
Abstract
Salmonella Enteritidis (SE) is a major foodborne pathogen responsible for causing gastrointestinal infections in humans, predominantly due to the consumption of contaminated eggs. In layer hens, SE colonizes the intestine and migrates to various organs, including the oviduct, thereby leading to egg yolk and shell contamination. This study investigated the efficacy of caprylic acid (CA), a medium-chain fatty acid, in reducing SE colonization and egg contamination in layers. Caprylic acid was supplemented in the feed at 0%, 0.7%, or 1% (vol/wt) from day 1 of the experiment. Birds were challenged with 1010 log colony-forming units (CFU)/mL of SE by crop gavage on day 10, and re-inoculated (1010 log CFU/mL) on day 35. After 7 days post first inoculation, eggs were collected daily and tested for SE on the shell and in the yolk separately. The birds were sacrificed on day 66 to determine SE colonization in the ceca, liver, and oviduct. The consumer acceptability of eggs was also determined by triangle test. The experiment was replicated twice. In-feed supplementation of CA (0.7% and 1%) to birds consistently decreased SE on eggshell and in the yolk (p<0.05). Supplementation of CA at 1.0% decreased SE population to ≈14% on the shell and ≈10% in yolk, when compared to control birds, which yielded ≈60% positive samples on shell and ≈43% in yolk. Additionally, SE populations in the cecum and liver were reduced in treated birds compared to control (p<0.05). No significant difference in egg production, body weight, or sensory properties of eggs was observed (p>0.05). The results suggest that CA could potentially be used as a feed additive to reduce eggborne transmission of SE.
Introduction
S
Despite adopting various pre- and postharvest control measures, SE remains a major cause of eggborne disease outbreaks (EFSA, 2014). The United States Centers for Disease Control and Prevention (CDC) recently reported that foodborne salmonellosis did not decrease significantly in the last decade, underscoring the need for novel approaches to control Salmonella (CDC, 2012). Due to the increasing evidence linking human salmonellosis with consumption of eggs, the U.S. Food and Drug Administration (FDA) in 2009 issued a final rule requiring egg producers to implement measures to prevent SE from contaminating eggs on the farm and further growth during storage and transportation (FDA, 2009).
The cecum is the primary site of SE colonization in chickens (Allen-Vercoe and Woodward, 1999; Stern, 2008), and cecal carriage of the pathogen leads to contamination of ovaries by the transovarian route (Gantois et al., 2009). Moreover, contamination of egg contents (yolk, albumen, and eggshell membranes) by SE can occur prior to oviposition (Miyamoto et al., 1997; Okamura et al., 2001), where Salmonella colonized in reproductive organs invades and multiplies in the granulosa cells of the pre-ovulatory follicles in the reproductive tract (Thiagarajan et al., 1994, 1996). Since SE colonization of the ceca results in the transovarian spread of the bacterium and subsequent eggborne transmission, interventions implemented at the flock level to decrease the pathogen prevalence in birds represent a viable strategy for reducing human salmonellosis from egg consumption (Altekruse et al., 1993; Keller et al., 1995; Clavijo et al., 2006; Namata et al., 2008; Gantois et al., 2008, 2009).
