
Abstract
The use of essential oils as a food preservative has increased due to their capacity to inhibit vegetative growth of some bacteria. However, only limited data are available on their effect on bacterial spores. The aim of the present study was to evaluate the effect of some essential oils on the growth and germination of three Bacillus species and Geobacillus stearothermophilus. Essential oils were chemically analyzed using gas chromatography and gas chromatography coupled to mass spectrometry. The minimal inhibitory and bactericidal concentrations of vegetative growth and spore germination were assessed using the macrodilution method. Germination inhibitory effect of treated spores with essential oils was evaluated on solid medium, while kinetic growth was followed using spectrophotometry in the presence of essential oils. Essential oil from Drypetes gossweileri mainly composed of benzyl isothiocyanate (86.7%) was the most potent, with minimal inhibitory concentrations ranging from 0.0048 to 0.0097 mg/mL on vegetative cells and 0.001 to 0.002 mg/mL on spore germination. Furthermore, essential oil from D. gossweileri reduced 50% of spore germination after treatment at 1.25 mg/mL, and its combination with other oils improved both bacteriostatic and bactericidal activities with additive or synergistic effects. Concerning the other essential oils, the minimal inhibitory concentration ranged from 5 to 0.63 mg/mL on vegetative growth and from 0.75 to 0.09 mg/mL on the germination of spores. Spectrophotometric evaluation showed an inhibitory effect of essential oils on both germination and outgrowth. From these results, it is concluded that some of the essential oils tested might be a valuable tool for bacteriological control in food industries. Therefore, further research regarding their use as food preservatives should be carried out.
Introduction
G
To prevent spore contamination, industries usually use high temperatures to sterilize. However, this treatment generally leads to the degradation of nutritional and organoleptic qualities of food (Islam et al., 2006). Thus, an alternative strategy is to use chemical substances with antimicrobial activities, provided that they are nontoxic. This explains all the renewed interest directed toward plant essential oils. Currently, these substances are used in aromatherapy and as additives to add flavor and preserve various foods due to their organoleptic and antibacterial properties (Ndoye, 2001; Mohammedi, 2006; Huertas et al., 2014).
However, efficient use of essential oils as food preservatives requires a good knowledge about their properties (i.e., the chemical composition, the minimal inhibitory and the bactericidal concentrations [MIC and MBC], the panel of target microorganisms, and the mechanism(s) of action). To the present, many studies have been done on the chemical composition and inhibitory effect of essential oils on the growth of a wide range of bacteria, particularly on their vegetative forms, but few have been done on their effect on bacterial spores. Nevertheless, the study of Chaibi et al. (1997) assessed the effect of essential oils extracted from Moroccan plants on spores and vegetative cells of B. cereus. The results showed an inhibitory effect on one or more stages of spore germination with sporostatic or sporocidal effect. This study, which was focused on the mechanism(s) of action of essential oils on bacterial spores, might be extended to other essential oils and more Bacillus species. The purpose of this work was to evaluate the effect of nine essential oils extracted from Cameroonian aromatic plants on growth, germination, and outgrowth of three Bacillus species and G. stearothermophilus.
Materials and Methods
Plant material, extraction, and chemical analyses of essential oils
For essential oil extraction, nine aromatic plants were harvested or purchased and identified at the National Herbarium of Cameroon (Table 1). Essential oil extractions were done by hydrodistillation using a Clevenger apparatus, according to the protocol described by Moni (2013). Oils were analyzed by gas chromatography–flame ionization detector and GC coupled with mass spectrometry as described earlier by Nyegue (2006).
Bacterial strains, production of vegetative cells and spores
B. cereus T, B. subtilis NCTC 3610, and B. megaterium 8174 came from the collection of The Microbiology Laboratory Institute of Food Research of Reading, UK. G. stearothermophilus CNCH 5781 was obtained from the Institut Appert, Paris, France. These strains were provided in the form of spores. To obtain vegetative cells, spores were subjected to activation at 80°C for 10 min. They were then grown on nutrient agar for 24 h at the optimal growth temperature of each species (35°C for B. cereus, B. subtilis, and B. megaterium and 63°C for G. stearothermophilus). For spore production, cells grown for 12 h on trypticase soy broth were spread on sporulation agar medium as described by Bayoï et al. (2014). The plates were incubated at the aforementioned temperatures for 5 days. The spores were collected with a glass spatula, suspended in distilled water, and purified by many centrifugation cycles at 3000×g for 15 min. The purified spores were stored at 4°C for 3 months to ensure stability of spores before use (Voundi et al., 2014).
