
Abstract
Foodborne bacteria are the leading cause of food spoilage and other related diseases. In the present study, the antibacterial activity of bio-oil (BO) manufactured by fast pyrolysis of pinewood sawdust (Pinus densiflora Siebold and Zucc.) against two disease-causing foodborne pathogens (Bacillus cereus and Listeria monocytogenes) was evaluated. BO at a concentration of 1000 μg/disc was highly active against both B. cereus (10.0–10.6 mm-inhibition zone) and L. monocytogenes (10.6–12.0-mm inhibition zone). The minimum inhibitory concentration (MIC) and minimum bactericidal concentration values of BO were 500 and 1000 μg/mL, respectively, for both pathogens. At the MIC concentration, BO exhibited an inhibitory effect on the viability of the bacterial pathogens. The mechanism of action of BO revealed its strong impairing effect on the membrane integrity of bacterial cells, which was confirmed by a marked release of 260-nm absorbing material, leakage of electrolytes and K+ ions, and reduced capacity for osmoregulation under high salt concentration. Scanning electron microscopy clearly showed morphological alteration of the cell membrane due to the effect of BO. Overall, the results of this study suggest that BO exerts effective antibacterial potential against foodborne pathogens and can therefore potentially be used in food processing and preservation.
Introduction
F
Natural compounds found in plants and various byproducts have been shown to possess enough potential to serve as a source of antimicrobial agents against food pathogens (Deans and Ritchie, 1987; Miksusanti et al., 2008; Bajpai et al., 2013; Cattelan et al., 2013; Diao et al., 2013, 2014; Dussault et al., 2014). In general, bio-oil (BO) is prepared from various lignocellulosic biomasses such as woody biomass, and agricultural residues by a thermochemical pyrolysis process conducted in the absence of oxygen to convert the biomass into liquid BO, bio-char, and gases. During this process, the polymeric constituents of organic material are thermally degraded into numerous small molecules and converted to volatile vapors to form BO (Bridgwater et al., 1999). Plant-based BO could be an alternative source that can act against pathogenic bacteria responsible for food-related diseases. Few studies have investigated the antimicrobial effect of BO against pathogenic microbes (Bedmutha et al., 2011; Kim et al., 2012; Phukan et al., 2013).
In this study, the effectiveness of BO on selected foodborne pathogens was evaluated using an in vitro model. Furthermore, the mechanism of antibacterial action was investigated by determining the leakage of potassium ion and cellular materials, and their effect on salinity tolerance.
Materials and Methods
Production of BO
Pinewood sawdust was used to produce BO in a lab-scale fluidized-bed fast pyrolyzer. The sawdust was air dried to approximately 7% moisture content and ground to powder (particle size of 0.5 mm). Fast pyrolysis of the powdered sample was performed at 500°C with a residence time of 1.3 s (Hwang et al., 2013). The yield of BO was 60.5 wt%, while that of char and gas was 12.9 wt% and 26.6 wt%, respectively. The produced BO was characterized by several analytical methods. Specifically, a Karl–Fischer titration with five hydranal composite solutions was used to determine the water content. The elemental composition (carbon, hydrogen, nitrogen, and oxygen) was analyzed using an elemental analyzer (CHNS-932, LECO Corporation, St. Joseph, MI). The kinetic viscosity at 40°C, pH, and calorific value of BO were determined using a capillary viscometer, pH meter, and bomb calorimeter (Parr 6400; Parr Instrument Company, Moline, IL), respectively. BO was kept at 4°C until further analysis.
Antibacterial screening of BO
Foodborne pathogenic bacteria
Four different foodborne bacterial pathogens from two species including Bacillus cereus (ATCC 10876 and ATCC 13061) and Listeria monocytogenes (ATCC 19114 [serotype 4a] and ATCC 19115 [serotype 4b]) were selected for the antibacterial evaluation. All strains were obtained from the American Type Culture Collection (ATCC, Manassas, VA) and maintained in nutrient broth (NB) media with glycerol at –80°C until use.
Determination of antibacterial activity of BO by disc-diffusion assay
The antibacterial activity of BO was evaluated using a standard disc-diffusion assay (Bajpai et al., 2013). Filter-paper discs (6 mm, Advantec, Toyo Roshi Kaisha Ltd., Tokyo, Japan) impregnated with 1000 μg per disc of BO dissolved in 5% dimethyl sulfoxide (DMSO) were used for the assay. Kanamycin (Sigma-Aldrich Co., St. Louis, MO) at 40 μg/disc and 5% DMSO were used as positive and negative control, respectively. The diameters of zone of inhibition were recorded. The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) were determined by the twofold serial dilution method (Bajpai et al., 2013).
