amplified PCR products were 756 bp and 279 bp for staph and nuc genes, respectively.
PCR detection of mecA, czrC, and tet(M) gene
DNA was extracted by boiling a suspension of a couple of colonies in sterile nuclease-free water for 10 min. The suspension was centrifuged at 20,000 ×g for 1 min and the supernatant was treated with lysostaphin (Sigma-Aldrich) for 1 h at 37°C. DNA of the isolates were tested for methicillin and zinc resistance determinants, mecA and czrC genes, respectively. The primers and PCR conditions for the identification of mecA [forward: 5'-GTA GAA ATG ACT GAA CGT CCG ATA A-3', and reverse: 5'-CCA ATT CCA CAT TGT TTC GGT CTAA-3'] and czrC [forward: 5'-TAGCCACGATCATAGTCATG-3', and reverse: 5'-ATCCTTGTTTTCCTTAGTGACTT-3'] genes were as per Zhang et al. (2004) and Cavaco et al. (2010), respectively. All the primers were procured from Integrated DNA technologies (IDT, Coralville, IA) and GoTaq PCR master mix (Promega, Madison, WI) was used for all PCR reactions. S. aureus strains 34-1 (provided by Dr. Peter Davies, University of Minnesota) and 9B served as positive controls for mecA and czrC genes, respectively. The PCR products were run using 1.5% agarose gel with ethidium bromide in 1X Tris-Borate-EDTA buffer for 1 h at 120 V. One kilobyte plus DNA ladder (Life Technologies, Grand Island, NY) was used as a molecular marker. The amplified PCR products were 310 bp (Fig. 1A) and 655 bp (Fig. 1B) for mecA and czrC genes, respectively. The tet(M) gene was detected by PCR as per Trzcinski et al. (2000), and an Escherichia coli strain harboring plasmid pFD 310 served as a positive control. The DNA was extracted by the boil-prep method and the expected size of the tet(M) amplicon was 156 bp.
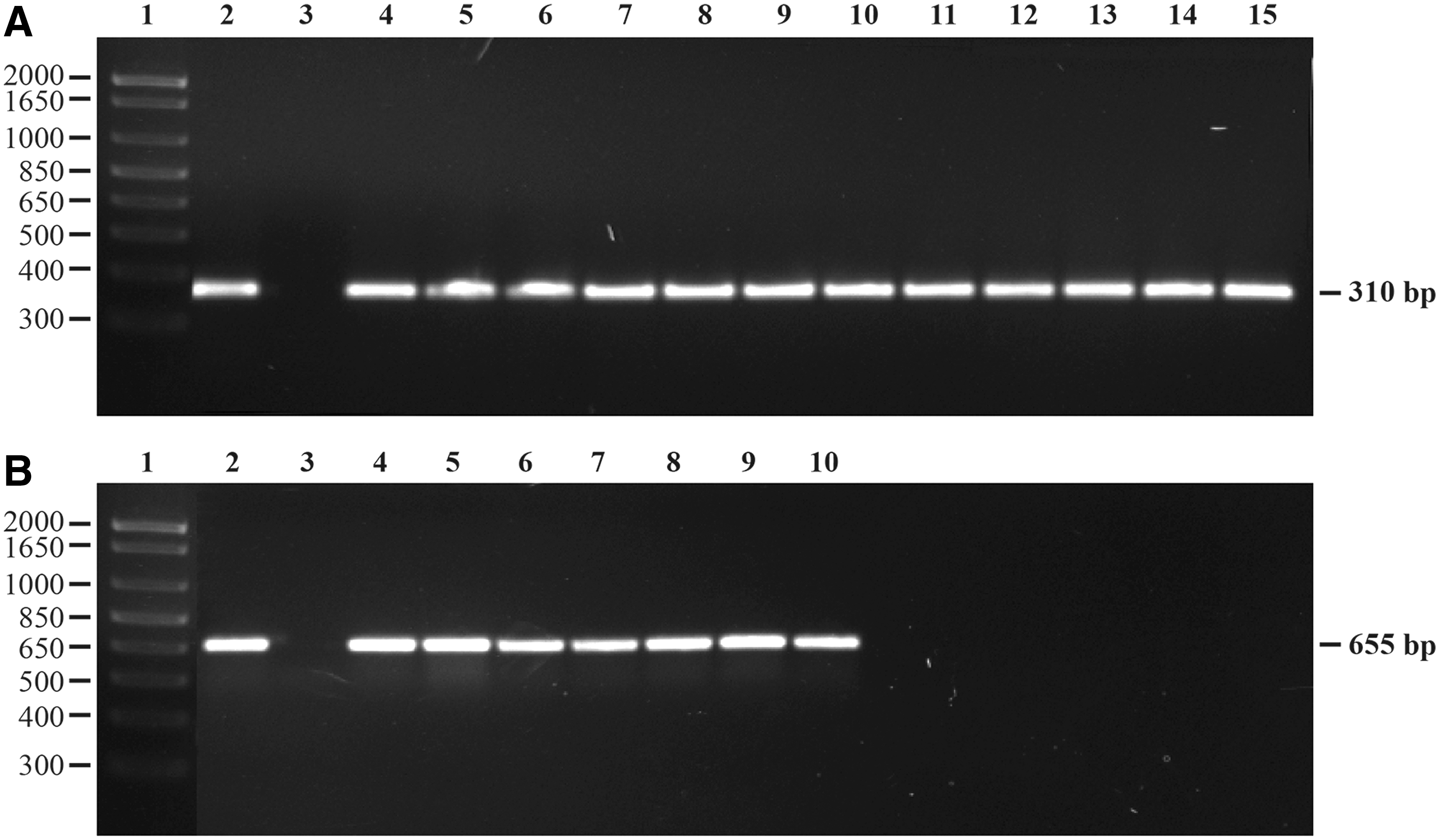
Polymerase chain reaction amplification of the mecA and czrC genes of Staphylococcus aureus isolates. (A) Lane 1: 1 kb plus DNA marker; Lane 2: Positive control for mecA gene (S. aureus strain 34-1); Lane 3: Negative control; and Lanes 4–15: mecA-positive isolates from this study. (B) Lane 1: 1 kb plus DNA marker; Lane 2: Positive control for czrC gene (S. aureus strain 9B); Lane 3: Negative control; and Lanes 4–10: czrC-positive isolates from this study.
Zinc and tetracycline susceptibility determinations
Zinc susceptibilities of S. aureus isolates were determined by the agar-dilution method (Aarestrup and Hasman, 2004). Mueller-Hinton agar plates containing 0, 1, 2, 4, 8, 12, and 16 mM of zinc chloride (Sigma-Aldrich) were used, with pH adjusted to 5.5 using 50% HCl. Bacterial inocula were prepared by growing the isolates in Mueller-Hinton II broth for 5–6 h and turbidity adjusted to a McFarland turbidity standard of 0.5. The plates, in duplicates, were spot inoculated with 15–20 μL of bacterial growth and incubated for 48 h at 37°C to determine growth or no growth. The minimum inhibitory concentration (MIC) of tetracycline was determined by microbroth dilution (adapted from CLSI [CLSI, 2008] with variations described