Macrodilution method
Using this method, minimal inhibitory concentration (MIC), minimal bactericidal (MBC) and minimal inhibitory concentrations for inhibition of spore germination (MICg) were assessed. The method on test tubes described by Gachkar et al. (2007) was used with the following modifications: for MIC determination, vegetative forms of Bacillus and G. stearothermophilus at 106 cells/mL were allowed to grow in 2 mL of nutrient broth (supplemented with 10 g/L of glucose, 0.2% Tween 80, and 0.05 mg/L of red phenol as growth indicator) at different concentrations of each essentials oil. The initial concentration was 20 mg/mL followed by a geometric progression of 1/2 until the 8th dilution for all the essential oils except for that of Drypetes gossweileri that went up to the 16th dilution.
For MICg determination, heat-activated spores (80°C, 10 min) at 106 spores/mL were inoculated in 2 mL of nutrient broth at a low concentration of essential oil to avoid its effect on the vegetative growth. The initial concentration was 1.5 mg/mL for all the essential oils except that of D. gossweileri (more active), which started at 0.004 mg/mL. A twofold dilution was made until the sixth dilution.
Tubes were incubated for 24 h at the optimal growth temperature of each species, and the MICg and MIC were respectively determined as the lowest concentration where visible growth or discoloration of red phenol was not observed.
MBCs were determined by inoculation of 100 μL of culture medium at greater than or equal to the MICs onto an agar plate and incubated for 24 h. The MBC was considered the lowest concentration of essential oils that allowed the survival of 0.01% of the initial inoculum (Oussou et al., 2008).
Effect of combinations of essential oils on the vegetative growth
According to their effectiveness, essential oils of Ocimum gratissimum, Zingiber officinale, and Eugenia caryophyllus were each combined with that of D. gossweileri and tested again on the vegetative growth of each species. Combinations were carried out by the checkerboard assay described by Burt et al. (2005) with slight modifications as follows: using the macrodilution method, 2.5 mg/mL of each of the first three essential oils were prepared in the first line and 0.019 mg/mL of D. gossweileri were prepared in the first column. The dilution was consistent with the description of Burt et al. (2005) on microplate. Each species at 106 cells/mL was added and then incubated for 24 h at the growth temperature. The MICs were determined as the smallest concentrations of combined essential oils having inhibited visible growth. The MBCs were determined by agar subcultures as described above.
The determined MICs were used to calculate the fraction inhibitory concentration (FIC) compared to individual MICs of essential oil with the formula shown below:
The FICs index were obtained as follows: FIC index (A/B)=FIC (A)+FIC (B).
Combination of essential oils was classified as synergistic (FIC index ≤0.5), additive (0.5 ≤FIC index ≤1.5), indifferent (FIC index=2), and antagonist (FIC index >2) (Bassolé and Juliani, 2012).
Treatment of spores with essential oil and evaluation of germination on solid medium
To perform this test, the method described earlier was used (Voundi et al., 2014). Fifty microliters of heat-activated spores (1.8×106 spores/mL) of each species were treated with each essential oil at the following concentrations: 20, 10, 5, 2.5, 1.25, and 0 mg/mL at 30°C for 20 min. Treatments were done in 0.2% Tween 80 for good dilution of essential oils, then 100 μL of appropriate decimal dilution of treated samples were spread on Muller–Hinton agar. The plates were incubated for 24 h at the optimal growth temperature of each species. The numbers of colony-forming units (CFU) were counted and expressed in terms of percentage of germination inhibition of spores using the formula below:
Kinetic of germination and outgrowth of spores with essential oils
To assess the kinetic of germination and outgrowth of spores with essential oils, heat-activated spores (1.8×106 spores/mL) were transferred to nutrient broth (pH 7) on test tubes, supplemented with 0.2% Tween 80 and essential oils of D. gossweileri (0.002 mg/mL), Cymbopogon citratus, Ageratum conyzoïdes, or Citrus limon (0.37 mg/mL). Spore germination and outgrowth kinetics were assessed by measuring the optical density using a Cary 100 UV-Vis–NIR spectrophotometer (Agilent Technologies, Les Ulis, France) at 620 nm (maximum absorption of the spores), every 10 min for 40 min at first and after every 1 h for 6 h after shaking (Maldonado et al., 2013). Control spores without essential oil were made under the same conditions. Optical densities recorded were used to draw the germination and growth kinetics representative curves of bacterial spores.