Although several approaches, including feeding chickens with competitive exclusion bacteria (Mead, 2000; Stern et al., 2001), bacteriophages (Carrillo et al., 2005), organic acids (Byrd et al., 2001; Heres et al., 2004), oligosaccharides (Schoeni and Wong, 1994; Spring et al., 2000; Fernandez et al., 2002), antibiotics (Chadfield and Hinton, 2004), and vaccination of birds (Inoue et al., 2008) have been investigated for reducing pathogen colonization in chickens, limited efficacy of the aforementioned approaches along with concerns for the development of multidrug resistance in Salmonella have triggered research investigating the potential of natural antimicrobials to control pathogens (Abee et al., 1995; Salamci et al., 2007). Lipids and their esters have been studied extensively in the past for their microbicidal activity (Bergsson et al., 1998; Nair et al., 2005). Free fatty acids, especially medium-chain fatty acids (MCFAs), are bactericidal against Gram-positive and Gram-negative bacteria (Nakai and Siebert, 2003; Dierick et al., 2004). Caprylic acid (CA, octanoic acid) is a natural, 8-carbon MCFA present in coconut oil, breast milk, and bovine milk (Jensen, 2002; Sprong et al., 2001). It is classified as generally recognized as safe (GRAS) by the FDA (CFR 184.1025). Our previous research indicated that supplementation of CA through feed reduced Campylobacter jejuni and SE carriage in broiler chickens (Solis de los Santos et al., 2008, 2009; Johny et al., 2009). Prior research from our laboratory also revealed that CA reduced SE invasion of avian intestinal epithelial cells by downregulating critical colonization-associated genes in the pathogen (Kollanoor-Johny et al., 2012). The present study was undertaken to investigate the prophylactic efficacy of CA as a feed supplement in reducing SE colonization and eggborne transmission in layer chickens. Moreover, the effect of CA supplementation on consumer acceptability of eggs was studied.
Materials and Methods
Bacterial strains and dosing
A four-strain mixture of SE strains isolated from chickens (obtained from the Connecticut Veterinary Diagnostic Medical Laboratory, University of Connecticut) was used to inoculate the birds. The SE isolates were SE-12 (chicken liver, phage type 14b), SE-21 (chicken intestine, phage type 8), SE-28 (chicken ovary, phage type 13a), and SE-31 (chicken gut, phage type 13a). Each strain was pre-induced for resistance to 50 μg/mL of nalidixic acid (NA; Sigma Aldrich, St. Louis, MO) for selective enumeration (Kollanoor-Johny et al., 2012). One hundred microliters of each NA-resistant strain was cultured separately in 10 mL tryptic soy broth (TSB; Difco, Becton Dickinson, Sparks, MD) overnight, transferred to flasks containing 100 mL TSB supplemented with 50 μg/mL of NA, and incubated overnight at 37°C with shaking (100 rpm). Equal volumes of the SE cultures were combined and centrifuged at 3600×g for 15 min at 4°C. The pellet was washed and resuspended in 100 mL of phosphate-buffered saline (PBS, pH 7.0), and used as the inoculum (≈1010 CFU/mL). The bacterial count in the individual cultures and the four-strain cocktail was confirmed by plating 0.1-mL portions of appropriate dilutions on xylose lysine desoxycholate agar (XLD; Difco) plates containing NA (XLD-NA), and incubating the plates at 37°C for 24 h.
Experimental birds and housing
All experiments with chickens were approved by the Institutional Animal Care and Use Committee at the University of Connecticut. Forty-week-old Salmonella-free layer hens (single comb, white leghorn) were procured from University of Connecticut poultry farm, and allocated to floor pens with ad libitum nonmedicated feed, Salmonella-free water, age-appropriate ambient temperatures, and bedding at the Isolation Facility of University of Connecticut.
Experimental design
Two experiments with CA were conducted, wherein 40-wk-old hens were randomly allocated to 6 treatments. CA from Sigma Aldrich (99% purity, SAFC grade) was used for all the experiments. Briefly, the treatments included a negative control (no SE challenge and no supplemental CA), low-dose compound control (no SE challenge, but 0.7% supplemental CA vol/wt), high-dose compound control (no SE challenge, 1% supplemental CA vol/wt), a positive control (SE challenge, but no supplemental CA), a low-dose treatment (SE challenge and 0.7% supplemental CA), and a high-dose treatment (SE challenge and 1% supplemental CA). On day 0, two birds from each experimental group were randomly selected and sacrificed to confirm that the birds were initially devoid of any Salmonella. In addition, cloacal swabs were collected from all birds prior to CA supplementation for Salmonella testing. CA was supplemented in the feed for 66 days starting from day 0. An appropriate amount of CA was added in feed and mixed thoroughly to obtain concentrations of 0.7% and 1.0% in the feed. On day 10, birds in the positive control, low-dose, and high-dose treatments were challenged with the 4-strain mixture of SE (5×10 log10 CFU/mL per bird) by crop gavage. After 3 days of SE challenge (day 13), 3 birds from each treatment group were sacrificed to determine pathogen colonization in the ceca, liver, and oviduct. After 7 days of challenge (day 17), eggs were collected daily from each treatment group and tested for the presence or absence of SE until 66 days. In order to simulate a reinfection occurring in layer flock, the birds were reinoculated orally with SE (5×10 log10 CFU/mL per bird) on day 35. Additionally, cloacal swabs from all birds were analyzed weekly until 66 days for the presence or absence of SE. At the end of 66 days, the birds from all treatment groups were euthanized via CO2 asphyxiation, and cecum, oviduct, and liver samples from birds were collected in 10 mL of sterile PBS for SE detection.