Effect of BO on viability of bacteria
The bactericidal effects of BO on the cell viability were evaluated by a standard viability test (Bajpai et al., 2009). Overnight-grown cultures of B. cereus ATCC 13061 (BC) and L. monocytogenes ATCC 19115 (LM) were prepared in NB media. One milliliter of each culture was centrifuged at 10,000 rpm for 10 min, and then pellets were collected and resuspended with 1 mL of phosphate-buffered saline. For viable counts, each tube containing resuspended bacterial suspension (approximately 107 colony-forming units [CFU]/mL) in 900 μL of NB was inoculated with 100 μL of BO at MIC and incubated at 37°C. Next, 100 μL of both control and treatment were taken out at 2-h intervals for 8 h, appropriately diluted, and spread on nutrient agar (NA) plates. The plates were incubated at 37°C for 24 h, after which the colonies were counted.
Effect of BO on release of 260-nm absorbing materials from bacteria
The release of 260 nm absorbing cellular materials from BC and LM was measured according to standard procedure (Carson et al., 2002). Briefly, BO at MIC was added to 2 mL of bacterial inoculum in sterile peptone water (0.1 g/100 mL) and incubated at 37°C. After 0, 60, and 120 min, cells were centrifuged at 3500 rpm and the absorbance of obtained supernatant was measured at 260 nm in a spectrophotometer (ASP 3700, ACT Gene Inc., Piscataway, NJ). Bacterial culture in sterile peptone water without BO was used as control. The results were expressed in terms of optical density (OD) at 260 nm at each interval with respect to ultimate time.
Effect of BO on cell membrane permeability of bacteria
The permeability of bacterial membranes was expressed in terms of relative electrical conductivity and determined as previously described (Kong et al., 2008). Prior to the assay, cultures of BC and LM were incubated at 37°C for 10 h. Next, the cultures were centrifuged at 5000 rpm for 10 min, then washed with 5% glucose solution (wt/vol) until their electrical conductivities were close to that of 5% glucose solution to induce an isotonic condition. BO at MIC was added to 5% glucose solution and the electrical conductivities of the mixtures were determined (L1). BO at MIC was added to the isotonic bacterial solution, mixed completely, and incubated at 37°C for 8 h. The conductivity of the bacterial solution was measured at 2-h intervals for 8 h (L2). The conductivity of bacteria in 5% glucose killed by boiling water for 5 min served as control (L0). The relative electrical conductivity was measured using a conductivity meter (Con 6; LaMotte Company, Chestertown, MD). The permeability of bacteria membrane was calculated according to the following formula:
Effect of BO on efflux of potassium ions from bacteria
The efflux of potassium (K+) ions from bacterial cells was determined by standard procedures (Lee et al., 2002). The concentration of free K+ ions in bacterial suspensions of BC and LM was measured after exposure to MIC of BO in peptone water (0.1 g/100 mL) and incubation at 37°C. The measurement was done at each 2-h time interval by a photometric procedure using Kalium/Potassium kit (Quantofix, GmbH, Wiesbaden, Germany). Bacterial suspension without BO was used as control. Results were expressed as the amount of extracellular free K+ (mg/L) released into the growth media at each interval of incubation time.
Effect of BO on loss of salt tolerance capacity of bacteria
The ability of BC and LM treated with BO to grow on NA supplemented with a high concentration of NaCl was investigated (Miksusanti et al., 2008). BO at MIC was added to overnight culture of BC and LM, then incubated at 37°C for 60 min. The samples were serially diluted and inoculated on NA plates supplemented with different concentrations of NaCl (0, 2.5, 5, and 10%). Bacterial samples without BO were used as control. Following incubation, the number of colony-forming units on each NA-NaCl plate was compared to that on the NA plates, and the results were expressed as Log CFU/mL.
Scanning electron microscopy analysis
Scanning electron microscopy (SEM) was conducted to investigate the effects of BO on the morphology of bacterial cells (Bajpai et al., 2009). For analysis, two vials each of one control and one treatment of overnight-grown culture of BC and LM were centrifuged and then washed with 50 mM phosphate buffer (pH 7.2), after which the culture was spread over a glass slide, fixed with 2.5% glutaraldehyde, and dehydrated using different concentrations (50–100%) of ethanol. Next, the ethanol was removed and t-butanol was added. Finally, the specimen was sputter coated with platinum and observed using a scanning electron microscope (S-4110, Hitachi, Tokyo, Japan).
Statistical analysis
All experiments were carried out in triplicate and the results expressed as the mean±standard deviation. Statistical analysis software version 9.4 (SAS Inc., Cary, NC) was used for analysis of variance.
Results
Determination of antibacterial activity of BO
BO exerted greatest inhibitory effect against LM (12-mm inhibition zone), followed by B. cereus ATCC 10876 (10.6-mm inhibition zone) (Table 1). Kanamycin, the positive control, showed higher antibacterial activity than BO, whereas the negative control (DMSO) had no inhibitory effect on the tested pathogens (Table 1). The MIC and MBC values of BO against all 4 foodborne bacteria were 500 and 1000 μg/mL, respectively.
Data are expressed as the mean±SD. Values in the same column with different letters are significantly different.
Bio-oil at 1000 μg/disc.
Kanamycin at 40 μg/disc.
Dimethyl sulfoxide 5%.