Statistical analysis
Each experiment was conducted in triplicate. The results of percent germination and spectrophotometer measurement were expressed in terms of mean±SD. The difference between means obtained was made using GraphPad Prism version 5.03 for Windows (GraphPad Software, San Diego, CA).
Results and Discussion
Chemical composition of essential oils
The results of the chemical composition of the essential oils are shown in Table 2. The analysis showed a particular composition of D. gossweileri essential oil. This consists essentially of two aromatic compounds with nitrogen groups: benzyl isothiocyanate (86.7%) and benzylcyanide (12.6%). This composition is similar to that obtained previously by Eyele et al. (1997) with samples of D. gossweileri harvested in Gabon and Central African Republic, but differed from that obtained by Agnaniet et al. (2003) with benzylcyanide as the major compound, which was ascribed to a possible interconversion of both compounds during extraction of essential oil.
K.I., Kovats Index; D.g., Drypetes gossweileri; O.g., Ocimum gratissimum; Z.o., Zingiber officinale; A.c., Ageratum conyzoïdes; C.r., Citrus reticulata; C.l., Citrus limon; C.c., Cymbopogon citratus; E.c., Eugenia caryophyllus; T.v., Thymus vulgaris.
GC, gas chromatography; GC-MS, gas chromatography coupled to mass spectrometry.
The other essential oils generally showed common composition with two or three predominant compounds. Essential oil of O. gratissimum was mainly composed of thymol (47.1%), γ-terpinene (16.6%), and p-cymene (14.1%). That of Z. officinale contains p-menth-1-en-4-ol (23.2%) and neral (15%). Germacreme (41.6%) and the trans-β-caryophyllene (24.6%) were predominantly present in the essential oil of A. conyzoïdes. Limonene was the major compound of Citrus reticulata and C. limon essential oils, accounting for 74.8% and 86.4%, respectively. C. citratus essential oil was mainly composed of geranial (49.2%) and neral (34.4%). Eugenol (87.7%) was the major component of the essential oil of E. caryophyllus, while thymol (45%) and p-cimene (25%) were predominantly found in the essential oil of Thymus vulgaris.
Effect of essential oils on vegetative growth and spore germination
This evaluation consisted of determining the inhibitory parameters (MIC, MICg, and MBC). The results summarized in Table 3 reveal a strong bacteriostatic effect of D. gossweileri essential oil, with MICs ranging from 0.0048 to 0.0097 mg/mL, and B. megaterium as the most sensitive microorganism. Essential oil of A. conyzoïdes also exerted bacteriostatic effect with greater activity observed on B. subtilis, which exhibited MIC of 0.312 mg/mL, while MICs of 1.25 and 2.5 mg/mL were obtained from the other essential oils. The bacteriostatic effects of essential oils were due to the presence of antimicrobial compounds in major proportion in most essential oils analyzed. Indeed, thymol, caryophyllene, limonene, geranial, eugenol, and benzyl isothiocyanate are well known for their strong antibacterial effects (Hyldgaard et al., 2012).
MIC, minimal inhibitory concentration; MBC, minimal bactericidal concentration; MICg, minimal concentration inhibiting germination of spores; (–), inactive; (*), not evaluated.
The essential oil of D. gossweileri was more active with MIC ≤0.0097 mg/mL. This MIC value was less than those previously obtained by Ngono (2008), ranging within 48.8–390.6 μg/mL on a panel of Gram-negative bacteria. This confirms the greater susceptibility of Gram-positive bacteria to essential oils than Gram negative, as reported by many studies (Hyldgaard et al., 2012). The strong antibacterial activity of D. gosssweileri essential oil is probably related to its isothiocyanate derivatives composition, known to have a very strong antimicrobial effect due to the R-N=C=S group in the molecule. This group has a high electrophilic central carbon, which can easily react with nucleophilic centers. Furthermore, it could cleave the disulfide bonds of proteins and could attack free amino acids by oxidative reaction (Wilson, 2011; Hyldgaard et al., 2012).