SE determination on egg surface and egg contents
The presence of SE on eggshell surface and in egg contents was determined according to the method of Miyamoto et al. (1997). After 7 days of SE challenge, eggs from birds were collected daily and checked for the presence or absence of the pathogen until 66 days of infection. Each egg was rinsed separately in a sterile stomacher bag containing 50 mL of selenite cysteine broth supplemented with NA (50 μg/mL) for 2 min. After washing, the egg was removed and the broth from stomacher bag was incubated at 37°C for 48 h, followed by streaking on XLD+NA plates to detect SE on eggshell. The bacterial colonies were confirmed as Salmonella using the Salmonella rapid-detection kit (Microgen Bioproducts Ltd., Camberley, UK). The eggs that were washed in selenite cysteine broth were disinfected by wiping with 70% ethanol, dried, cracked open aseptically, and the shell and egg contents were collected into separate, stomacher bags containing 50 mL of selenite cysteine broth containing NA. The bags with the egg contents or shell were homogenized for 1 min in a stomacher, and incubated at 37°C for 24–48 h to detect Salmonella present inside the egg. The bacterial colonies were confirmed as SE as described previously.
SE determination in internal organs
The presence of SE in the oviduct, liver, and cecum were determined as described previously (Kollanoor-Johny et al., 2012). The organ samples and their contents from each bird were weighed and homogenized. Each homogenate was serially diluted (1:10) in PBS, and appropriate dilutions were plated on XLD-NA plates for bacterial enumeration. Representative colonies from XLD-NA plates were confirmed as Salmonella using the Salmonella rapid detection kit (Microgen Bioproducts Ltd.). When colonies were not detected by direct plating, samples were tested for surviving Salmonella by enrichment in 100 mL selenite cysteine broth (Oxoid Ltd., Basingstoke, UK) for 48 h at 37°C (Kollanoor-Johny et al., 2012), followed by streaking on XLD-NA plates. In addition, cecal endogenous bacteria were enumerated by plating appropriate dilutions of the ceca samples on duplicate thioglycollate agar plates (Difco, Becton Dickinson), followed by incubation at 39°C under 5% CO2 for 24 h.
Sensory evaluation of eggs
The sensory evaluation of eggs was conducted at the Sensory Laboratory, Department of Poultry Science, Auburn University, Alabama. Eggs were collected from unchallenged control and CA-treated birds once a week for 3 wks, and were tested using the triangle test (Roessler et al., 1978) to assess whether consumers can detect a difference between the eggs from CA treatments and control birds. Sensory testing was done with 36 panelists (students, staff, faculty, and local townsmen) per experiment and the experiment was repeated three times over a period of 3 wks. The panelists were randomly served with three coded scrambled egg samples for tasting and detection of organoleptic differences in a sensory booth under white light. The effect of residual taste in the mouth was minimized by using a distilled water (at room temperature) mouth rinse between each sampling.
Statistical analysis
The number of SE colonies in the organs was logarithmically transformed (log10 CFU/g) before analysis to achieve homogeneity of variance. These data were analyzed using the PROC-GLM procedure of the statistical analysis software (version 9.2, SAS Institute Inc., Cary, NC). Differences among the means were detected at p<0.05 using Fisher's least significant difference test for SE on eggshell and in yolk, whereas means were detected at p<0.05 in organs. For the sensory study, analysis of results was done for a probability level of 5% using a table of “Minimum Numbers of Correct Judgments” (Roessler et al., 1978).