Effect of BO on viability of bacteria
In order to elucidate the mechanism of action of BO on the bacterial cell, one strain from each species (BC and LM) was selected. The effect of BO on bacterial cell viability was observed by change in cell count of both the bacteria exposed to MIC of BO compared to the untreated bacteria in a time-dependent manner (Fig. 1A and B). Specifically, there was no significant decrease in cell viability from 0 min to 4 h of exposure to BO; however, the cell viability decreased greatly and growth completely ceased after 6 h of incubation of both pathogens (Fig. 1). The control plate showed no change in viable count with respect to incubation period.

Effect of bio-oil manufactured from pyrolysis of Pinus densiflora at the minimum inhibitory concentration on the viability of Bacillus cereus ATCC 13061
Effect of BO on release of 260-nm absorbing materials
BC and LM treated with BO showed a continuous increase in OD values at 260 nm with respect to incubation time (Fig. 2). However, there was no distinct change in the OD of untreated control in both of the bacteria. BO was found to have a greater effect on LM than BC at all-time intervals (Fig. 2).
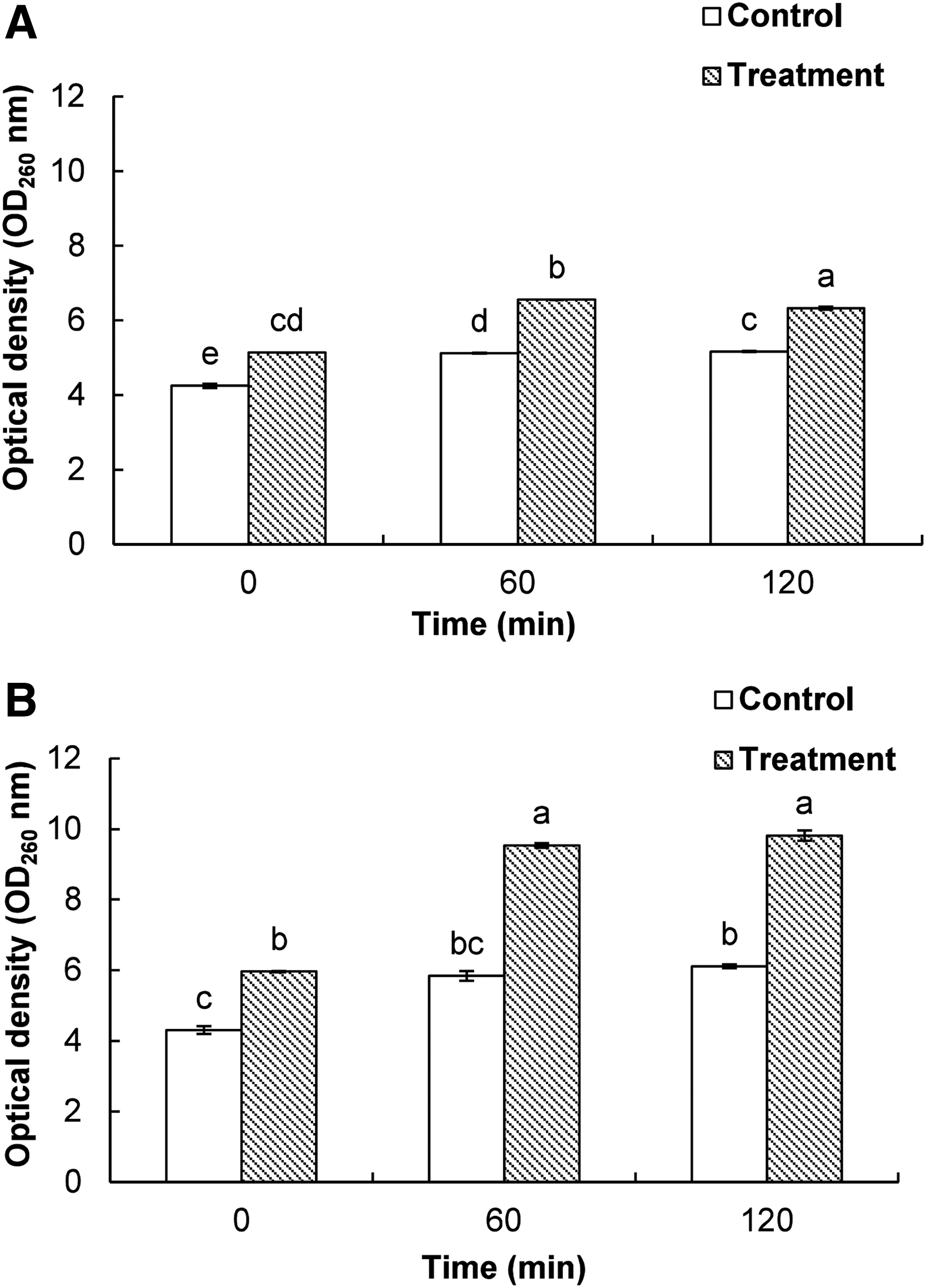
Effect of bio-oil manufactured from pyrolysis of Pinus densiflora at the minimum inhibitory concentration on the release rate of 260 nm absorbing material from Bacillus cereus ATCC 13061
Effect of BO on cell membrane permeability and leakage of K+ ion
Figure 3 shows the effect of BO on the membrane permeability of BC and LM in terms of their relative electrical conductivity. A continuous increase in the relative electrical conductivity of both BC and LM treated with BO was observed with respect to time interval. BO was found to be more effective against LM relative to BC (Fig. 3). The untreated control sample showed no relative electrical conductivity.