Concerning the inhibitory effect of spore germination, essential oil of D. gossweileri appeared to be the most active, with inhibitory effect at 0.001 mg/mL on B. subtilis spores and 0.002 mg/mL on spores of the other Bacillus and G. stearothermophilus. For the other essential oils, the MICg ranged from 0.09 to 1.5 mg/mL, with essential oils of C. limon, C. citratus, and A. conyzoïdes seeming to be more active, with MICg ≤0.37 on at least two of the four species tested. The MICg of 0.37–1.5 mg/mL are close to those obtained by Chaibi et al. (1997) on spores of B. cereus T, with the essential oils of some Moroccan plants. However, the MICg shown by D. gossweileri essential oil were generally lower, further confirming its strong antimicrobial activity. On the whole, MICg were much lower than MICs (Table 2), reflecting the fact that the inhibitory action of essential oils is more effective on germination of spores compared to vegetative growth.
Regarding the bactericidal effect, essential oils of C. reticulata, C. citratus, and E. caryophyllus generally showed an absolute effect. MBCs were observed at MIC concentration of 2.5 mg/mL. O. gratissimum and Z. officinale were also bactericidal but at higher concentrations, generally at 10 mg/mL.
Effect of combination of essential oils on vegetative growth
The effect of the combination of essential oils is summarized in Table 4. The results obtained show an improvement of the antibacterial effect due to the combinations of D. gossweileri and E. caryophyllus, A. conyzoïdes, or O. gratissimum essential oils, as shown by a decrease in MICs. Furthermore, all the combinatory effects appeared to be either additive or synergistic. Combination of D. gossweileri and E. caryophyllus proved significant activity with a synergistic effect on two (B. subtilis and G. stearothermophilus) of the four species. Synergy or additive effects observed could be explained by the combined effect of benzyl isothiocyanate, major compound of D. gossweileri, and antimicrobial compounds found in high percentage in the others essential oils such as thymol, neral, trans-β-caryophyllene, and eugenol. In addition, the increased effect of the combination could be explained by the synergistic effect of benzyl isothiocyanate and the terpene hydrocarbons within the other essential oils. Nguefack et al. (2012) reported that terpene hydrocarbons have low effect, but could swell cell membranes and enable more antimicrobial compounds such as benzyl isothiocyanate to be easily transported into the cell.
(—), inactive; MIC1, minimal inhibitory concentration of first essential oil in combination; MIC2, minimal inhibitory concentration of second essential oil in combination; MBC1, minimal bactericidal concentration of first essential oil in combination; MBC2, minimal bactericidal concentration of second essential oil in combination; FIC1, fractions inhibitory concentration of first essential oil; FIC2, fractions inhibitory concentration of second essential oil; FICindex, fractions inhibitory concentration index.
Treatment of spores with essential oil and evaluation of germination inhibition on solid medium
Figure 1 shows the curves representing the percentage germination inhibition of spores, at different concentrations of essential oils. The essential oil of D. gossweileri was the most effective. At 1.25 mg/mL, this oil was active on B. cereus, B. megaterium, and G. sterotherophilus spores, with germination inhibition percentages of 66.23, 48.54, and 51.66%, respectively. Furthermore, a total inhibition of germination was observed on all the spores at 20 mg/mL. The other essential oils had effect but at higher concentrations ranging from 10 to 20 mg/mL. However, Z. officinale essential oil was found to be active on spores of B. subtilis, with a total inhibition at 2.5 mg/mL. O. gratissimum was more active on B. cereus spores where significant activity was obtained at 2.5 mg/mL and 5 mg/mL. These results might suggest a probable irreversible effect of these three essential oils as compared to others.