Results
Effect of CA on SE contamination of eggshell and yolk
In-feed supplementation of CA at 0.7% or 1% did not significantly change (p>0.05) the body weight or the egg production in birds compared to controls (data not shown). However, CA supplementation (0.7% and 1%) decreased SE on shell and in the yolk (p<0.05). A total of 2543 eggs from the first experiment, and 2518 eggs from the second experiment, were tested over a period of 7 wks for the presence of Salmonella on the shell and in yolk. The average weekly results (N=5061) from both experiments are depicted in Figure 1A and B, wherein CA at 0.7% and 1% consistently decreased Salmonella both on the shell (Fig. 1A) and in yolk (Fig. 1B) from wk 1 to wk 7 of supplementation (p<0.05). A significant difference (p<0.05) between the controls and treated groups was observed from wk 1, wherein control birds yielded ≈72% of SE-contaminated shell, while birds fed with CA had only 60% (0.7% CA) and 42% (1% CA) of their shells positive for SE (Fig. 1A). A similar decrease in SE was observed in treated birds during wk 2 and wk 3 (Fig. 1A). Moreover, despite reinoculation of SE at the end of the 3rd wk, CA at both concentrations effectively reduced the pathogen on the shell and in the yolk throughout the rest of the experiment until wk 7 (Fig. 1A and B). Analysis of the cumulative data on SE prevalence from 5061 eggs over the 7-wk period revealed that dietary supplementation of CA at 1.0% decreased SE presence to ≈14% on the shell and ≈10% in yolk, when compared to controls, which yielded ≈60% positive samples on shell and ≈43% in yolk (Fig. 2).