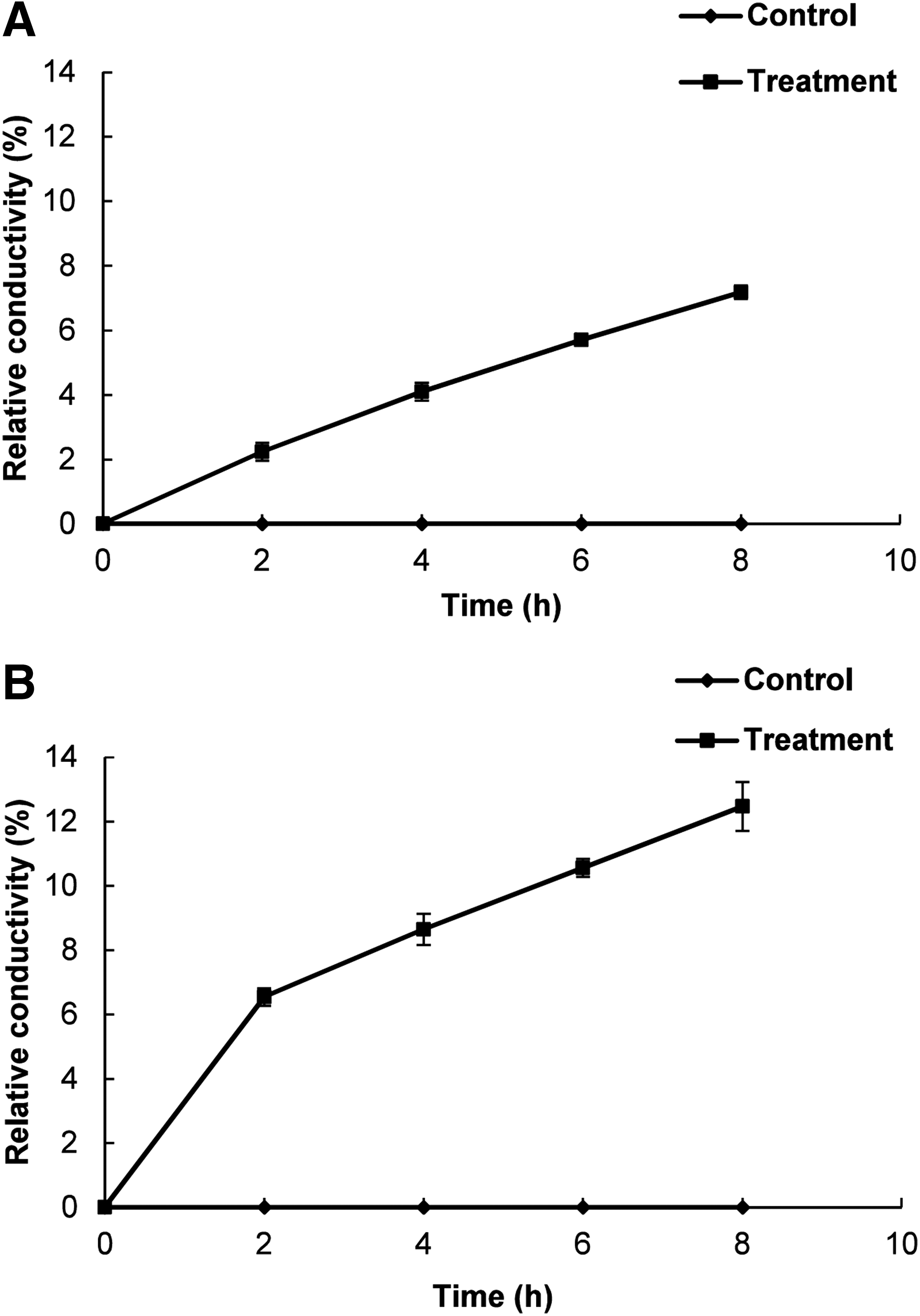
Effect of bio-oil manufactured from pyrolysis of Pinus densiflora at the minimum inhibitory concentration on the cell membrane permeability of Bacillus cereus ATCC 13061
As shown in Figure 4, both BC and LM treated with BO induced an efflux of intracellular K+ ions to the outside of the cell with respect to time specifically after 6 h of incubation, whereas the untreated control showed no release of K+ in both of the bacteria, throughout the incubation period.
