
Abstract
Periplasmic proteins are particularly vulnerable to denaturation upon entry into a highly acid environment. In Escherichia coli, a level of protection of these proteins is afforded by acid-inducible chaperonins encoded by hdeAB. In contrast, Salmonella enterica only harbors an hdeB-like gene and it is currently not known what function it plays in this genus. In the present study, the hdeB-like gene was deleted in Salmonella enterica Enteritidis NalR and Salmonella enterica Kentucky 3795. When grown overnight in tryptic soy broth (TSB) medium buffered at pH 5.5 and then exposed to TSB pH 2 for 20 min, Enteritidis wild-type strain experienced a 0.5-log10 reduction in colony-forming units, whereas the deletion strain's surviving cells were reduced by 1.6 log10. No difference in survival was observed in the corresponding Salmonella enterica Kentucky 3795 strains treated the same way. Exposure of the strains to pH 2.5 or 3 resulted in the same log reduction regardless of the presence of the hdeB-like gene. When wild-type and deletion strains of both serovars were grown in medium buffered at pH 7 prior to exposure to the acidic pHs, no difference in survival with respect to serovar or presence/absence of the hdeB-like gene was found. Salmonella enterica Enteritidis NalR carrying its own or the intragenic region upstream of the hdeB-like from Salmonella enterica Kentucky 3795 cloned in front of the gfp gene from pFPV25 showed maximum fluorescence when grown at pH 5.5, whereas the corresponding plasmid-carrying Salmonella enterica Kentucky strains did not exhibit fluorescence regardless of the pH of the growth medium. Therefore, the hdeB-like gene in Salmonella enterica Enteritidis, but not in Salmonella enterica Kentucky 3795, contributed to survival at pH 2 and its expression is responsive to the pH of the medium.
Introduction
H
In vivo, optimal resistance of E. coli K-12-type strains to pH 2 and pH 3 requires the presence of both hdeA and hdeB (Kern et al., 2007). It was therefore surprising that not all strains of E. coli express both hdeA and hdeB. In E. coli O157:H7 isolates, only HdeA is produced as the strains studied carry a mutation that changes the hdeB start codon from ATG to ATA (Carter et al., 2012). Comparison of survival of wild-type E. coli K-12 and E. coli O157:H7 and their corresponding hdeA or hdeB deletion strains upon exposure to LB medium pH 2 revealed a complex picture of the effects of HdeA and HdeB on survival at pH 2 (Carter et al., 2012). Prior growth temperature and salt content of the medium influenced survival of the strains. The most pronounced decrease in survival was observed with the K-12 hdeB deletion strain grown in salt-free LB medium at 28°C prior to exposure to LB medium pH 2. This observation suggests that despite the previously observed lower efficiency of HdeB at pH 2 in vitro, situations may exist in vivo that require the function of HdeB at pH 2.
In a survey for the presence of hdeAB-like genes in other bacteria, Wang et al. (2012) identified the genera Shigella, Enterobacter, and Providencia as also harboring hdeAB-like genes. Achromobacter, Laribacter, Brucella, Bordatella, Thiocapsa, Methylocystis, and Methylobacter genomes only contained an hdeA-like gene, whereas Yersinia, Edwardsiella, Klebsiella, Citrobacter, and Salmonella only harbored an hdeB-like gene. The role of these hdeA- or hdeB-like genes in these organisms has not been studied, and it is not known why some bacteria carry both genes and some only one of them. Transposon insertions in the hdeB-like gene of Salmonella enterica Typhimurium created mutants that survived similarly to the wild type in mice injected with these bacteria (Chaudhuri et al., 2009, 2013); however, some level of attenuation was observed in orally challenged chickens, cattle, and pigs (Chaudhuri et al., 2013). This observation suggests that hdeB from Salmonella enterica Typhimurium might be important for survival during passage through the acidic stomach environment.
Microarray and quantitative polymerase chain reaction data obtained with serovars Enteritidis and Kentucky of Salmonella enterica suggest that the expression of the hdeB-like genes might not be the same in all Salmonella (Joerger et al., 2012), and it was speculated that the differences in the acid-tolerance response by the two serovars could be due, at least to some extent, to differences in the expression of their hdeB-like genes. The Enteritidis strain showed increased hdeB-like mRNA levels when exposed to pH 5.5 for 10 min, whereas the level of hdeB-like transcripts remained low in the Kentucky strain under the same conditions.