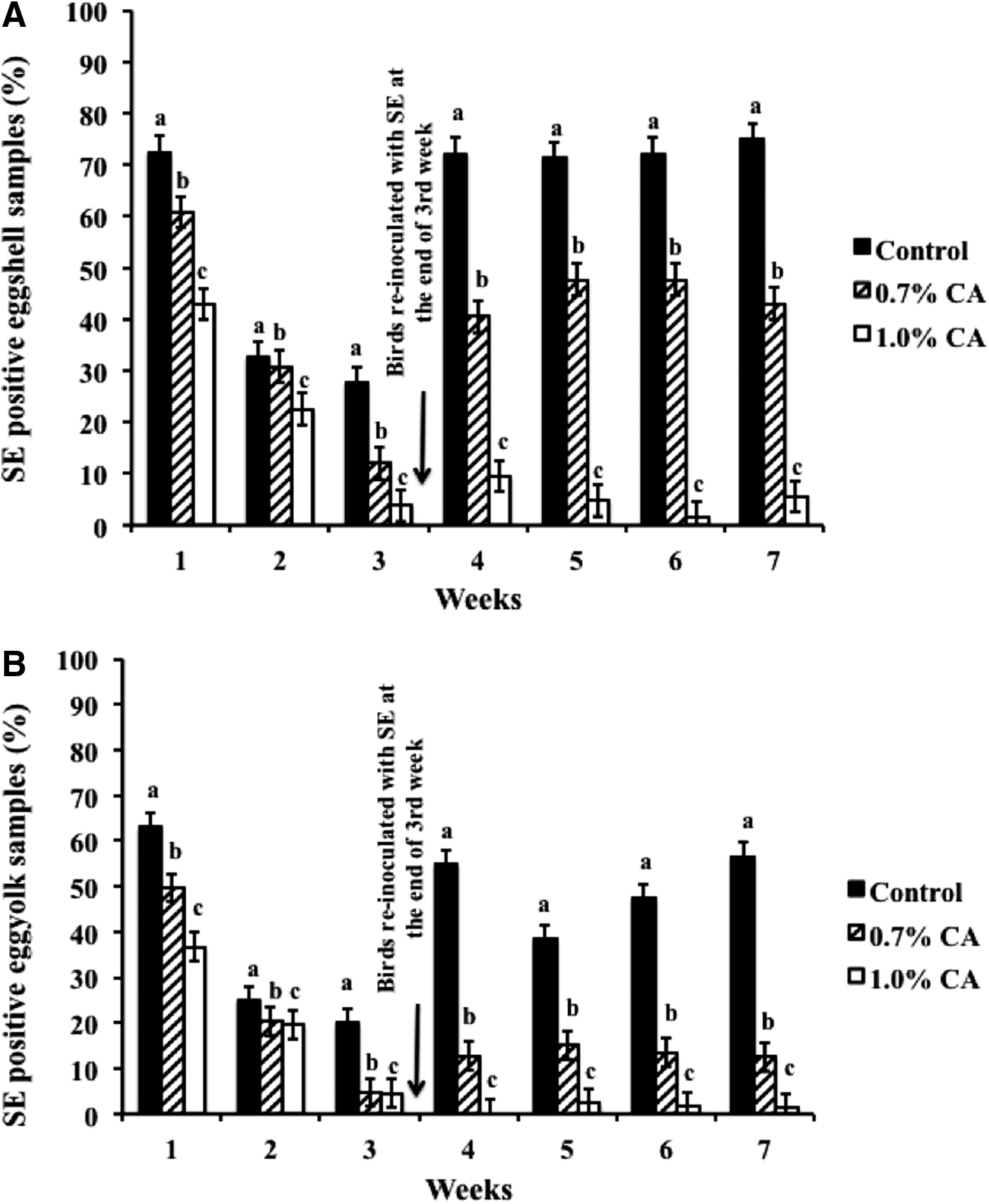
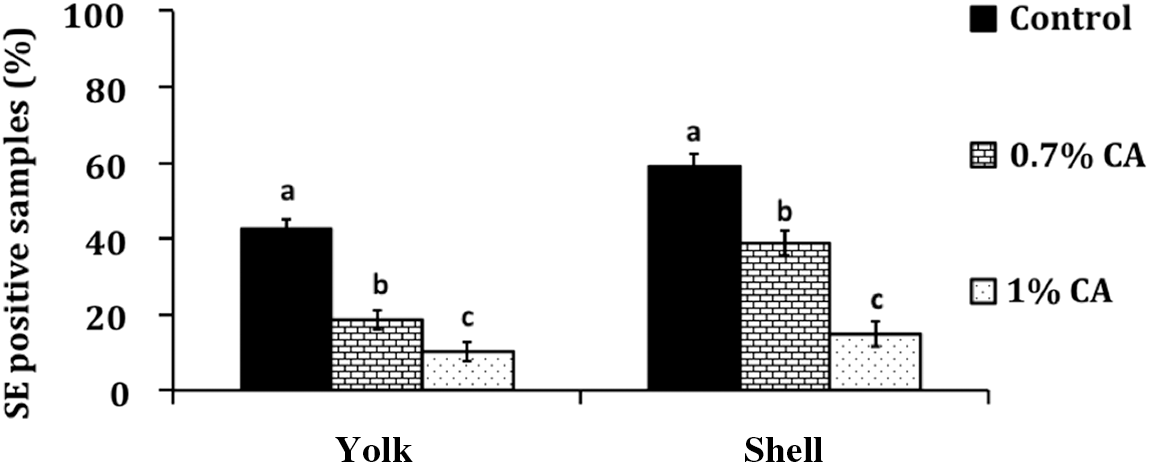
Cumulative effect of 0.7% and 1% caprylic acid (CA) treatment for 7 wks on eggshell, and in egg yolk, with N=5061 and p<l0.05. Bar graphs represents significant difference between treatment and control±standard error. SE, Salmonella enterica serovar Enteritidis.
Effect of CA on SE colonization of internal organs and cloaca
As observed in egg, CA supplementation significantly reduced SE colonization of the cecum and liver (p<0.05). At the end of 66 days, 70% of the cecal samples and liver samples from control birds tested positive for SE (Fig. 3A). Moreover, 40% of the oviduct samples from control birds were positive for the pathogen (Fig. 3A). However, feeding 1% CA-supplemented diet decreased SE in cecum and liver, with the pathogen recovered from only 25% of the cecum and 20% of liver samples from birds (p<0.05). Oviduct colonization of SE was reduced to 20% in birds supplemented with 1% CA; however, this difference was not statistically significant from control birds. In addition, the cecal endogenous bacterial counts did not differ (p>0.05) among birds from the various treatment groups (data not shown).