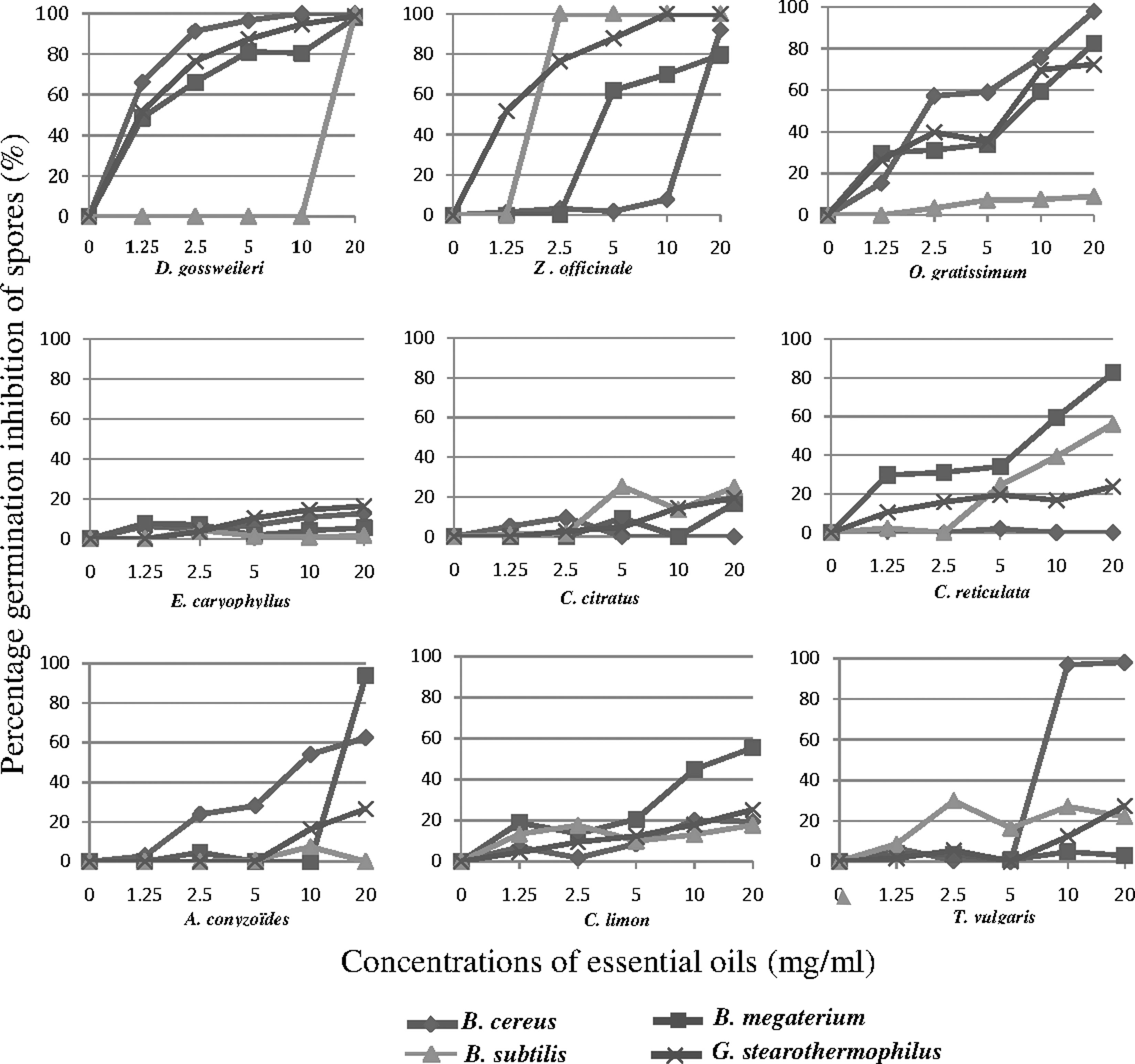
Percentage germination inhibition of spores obtained on solid medium after treatment with essential oils.
Kinetics of spores germination and outgrowth with and without essential oils
The kinetics of the germination and outgrowth of spores of B. cereus, B. megaterium, and B. subtilis were performed in the absence (control) and the presence of essential oils using spectrophotometric measurements. Figure 2 shows the kinetics obtained with spores of B. cereus. The curves show a constancy of optical densities between 0 and 120 min for both control and tested spores. At 120 and 300 min, respectively, for control and spores in the presence of C. limon essential oil, an increase of optical densities was recorded due to the start of vegetative growth. This observation was not revealed with spores in the presence of the other essential oils, suggesting that these essential oils probably inhibited the germination and/or outgrowth of spores. Complete inhibition of spore germination was observed in the presence of the essential oil of A. conyzoïdes because the optical densities remained constant during the test period. Concerning the essential oils of D. gossweileri and C. citratus, slight decreases in optical densities were recorded after 100 and 180 min, respectively, characterizing spore germination. However, after 200 min, an increase in optical densities was observed, suggesting that these germinated spores might return to their initial state of dormancy. This phenomenon called microcycle of spores was already observed with B. cereus spore by Edima et al. (2010).