In the present study, the hdeB-like genes from Salmonella enterica Enteritidis NalR (SEN1493) and Salmonella enterica Kentucky 3795 (SeKA_1012) were deleted and the acid-response phenotypes of the wild-type and mutant strains were compared. In addition, the genomic regions between the gene preceding the hdeB-like genes, osmC, and the start codon of the hdeB-like genes were inserted into vector pFPV25 in front of the promoterless gfp gene to analyze the activities of the promoters of the hdeB-like gene in cells grown at different pH.
Materials and Methods
Strains, plasmids, and media
Salmonella enterica Enteritidis NalR and Salmonella enterica Kentucky 3795 (Joerger et al., 2009) were maintained on tryptic soy agar (TSA) and grown in tryptic soy broth (TSB) for most experiments. Since the pH of cultures in TSB increases slightly with the age of the culture, cultures with stable pH were grown in TSB supplemented with 50 mM 2-(N-morpholino)ethanesulfonic acid (MES) adjusted to pH 7 or pH 5.5. Cultures were routinely grown at 37°C, but strains were also grown at 28 and 42°C as required by the mutagenesis protocol (Datsenko and Wanner, 2000). When required, kanamycin or ampicillin was added to TSA or TSB at a concentration of 50 μg/mL. Plasmid isolations from E. coli or Salmonella strains were accomplished using plasmid isolation kits (Origen, Rockville, MD). For lambda red-based mutagenesis, plasmid-carrying E. coli strains were acquired from the E. coli Genetic Stock Center (New Haven, CT), whereas plasmid pFPV25 (Valdivia and Falkow, 1996) was a gift from R. Valdivia. Primers were obtained from Eurofins MWG Operon (Huntsville, AL) and are listed in Table 1.
PCR, polymerase chain reaction.
Deletion mutagenesis
The Salmonella enterica Enteritidis NalR SEN1493 and the Salmonella enterica Kentucky 3795 SeKA_1012 genes were replaced with kanamycin-resistance cartridges by lambda red-mediated gene replacement (Datsenko and Wanner, 2000) and the cartridge was removed according to Cherepanov and Wackernagel (1995) using the strains, plasmids, and primers as listed in Table 1. Plasmid and linear DNA was introduced into the Salmonella enterica strains using electroporation with a BTX® ECM630 instrument (Harvard Apparatus, Hollister, MA) at 1.8 kV, 200 Ω, 25 μF. The Salmonella strains were made electrocompetent by growing cells to OD600∼0.6 in 20 mL TSB. The cells were washed 3 times in 10% ice-cold glycerol and finally suspended in 200 μL of 10% glycerol. Successful deletion of the hdeB-like genes from the two Salmonella isolates was verified by sequencing using the primers listed in Table 1.
Cloning of promoter regions
The regions upstream of Salmonella enterica Enteritidis NalR SEN1493 and the Salmonella enterica Kentucky 3795 SeKA_1012 were polymerase chain reaction (PCR)–amplified using the primers listed in Table 1. The resulting fragments were ligated to BamHI-EcoRI-digested pFPV25 (Valdivia and Falkow, 1996), resulting in plasmids that carried the inserts in both orientations with respect to the gfp gene. The plasmids were introduced into the two Salmonella enterica strains by electroporation and selection for ampicillin resistance. The presence of the inserts was verified by PCR amplification.
Acid survival tests
The Salmonella enterica Enteritidis NalR SEN1493 and the Salmonella enterica Kentucky 3795 SeKA_1012 deletion strain were grown in TSB, 50 mM MES pH 7 or pH 5.5 overnight with shaking at 200 rpm. A pH of 5.5 was chosen to allow the development of the acid-tolerance response (Foster and Hall, 1991). Viable cell counts were established by dilution in TSB 100 mM TRIS pH 8. For acid exposure, the overnight cultures were diluted 100-fold into TSB adjusted to pH 2, 2.5, or 3 with HCl. Exposure times and pHs were chosen based on initial experiments. Exposure times (Fig. 1) were chosen so that the highest levels of inactivation could be achieved reliably without a reduction in the cell counts of any of the exposed strains below the limit of detection. The 3 pHs were chosen because at a pH below 2, inactivation was too rapid and above 3, too little inactivation was observed over a 12-h period. After exposure times as indicated in Figure 1, the cells were diluted into TSB 100 mM TRIS-HCl pH 8 and plated onto TSA. Colonies were counted after incubation at 37°C for 24 h. Differences in the observed reductions in colony-forming units were analyzed using the Student t-test function of JMP® 11.2.0 (SAS, Cary, NC).