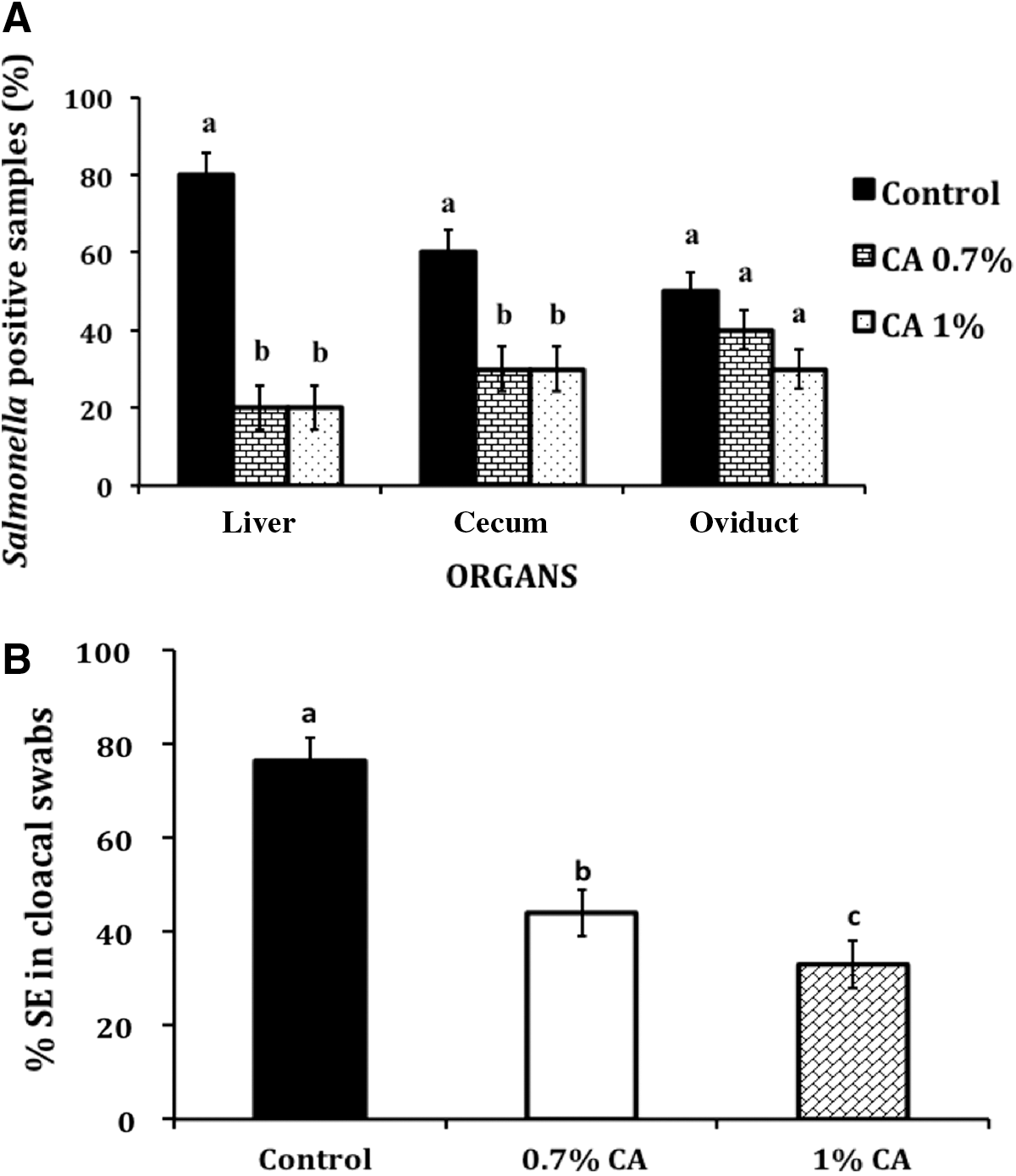
The presence of SE in cloaca was determined by testing cloacal swabs in all birds prior to CA supplementation, and this was followed by weekly collection for 7 wks in the inoculated group. The results showed that cloacal swabs collected before oral inoculation tested negative for SE (data not shown). Postinoculation, birds fed with CA consistently yielded a reduced number of SE-positive samples compared to control birds (p<0.05). The cumulative data at the end of the experiment revealed that 75% of the cloacal swabs from control birds were positive for SE (Fig. 3B), whereas 0.7% and 1% CA-supplemented birds yielded only 40% and 35% positive samples, respectively (p<0.05).