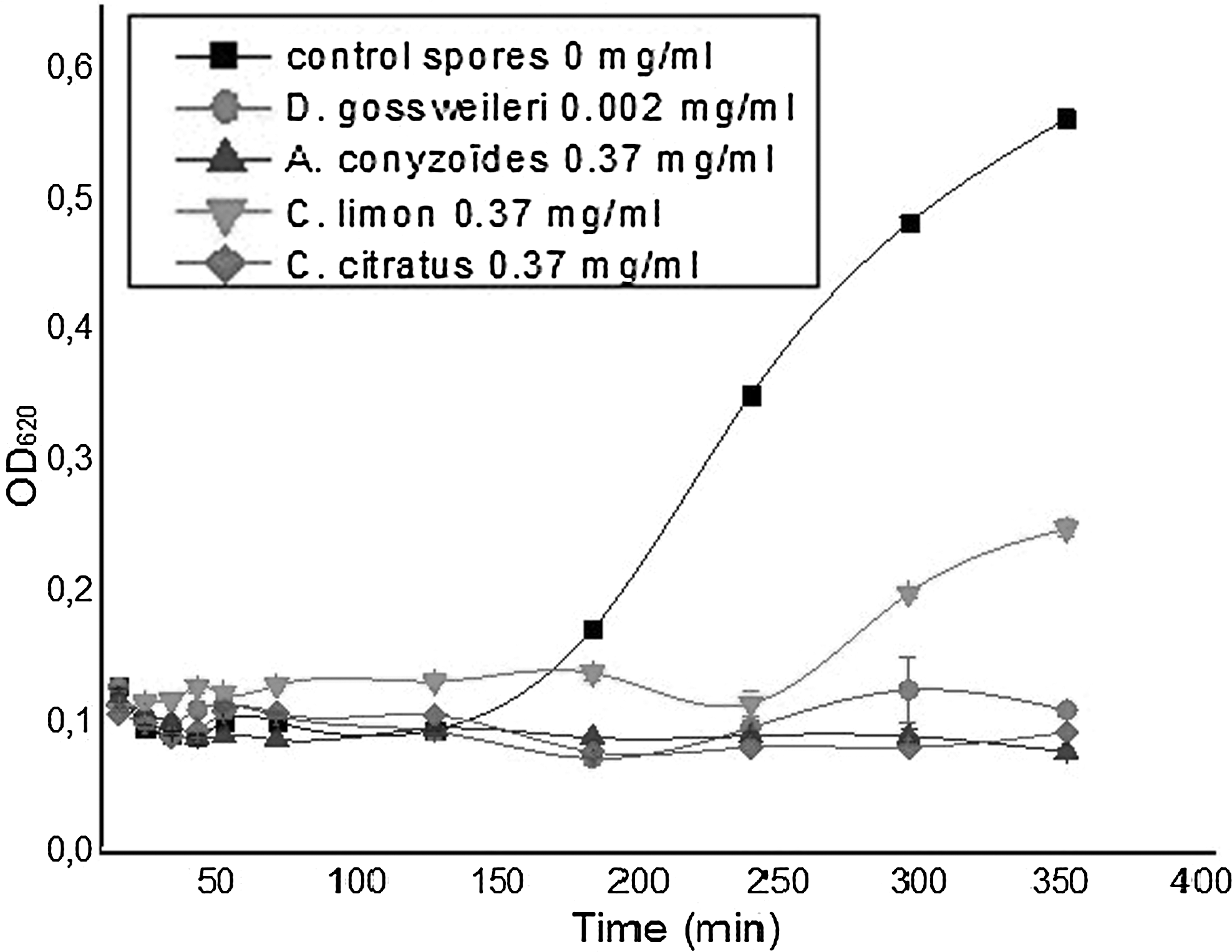
Germination, outgrowth, and growth kinetic of spores of Bacillus cereus in the presence of essential oils. OD, optical density.
Figures 3 and 4 show that the kinetics conducted on spores of B. megaterium and B. subtilis, respectively, revealed that from 0 to 40 min, the optical densities recorded are almost constant with control and tested spores. This period probably corresponds to the lag phase. From 60 min for the control and the spores with D. gossweileri essential oil, a decrease of optical densities was observed, probably due to the germination of spores. This germination step starts at 120 min with spores in the presence of the other essential oils. After 180 min, an exponential growth only with the control was observed, probably due to the start of vegetative growth. This observation is not shown with spores in the presence of essential oils, suggesting that the essential oil prevented the germination of spores.