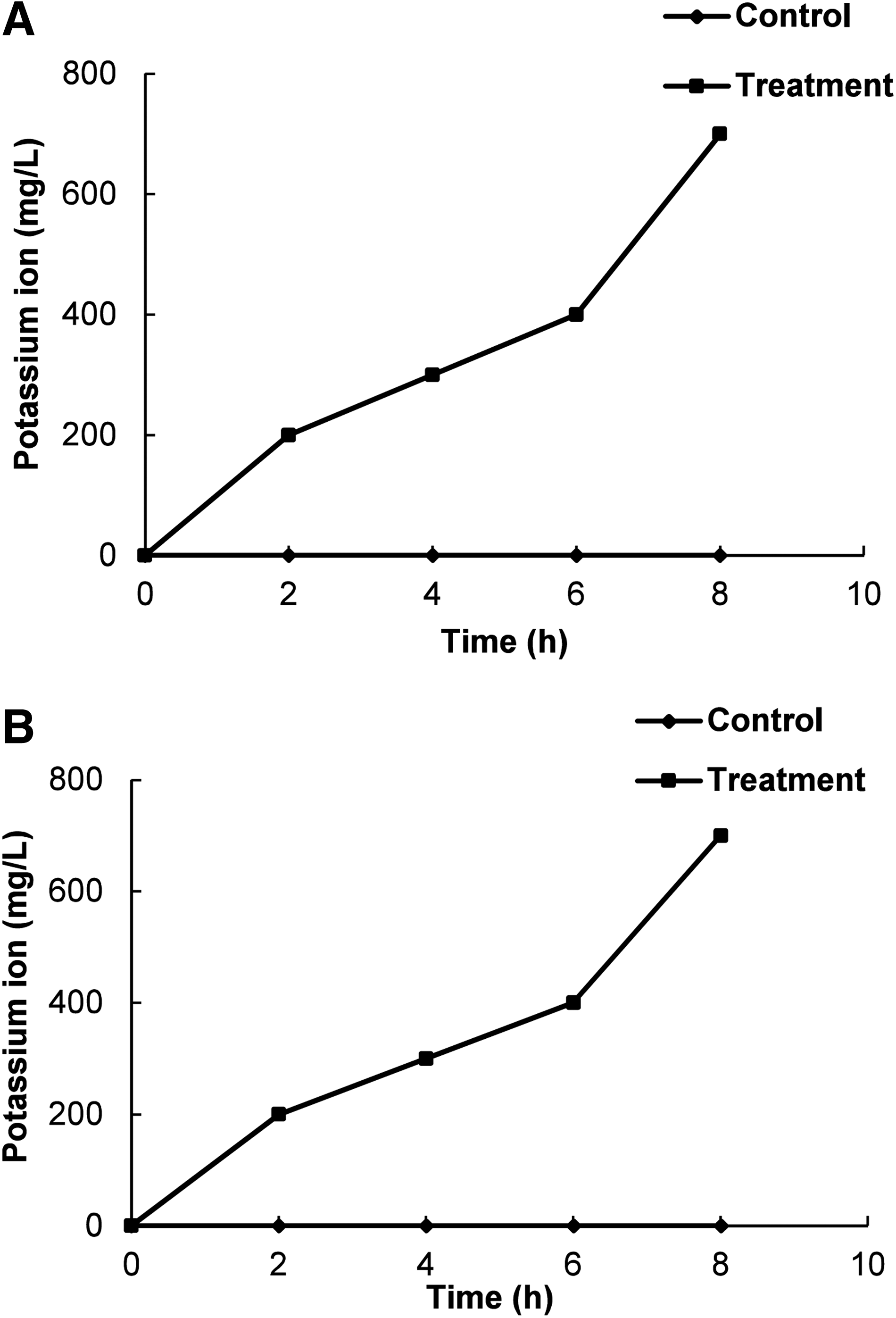
Effect of bio-oil manufactured from pyrolysis of Pinus densiflora at the minimum inhibitory concentration on the leakage of potassium ions from Bacillus cereus ATCC 13061
Effect of BO on loss of salt tolerance
When both of the bacteria pretreated with BO (treatment) or DMSO (control) were allowed to grow on NA plates supplemented with NaCl (0, 2.5, 5, and 10%), there was a gradual decrease in the number of colony-forming units at each concentration in NaCl-NA plates (Fig. 5). Moreover, the number of bacteria treated with BO was significantly lower than that of the DMSO-treated bacteria at each concentration of NaCl (Fig. 5). Neither treated nor control samples grew in the presence of 10% NaCl.