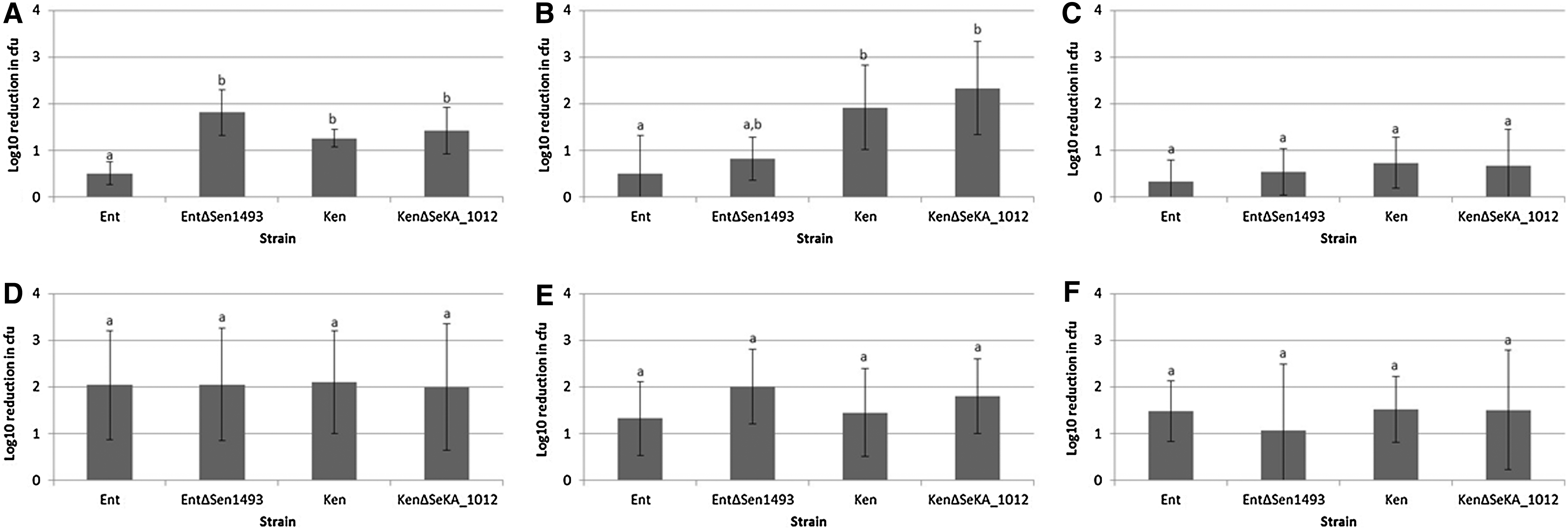
Log10 reductions in colony-forming units (cfu) of wild-type strains and hdeB-like gene deletion strains after exposure to tryptic soy broth (TSB) medium acidified with HCl. The values are the medians of results from nine trials and the error bars indicate the standard deviation.
gfp expression experiments
The plasmid-carrying strains were grown in 200-μL volumes in microtiter plates overnight in TSB 50 mM MES pH 7 or pH 5.5 containing ampicillin at a concentration of 50 μg/mL. The cultures were diluted 100-fold into TSB containing ampicillin (50 μg/mL) and 50 mM MES pH 4.5, 5, 5.5, or 6 or in TSB with 50 mM MOPS pH 6.5, 7, 7.5, or 8. The different buffers were chosen to maximize buffering capacity at the different pH ranges. Optical density was read by a BioTek Synergy 2 plate reader (BioTek Instruments, Inc., Winooski, VT) at 600 nm. Fluorescence was measured with an excitation wavelength of 485 nm and an emission wavelength of 528 nm. Plates were incubated at 37°C and readings were taken every hour. The fluorescence readings were divided by the corresponding OD600 readings and the values obtained with strains carrying pFPV25 without an insert were subtracted from the values measured with the strains carrying inserts in pFPF25. A total of six independent experiments were run.
Results and Discussion
Although the amino acid sequences of HdeA and HdeB from E. coli share only 19% sequence identity, their structure and biochemical properties are very similar (Wang et al., 2012). Based on Blast alignment analyses, the predicted HdeB-like proteins from Salmonella share 43% amino acid sequence identity with their E. coli counterparts and a 27% level of identity with that of HdeA from E. coli which is, interestingly, higher than the identity of HdeA and HdeB from E. coli (19%). The amino acid sequences of the HdeB-like proteins from Salmonella enterica are identical, and the Salmonella bongori counterparts only differ in 1 of 109 amino acids. This observation could suggest a conserved and useful role of the proteins in this genus, but differences in acid response gene expression profiles of Salmonella serovars (Joerger et al., 2012) indicate that, as in E. coli, expression of the hdeB-like genes in Salmonella might not be the same in all serovars.