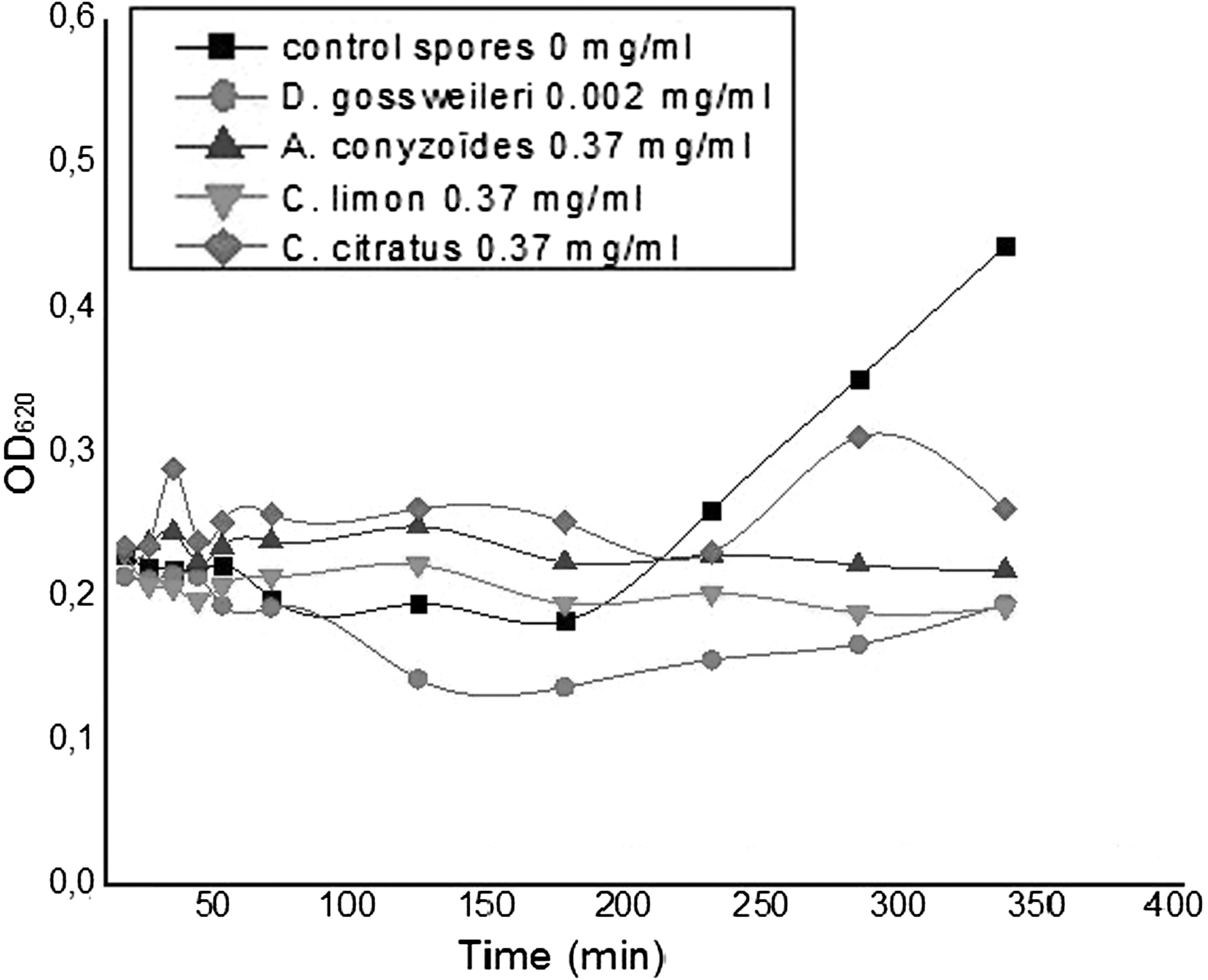
Germination, outgrowth, and growth kinetic of spores of Bacillus megaterium in the presence of essential oils. OD, optical density.