below). Tetracycline susceptibilities were tested at concentrations of 100, 50, 25, 12.5, 6.25, 3.125, 1.56, 0.78, 0.39, 0.195, and 0.098 μg/mL by broth microdilution as per Amachawadi et al. (2011). Bacterial cultures were grown in 10 mL of Mueller-Hinton II broth for 6 h, and the inocula concentrations were adjusted to 0.5 McFarland turbidity standards (Remel, Lenexa, KS). The susceptibility determinations for antibiotics were carried out using 96-well microtiter plates (Becton and Dickson, Spark, MD) by incubating plates with the bacterial inocula at 37°C for 24 h and results were recorded as growth or no growth. Both zinc and tetracycline susceptibility determinations were repeated with different bacterial inocula preparations.
Statistical analysis
Data analysis was carried out using STATA SE version 12.1 (Stata Corp., College Station, TX). A descriptive analysis was performed prior to multivariable analysis. The analyses for binary endpoints were carried out via the multivariable exact logistic regression procedure, considering categorical dose of zinc (referent=basal diet) and controlling for the fixed effect of production age group. Zinc MIC data were analyzed using nonparametric survival analyses. Results were considered significant at p<0.05.
Results and Discussion
A nutritional study, primarily designed to evaluate the effects of feeding various concentrations of zinc on growth performance of nursery and finisher pigs, and reported on elsewhere (Cromwell, 2001), was utilized to determine the effects of zinc on the prevalence and association of mecA-positive and czrC-positive MRSA. In the nursery pigs, the prevalence of mecA-positive MRSA was 0% (0/20) in the control group and 20% (4/20) in piglets fed a diet supplemented with 1800 mg/kg of feed of ZnO (Fisher's one-sided exact test with 1 df: p=0.05; Fisher's two-sided test with 1 df: p=0.10). In the finisher pigs, the prevalence of mecA-positive MRSA was 0% (0/10), 10% (1/10), 20% (2/10), and 50% (5/10) in the groups that received 0, 75, 150, and 225 mg/kg of feed of supplemental zinc, respectively (Fisher's 2-sided exact test with 3 df: p=0.048). Zinc is supplemented as ZnO to nursery pigs at pharmacological levels (1800–3000 ppm) primarily to prevent enteric infections. However, in finisher pigs, zinc dose (50–250 ppm) varies, often supplemented as ZnO in phases, and primarily intended to promote growth (NRC, 1998). Of the total 12 mecA-positive S. aureus isolates from nursery and finisher pigs, 7 had the czrC gene (58.3%) compared to none among the 68 mecA-negative isolates (p<0.0001). In finisher pigs, the presence of czrC gene was also associated (Fisher's 2-sided test with 3 df: p=0.04) with increasing levels of zinc supplemented as ZnO in the diet (Table 1). The median MICs of zinc for czrC-positive and czrC-negative isolates were 12 and 2 mM, respectively (p<0.0001; Fig. 2). Our study is the first report of an association between MRSA prevalence and zinc supplementation in swine in the United States.