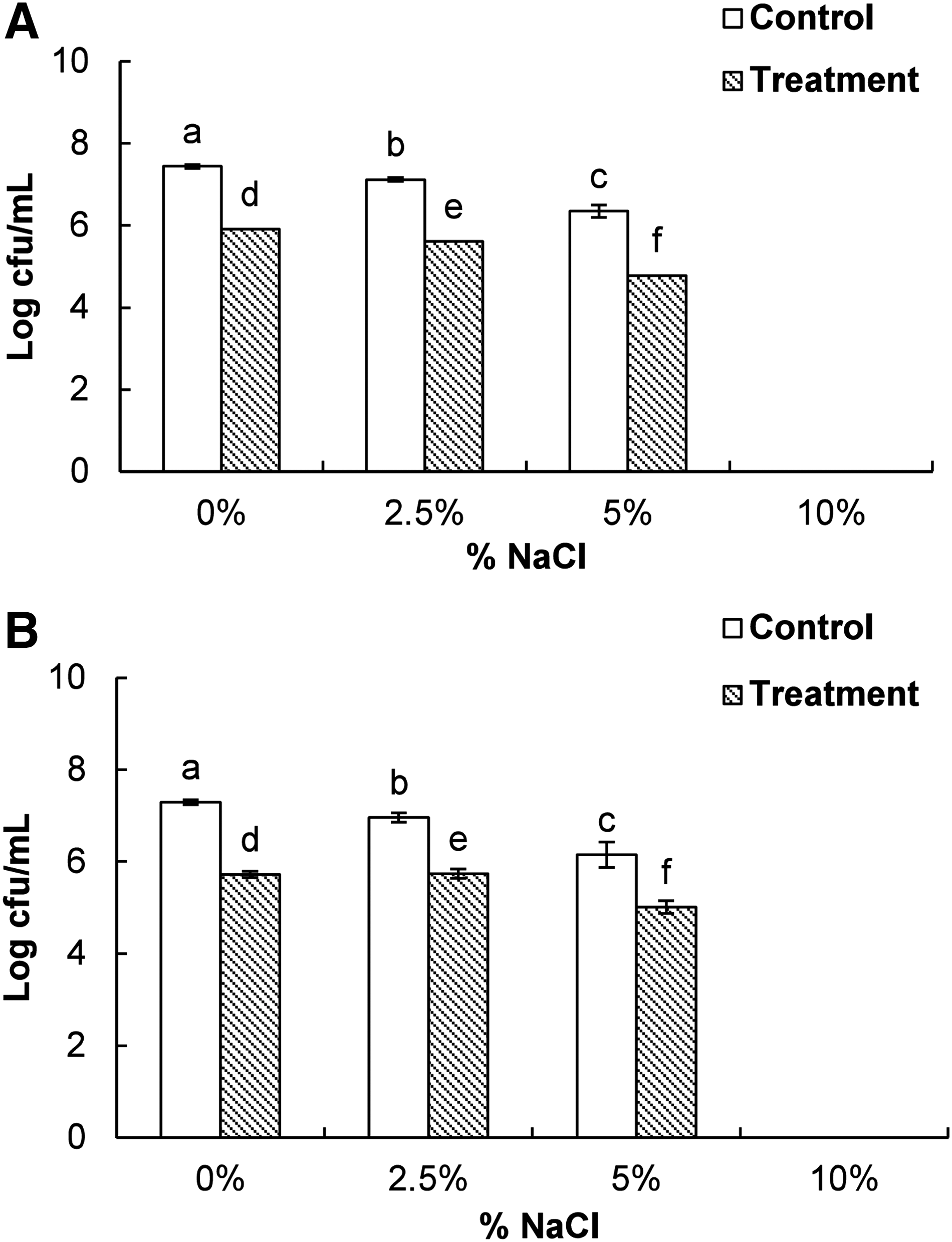
Effect of bio-oil manufactured from pyrolysis of Pinus densiflora at the minimum inhibitory concentration on loss of salt tolerance of Bacillus cereus ATCC 13061
SEM analysis
There were distinct morphological alterations on the surface of BC and LM treated with BO (Fig. 6). Specifically, the cells showed irregular shapes in contrast to the smooth and regular surface of the untreated control (Fig. 6).
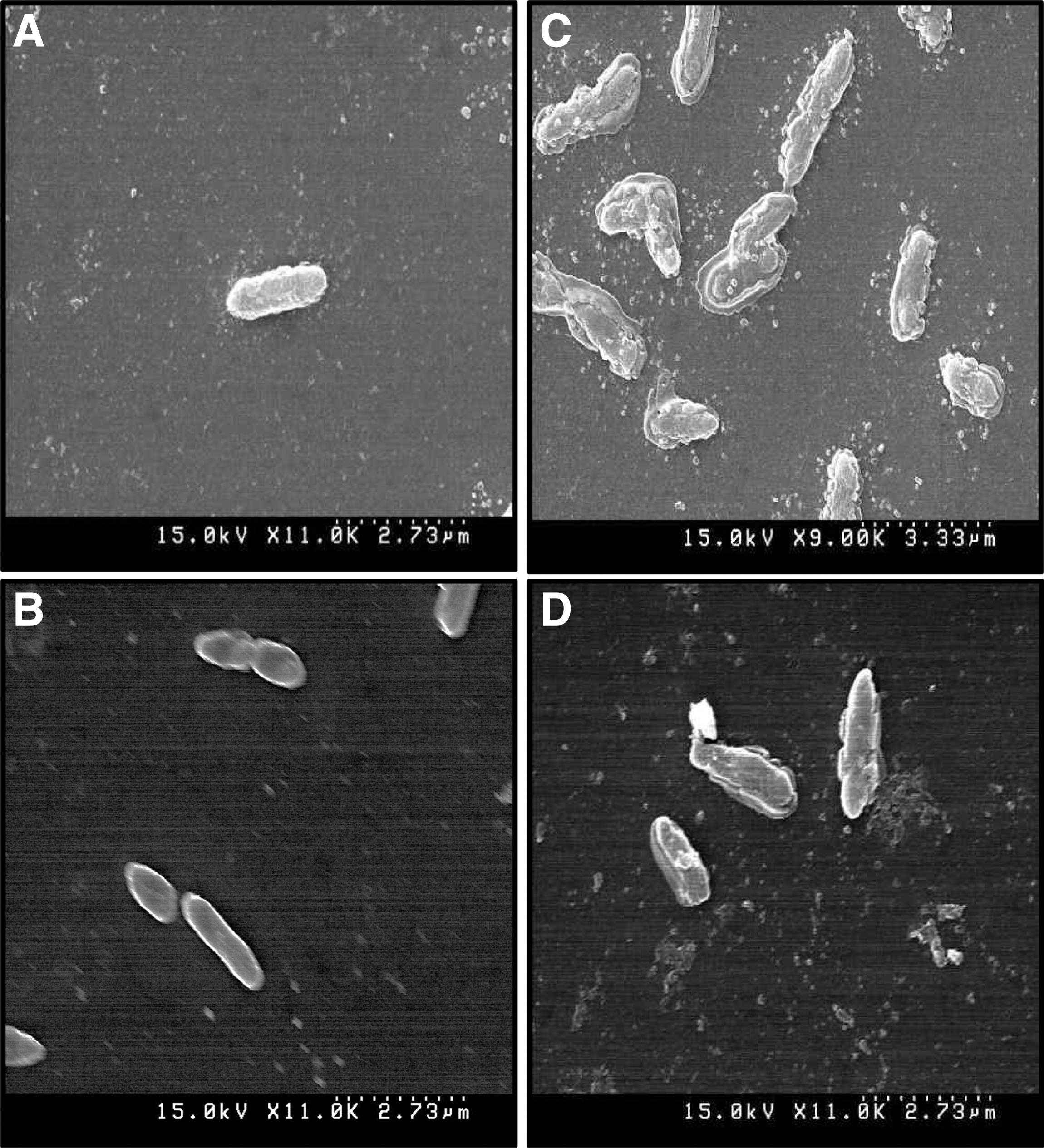
Scanning electron microscopy of Bacillus cereus ATCC 13061
Discussion
Food-related problems such as food contamination, foodborne illness, and food spoilage are related to a group of bacterial pathogens that decreases the nutritional quality of food (Diao et al., 2014). The use of natural preservatives and chemicals derived from natural sources in food preservation has garnered much attention in the food and pharmaceutical industries (Li and Yi, 2003; Sharma et al., 2013). Medicinal plants and their products have long been used to preserve foods (Balchin and Deans, 1997); hence, in the present study, the antibacterial effects of BO were investigated.
BOs have been reported to possess certain active compounds effective against various groups of pathogenic microorganisms (Bedmutha et al., 2011; Kim et al., 2012; Phukan et al., 2013). In the present study, BO was found to be effective against all four strains of B. cereus and L. monocytogenes (Table 1). The effective potential of BO might be due to its hydrophobic nature, which easily penetrates the bacterial cell wall due to their structure, causing cell death. Specifically, the hydrophobic nature of BO might also have enabled its accumulation in the bacterial cell membrane, resulting in leakage of intracellular materials and excessive loss of contents, eventually causing cell death (Sikkema et al., 1994). BO contains a large amount of oxygenated compounds such as phenolics, which might exert an antibacterial effect by forming agglomerated clusters against the bacteria (Kim et al., 2012). A proposed mechanism of action of the antibacterial property of BO against Gram-positive foodborne bacterial pathogens is presented in Figure 7.