The present study demonstrated that the loss of the hdeB-like gene from Salmonella enterica Enteritidis NalR caused a decrease in survival of cells previously exposed to mildly acid conditions (pH 5.5) when transferred to medium of pH 2 (Fig. 1A). Cells harboring an hdeB-like gene and previously grown in medium buffered at pH 7 did not exhibit a difference in survival when compared with the hdeB deletion strain (Fig. 1D). The cells were considerably more sensitive to pH 2 (exposure time of only 5 min rather than 20 min was required to obtain reliable cell counts). Upon exposure to an only marginally higher pH of 2.5 or 3, no protective effect of having an hdeB-like gene was observed for cells pre-exposed to pH 5.5 (Fig. 1B,C,E,F). Thus, under the conditions tested, the putative HdeB protein from Salmonella enterica Enteritidis NalR appears to be important at a pH lower than that at which HdeB from E. coli was reported to be most efficient in preventing protein aggregation in vivo (pH 3) (Kern et al., 2007).
Previous studies had shown that Salmonella enterica Kentucky strains were more acid sensitive than other Salmonella serovars examined (Joerger et al., 2009), and that expression of hdeB in Salmonella enterica Kentucky 3795 was lower than that in Salmonella enterica Enteritidis NalR under acidic conditions and unresponsive to exposure to pH 4.5 and 5.5 (Joerger et al., 2012). In line with these findings, the current experiments also showed that serovar Kentucky cells pre-exposed to pH 5.5 survived less than the Salmonella enterica Enteritidis NalR counterparts when exposed to pH 2, 2.5, and 3 regardless of the presence of an hdeB-like gene (Fig. 1A–C).
In order to better understand differences in regulation of the expression of the hdeB-like genes between Salmonella enterica Enteritidis NalR and Kentucky 3795, the intragenic region between osmC and hdeB was placed upstream of the gfp gene in pFPV25, and fluorescence was monitored upon growth of the cells at different pHs. Fluorescence above the level measured in strains with pFPV25 without an insert was only observed in Salmonella enterica Enteritidis NalR harboring plasmids with inserts of its own hdeB promoter region or the promoter region from Salmonella enterica Kentucky 3795 in the orientation found in the respective chromosomes (Fig. 2). Insertion of the intragenic region in the reverse orientation resulted in plasmids that did not confer fluorescence above the level conferred by the insert-free plasmid. These observations suggest that the intragenic region is transcriptionally active and that the previously observed inability to increase expression of the hdeB-like gene in Salmonella enterica Kentucky 3795 under acidic conditions (Joerger et al., 2012) was not due to a defect in its promoter region, but might be due to regulatory elements not being present in the Kentucky strain under the tested conditions. The appearance of fluorescence took several hours, perhaps because expression is connected to stationary phase and/or because of limited diffusion of oxygen into the medium in the microtiter plates. The highest level of fluorescence units per OD600 unit was observed around pH 5.5 after 8 h of culturing. Very little fluorescence was observed at or above neutral pH. The differences in the values seen in Figure 2 did not originate from differences in growth rates of the strains, as the rates were the same in all cases. These data indicated that in Salmonella enterica Enteritidis NalR, the hdeB-like gene might be responsive to growth stage and medium acidity as was observed for E. coli (Tucker et al., 2002; Weber et al., 2005). It remains to be determined whether the hdeB-like gene from Salmonella is subject to the same complex regulation as hdeAB from E. coli that includes Lrp, H-NS, GadEXW, MarA/SoxS, and TorR (Ruiz et al., 2008).