Failure function graph depicting cumulative susceptibilities of czrC-positive and czrC-negative isolates to zinc. MIC, minimum inhibitory concentration.
However, the observation was based on one-time sampling in a small number of pigs, and without knowing the source and feeding history of the pigs. Our results on the phenotypic and genotypic associations between MRSA and zinc provide further evidence to corroborate the findings from Denmark that zinc may play a role in the co-selection of MRSA. The zinc resistance determinant, czrC, has been reported to reside within the typeV SCCmec element (Li et al., 2011). Aarestrup et al. (2010) examined 32 MRSA and 60 MSSA isolated from different swine farms in Denmark for their susceptibility to antimicrobial compounds including zinc chloride. Twenty-three (74%) of the MRSA had higher MIC to ZnCl2 (4–12 mM) compared to MSSA (0.5–2 mM). The study reported by Cavaco et al. (2011) was on the occurrence of zinc resistance in MRSA of swine and veal calves origin from a global collection of isolates (n=476) from Europe, Canada, and China. Zinc resistance, based on MIC (>2 mM) and presence of czrC, was observed in 80.7% (363/451) of European strains, 44% (8/18) of Canadian strains, and none (0/8) of Chinese strains. Based on the widespread occurrence of zinc resistance in MRSA, Cavaco et al. (2011) suggested that the use of zinc may have contributed to the emergence of MRSA. Another study from Denmark (Moodley et al., 2011) indicated that pigs fed diets supplemented with tetracycline or zinc had higher counts of MRSA in the nasal cavity. In our study, the prevalence of the tet(M) gene in S. aureus isolates was 28.7% (23/80). The association of tet(M) with mecA [OR=0.85 (95% CI: 0.29–2.42), p=0.75] or czrC [OR=0.48 (95% CI: 0.07–3.00), p=0.37] gene was nonsignificant. The median MICs of tetracycline for tet(M)-positive and tet(M)-negative isolates were 100 μg/mL and 12.5 μg/mL, respectively (p<0.0001).
It has been previously reported that all MRSA strains from swine are tetracycline resistant (de Neeling et al., 2007), suggesting tetracycline use may be one of the reasons for the emergence of MRSA. However, the results from our study showed that there was no difference in tet(M)-prevalence and in MIC between MRSA and MSSA isolates.
Metals, such as copper and zinc, are extensively used as alternatives to antibiotics for promoting growth and feed efficiency in pigs. It is reported that supplementation with heavy metals selects for metal resistance and can also co-select for resistance to other antimicrobial agents, such as antibiotics (Hasman and Aarestrup, 2002; Amachawadi et al., 2011). Associations between the acquired copper resistance gene (tcrB) and macrolide and vancomycin (Hasman and Aarestrup, 2002), or macrolide and tetracycline resistance (Amachawadi et al., 2011) have been shown in fecal enterococci of piglets. The emergence and propagation of livestock-associated MRSA in food animal agriculture are of major concern for public health (Smith et al., 2013). From our study, it is evident that zinc, if fed at an elevated level in both nursery and finisher pigs, is shown to increase the prevalence of czrC- and mecA-positive S. aureus. The genetic link between czrC and mecA genes suggests the importance of elevated zinc supplementation in co-selection and propagation of antibiotic resistance. From our study, it is likely that the basal diet alone (as ZnSO4) in both nursery (55 mg/kg of feed) and finisher (110 mg/kg of feed) pigs did not select for czrC-positive MRSA; however, since a Zn-negative control group was not included, we cannot rule this out completely. Importantly, zinc is a required micronutrient in swine diets and so a truly negative zinc diet is impractical at best, and potentially dangerous to swine health and well-being at worst. A bias toward the null hypothesis of no effect is the most likely statistical impact of the ZnSO4 included in the diet at basal levels; despite this, we detected a significant association with the ZnO levels added beyond the basal levels provided by the ZnSO4. Further studies are needed to better understand the effects of feeding zinc and other antimicrobials alone, or in combination, on the prevalence of MRSA in pigs.
Footnotes
Acknowledgments
The authors would like to thank Dr. Peter Davies and Dr. Henrik Hasman, for providing the reference strains for our experiments, and Mal Hoover for help with the images.
Disclosure Statement
No competing financial interests exist.