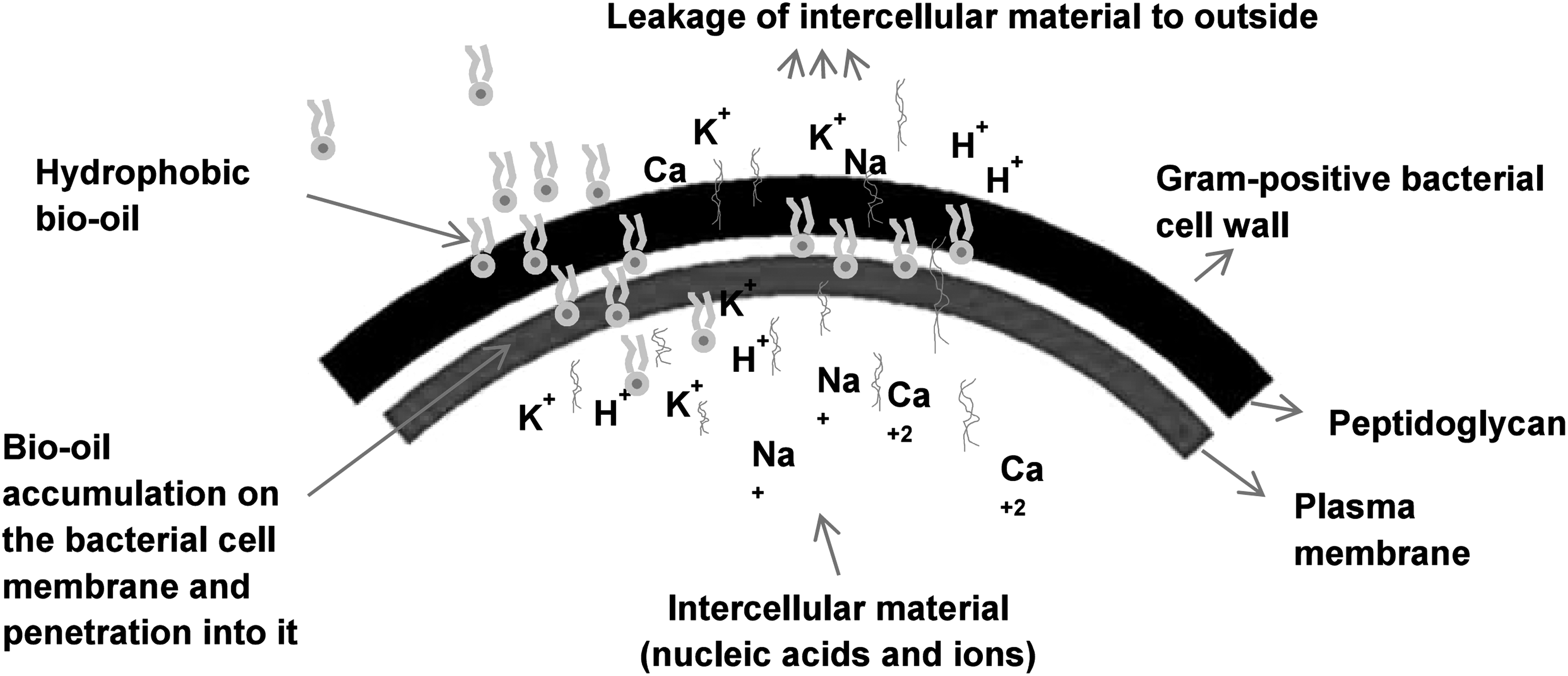
Proposed mechanism of the mode of antibacterial action of bio-oil manufactured from pyrolysis of Pinus densiflora against Gram-positive foodborne bacteria.
The antibacterial modes of action of BO against BC and LM were evaluated by various assays. Exposure of BO to BC and LM had a detrimental effect on their viability, and the cells were completely destroyed after exposure for 6 h (Fig. 1).
Following exposure of the bacterial cells to BO, there was a significant increase in the OD at 260 nm for both bacterial cultures (Fig. 2), indicating significant loss of intracellular materials such as nucleic acids, metabolites, and ions into the suspensions that were absorbed at 260 nm (Cox et al., 2003). This was likely a result of cellular leakage in response to deformities such as holes in the cell surface caused by BO (Cox et al., 2003; Zhu et al., 2005). Many antibacterial compounds act by facilitating leakage of cytoplasm and its coagulation, which affects the integrity and function of the cell, eventually resulting in death (Denyer, 1990). The plasma membranes of bacteria provide a barrier to the passage of small ions such as H+, K+, Na+, and Ca2+ and allow cells and organelles to control entry and exit of different compounds. This permeability barrier role of the cell membrane is essential to many cellular functions, including maintenance of the energy status of the cell, other membrane-coupled energy-transuding processes, solute transport, regulation of metabolism, and control of turgor pressure (Cox et al., 1998). The permeability of the cell membrane of BC and LM was significantly influenced by the addition of BO to the culture solution, as indicated by the increase in the relative electrical conductivity and K+ ion in the bacterial solution (Figs. 3 and 4). The leakage of K+ ions to the outer environment is an indication of cellular disruption and cellular lysis (Bajpai et al., 2013), which was also the same in the present case.