Effect of CA on sensory characteristics of eggs
When the eggs were subjected to consumer acceptance by triangle test, only 43 of the 111 panelists were able to detect the eggs from CA-treated birds, and the remaining 68 of them failed to identify the treatments from controls, thus denoting that the panelists were not able to detect a difference between the eggs from CA-supplemented and untreated birds (p>0.05).
Discussion
The cecal colonization of layers with SE results in the systemic spread of the pathogen to reproductive organs, thus contributing to direct transmission of the bacterium to eggs from infected ovaries and/or oviducts by the transovarian route. In addition, the pathogen can reach the spleen and liver by lymphatic or circulatory routes (Thiagarajan et al., 1994, 1996). Therefore, in the current study, we determined the efficacy of in-feed supplementation of CA in chickens in reducing SE in the aforementioned organs. Additionally, due to the fecal and systemic transmission of SE to the egg, CA's effect on decreasing the bacterium in cloacal contents, eggshell, and yolk was investigated.
The results from this study indicated that in-feed administration of CA significantly reduced SE colonization in layer chickens (p<0.05). Specifically, both concentrations of CA consistently decreased SE on eggshell and yolk from wk 1 to wk 7, with the higher concentration of 1% CA being more effective (p<0.05). Since reinfection with Salmonella can occur in a flock due to its persistence on a variety of sources in the farm environment (Hassan et al., 1991; Wilson, 2002), the birds were reinoculated with the bacterium at the end of 3 wks. However, despite the reinoculation, CA was effective in decreasing the pathogen prevalence on eggshell and yolk compared to control birds (Fig. 1A and B). Moreover, CA supplementation to birds reduced SE populations in the internal organs such as the cecum and liver (Fig. 3A), and in feces (Fig. 3B) (p<0.05).
The mechanism behind CA-mediated SE reduction in chickens could be multifold. Fatty acids, including CA, can diffuse into bacterial cells in their undissociated form and subsequently dissociate in the protoplasm, leading to acidification (Sun et al., 1998). Another potential bactericidal mechanism of CA involves inducing alterations in bacterial membrane permeability, thereby resulting in cell death (Bergsson et al., 1998). Additionally, the inhibitory effect of MCFAs on virulence-associated genes in SE has been suggested as a mode of antibacterial action. For example, Van Immerseel et al. (2004) observed that MCFAs suppressed the expression of hilA, a key gene regulator involved in Salmonella invasion, thereby resulting in decreased Salmonella colonization in chicks. Previous research from our laboratory also revealed that CA reduced SE invasion of avian intestinal epithelial cells by downregulating critical colonization-associated genes in the pathogen (Kollanoor-Johny et al., 2012).
Conclusions
The prophylactic supplementation of CA to chickens reduced SE contamination of egg yolk and shell without adversely affecting body weight, egg production, or consumer acceptability of eggs. These results suggest that CA could potentially be used as an antimicrobial feed additive to control eggborne transmission of SE when coupled with other standard biosecurity measures in the farm. Our future experiments will validate the findings of this study in commercial poultry farms.
Footnotes
Acknowledgments
This study was supported by a grant (2010-01346) from the U.S. Department of Agriculture National Integrated Food Safety Program.
Disclosure Statement
No competing financial interests exist.