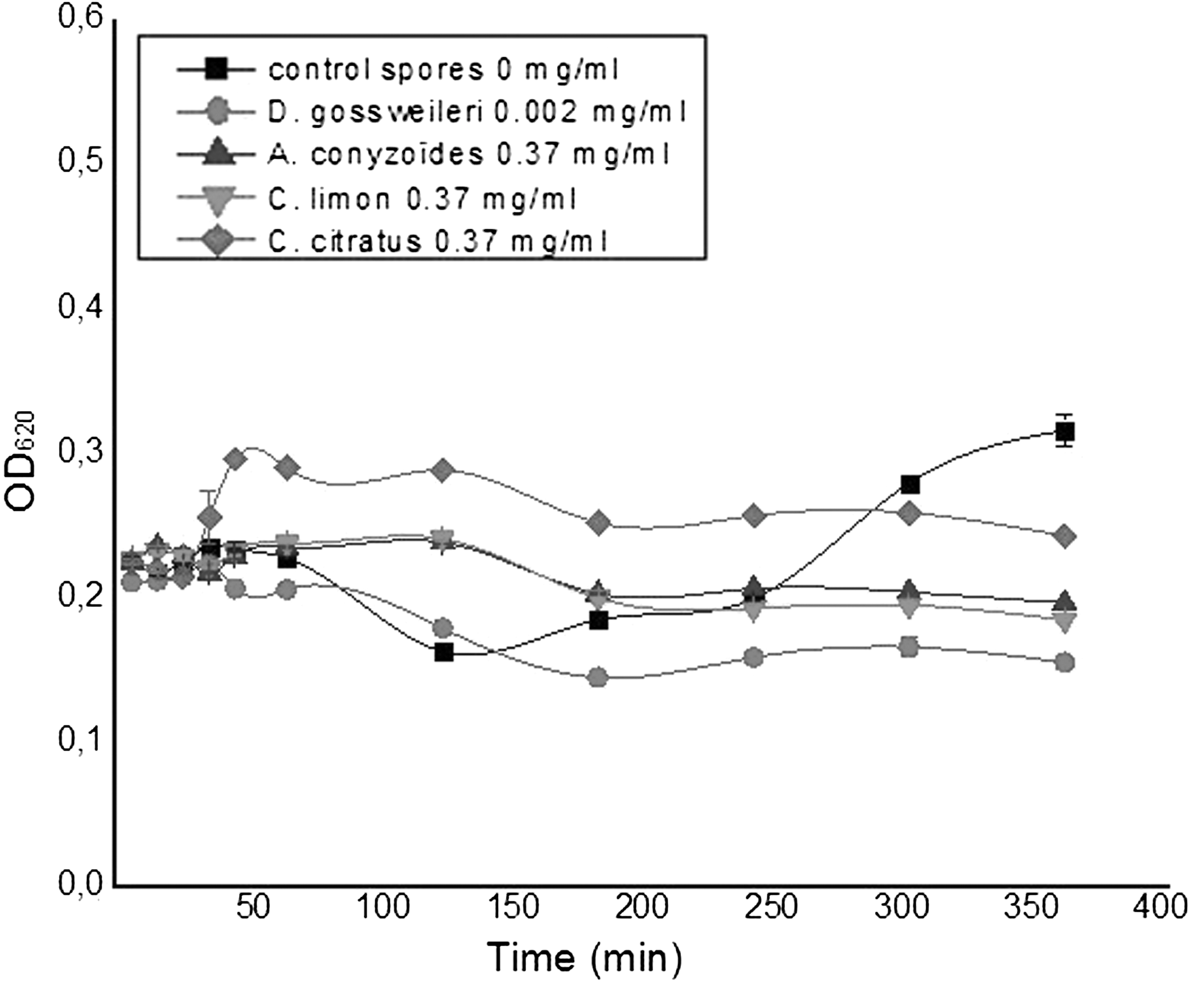
Germination, outgrowth, and growth kinetic of spores of Bacillus subtilis in the presence of essential oils. OD, optical density.
Conclusions
Essential oils used in this work have displayed the capacity to inhibit germination as well as vegetative growth of spores of Bacillus and G. stearothermophilus. Among the essential oils tested, D. gossweileri appears to be more active as compared to others. Furthermore, its combination with other essential oils improves the antibacterial effect. Thus, D. gossweileri essential oil could be suggested as a food preservative. Nevertheless, further studies must be conducted upon other bacterial spores.
Footnotes
Acknowledgments
We are grateful to Professor Chantal Menut of the Institut des Biomolécules Max Mousseron (IBMM) de Montpellier, the University of Yaounde I, and the Vasile Alecsandri University of Bacau for laboratory facilities. We thank the Romanian Government, the Agence Universitaire de la Francophonie (AUF), and the Service de la Cooperation et de l'Action Culturelle (French Embassy in Cameroon) for funding this project. We also thank the Microbiology Laboratory Institute of Food Research of Reading and the Institut Appert, Paris for providing us with bacterial spore strains.
Disclosure Statement
No competing financial interests exist.