Sublethal injury of microbial cell membranes may also affect the membrane's ability to osmoregulate the cell adequately or exclude toxic materials (Carson et al., 2002). Consequently, the loss of tolerance to salts or other potentially toxic compounds might be used to reveal membrane damage in sublethally injured bacteria (Miksusanti et al., 2008). Treatment of BC and LM with BO reduced the ability of survivors to form colonies on media containing NaCl at 2.5% and 5.0% (Fig. 5), indicating that osmoregulation failed in both bacteria under high salt concentrations.
BC and LM treated with BO underwent several morphological changes, including a distinct loss of cellular integrity and deformations on the surface of the bacterial cell (Fig. 6). When coupled with the results of a previous study, these findings suggested that the active compounds present in BO might have bound to the surface of bacterial cells, penetrated the cytoplasmic membrane, and resulted in disruption of the cell structure (Fig. 7) (Zhu et al., 2005).
Conclusions
In conclusion, this study showed that BO manufactured from P. densiflora Siebold and Zucc. possesses significant antibacterial activity against B. cereus and L. monocytogenes. The mechanism of bactericidal activity of BO was permeabilization of the cell membrane via a membrane-disruption effect. Overall, the results indicate that BO exerts an antibacterial effect and can potentially be used as a natural source of antibacterial agents in food processing and preservation against foodborne pathogens.
Footnotes
Acknowledgment
This research was conducted under a research fund provided by Yeungnam University.
Disclosure Statement
No competing financial interests exist.