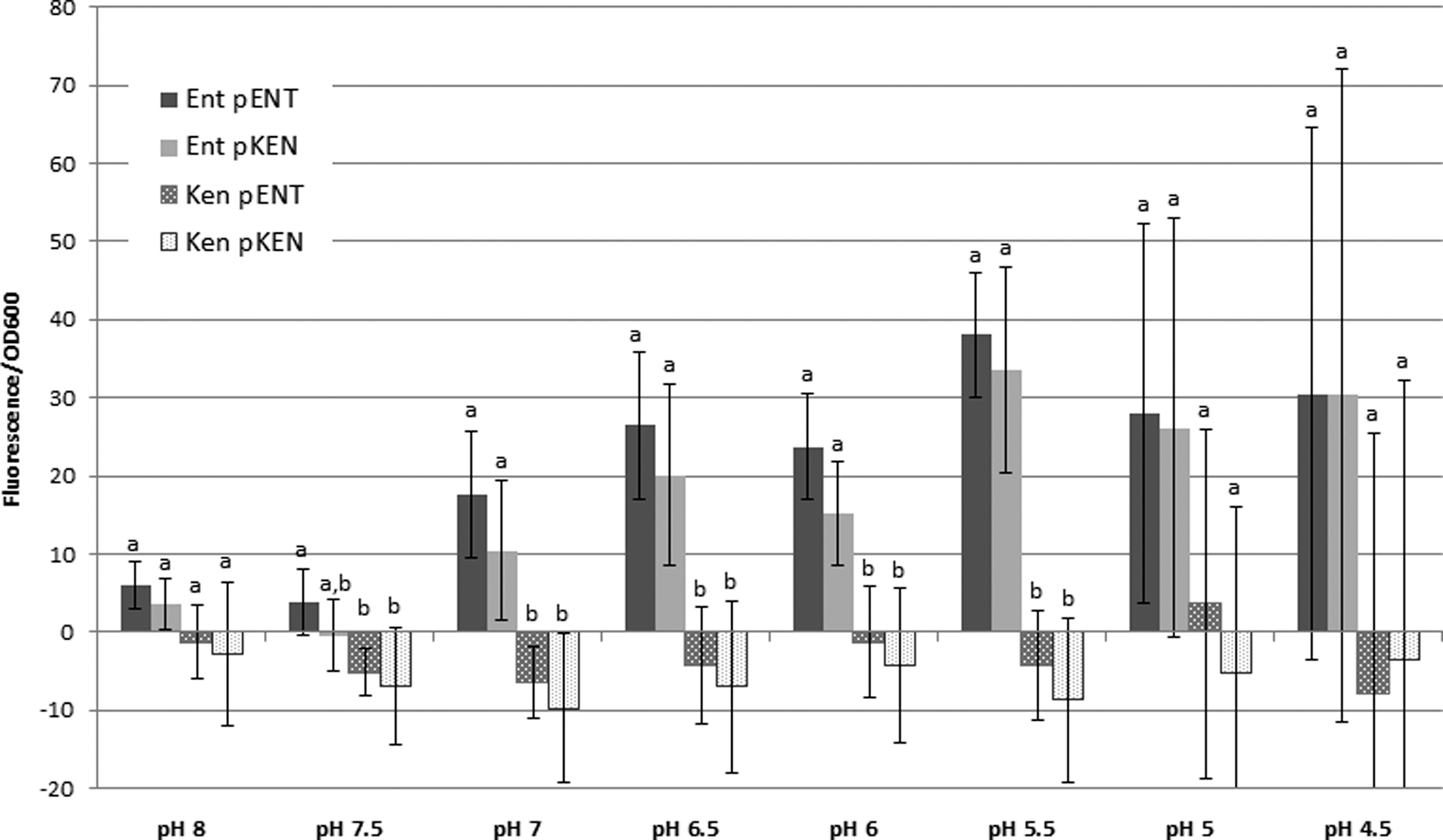
Difference in fluorescence/OD600 of strains carrying pFPV25 with insertions and the corresponding strains without the insertions in the plasmid when grown in tryptic soy broth media of different pHs for 8 h. The values are averages of results from six trials and error bars indicate standard deviation. Ent pENT, Salmonella enterica Enteritidis pFPV25 with the Salmonella enterica Enteritidis SEN1493 promoter region inserted upstream of the gfp gene of the plasmid; Ent pKen, Salmonella enterica Enteritidis NalR pFPV25 with the Salmonella enterica Kentucky 3795 SeKA_1012 promoter inserted upstream of the gfp gene of the plasmid. Ken pENT, Salmonella enterica Kentucky 3795 pFPV25 with the Salmonella enterica Enteritidis SEN1493 promoter region inserted upstream of the gfp gene; Ken pKEN, Salmonella enterica Kentucky 3795 pFPV25 with the Salmonella enterica Kentucky 3795 SeKA_1012 promoter region inserted upstream of the gfp gene. Different letters above the bars indicate that the results are different at a 95% CI level (Tukey HSD).
Footnotes
Disclosure Statement
No competing financial interests exist.