
Abstract
Enterotoxigenic Escherichia coli (ETEC) is the major pathogen responsible for neonatal diarrhea, postweaning diarrhea, and edema disease in pigs. Although it can be harmless, ETEC is also present in the intestines of other animal species and humans, causing occasional diarrhea outbreaks. The evaluation of this pathogen's presence in food sources is becoming an increasingly important issue in human health. In order to determine the prevalence of ETEC in nondiarrheic pigs, 990 animals from 11 pig farms were sampled. Using end-time polymerase chain reaction (PCR), eltA, estI genes, or both, were detected in 150 (15.2%) animals. From the positive samples, 40 (26.6%) ETEC strains were isolated, showing 19 antibiotic-resistance patterns; 52.5% of these strains had multiple antibiotic resistances, and 17.5% carried the intI2 gene. The most prevalent genotypes were rfb O157 /estII/aidA (32.5%) and estI/estII (25.0%). The estII gene was identified most frequently (97.5%), followed by estI (37.5%), astA (20.0%), and eltA (12.5%). The genes coding the fimbriae F5, F6, and F18 were detected in three single isolates. The aidA gene was detected in 20 ETEC strains associated with the estII gene. Among the isolated ETEC strains, stx 2e/estI, stx 2e/estI/estII, and stx 2e/estI/estII/intI2 genotypes were identified. The ETEC belonged to 12 different serogroups; 37.5% of them belonged to serotype O157:H19. Isolates were grouped by enterobacterial repetitive intergenic consensus–PCR into 5 clusters with 100.0% similarity. In this study, we demonstrated that numerous ETEC genotypes cohabit and circulate in swine populations without clinical manifestation of neonatal diarrhea, postweaning diarrhea, or edema disease in different production stages. The information generated is important not only for diagnostic and epidemiological purposes, but also for understanding the dynamics and ecology of ETEC in pigs in different production stages that can be potentially transmitted to humans from food animals.
Introduction
E
ETEC-induced diarrhea holds negative economic implications for the pig industry due to the associated high mortality and reduced growth rate (Zhang et al., 2007). ETEC strains adhere to the small intestinal microvilli via fimbriae, such as F4, F5, F6, F18, and F41, encoded by faeG, fanC, fasA, fedA, and F41 genes, and produce enterotoxins that act locally on enterocytes. Based on their thermal stability, ETEC enterotoxins are classified as heat-labile toxins (LT-I and LT-II), encoded by eltA and eltB genes or heat-stable toxins (STa, STb and EAST1) encoded by estI, estII, and astA genes (Dubreuil, 2008). These toxins are the primary virulence traits responsible for diarrhea; however, the presence of virulence factors such as the EAST1 toxin (Vu Khac et al., 2006), the adhesin involved in diffuse adherence (AIDA-I), and the porcine attaching and effacing-associated factor (Paa) encoded by aidA and paa genes, have been implicated in porcine diarrhea (Ngeleka et al., 2003; Sherlock et al., 2004; Zhang et al., 2007). None of these potential virulence factors have been well characterized for their significance in porcine diarrhea, and studies of their association with other virulence factors are limited (Ngeleka et al., 2003; Chapman et al., 2006; Zhang et al., 2007).
Antimicrobial agents are widely used in swine production as therapeutic agents and growth promoters, and many of these antibiotics are important in human clinical medicine (Mathew et al., 2007). Recent studies have shown that ETEC serotypes have developed resistance to antimicrobial agents used in human and veterinary medicine (Smith et al., 2010; Wang et al., 2011). The dissemination and acquisition of such genes by horizontal gene transfer has led to the rapid emergence of antibiotic resistance among bacteria (Carattoli, 2013). Class 1 and 2 integrons are the most prevalent in isolates and are largely implicated in the dissemination of antibiotic resistance (de la Torre et al., 2014).
The purpose of this study was to investigate (1) the prevalence of ETEC-positive pigs in 11 farms without clinical signs of diarrhea, (2) the phenotypic and genotypic characterization of ETEC isolated from nondiarrheic pigs, and (3) antibiotic resistance among isolated ETEC strains.
Materials and Methods
Study design and sample collection
A cross-sectional study was carried out on 11 farms located in Buenos Aires, Argentina, with no history or clinical signs of colibacillosis. Swine units were included by 1 of the following criteria: sow herd size (225±30) and finisher pigs delivered to slaughter/year (at least 2500 finisher pigs to market/year). The selected farms have the same management style and are farrow-to-finish. The geographic distribution represents the most swine-dense areas of Buenos Aires. Ninety rectal swabs were collected from each farm to determine the presence of eltA, estI, or both, and the subsequent prevalence of different genotypes in each production stage. Pigs considered being ETEC positive (ETEC+) were those in which eltA, estI, or both were detected. The DNA template was prepared from the confluent growth zone. Thirty randomly selected pigs from each production stage were sampled as follows: farrowing (weaning piglet, 21±3 days old); nursery (nursery piglets, 86±3 days old); and finishing (finishing pigs, 165±3 days old). Samples were collected over a period of 16 weeks. Samples consisted of rectal swabs (EUROTUBO® Collection Swab; DELTALAB, Barcelona, Spain), which were stored at 4°C and processed within 24 h.
Extraction of bacterial DNA and screening of eltA and estI genes from nondiarrheic pigs
A total of 990 rectal swabs were tested for the presence of ETEC genes by PCR detection of eltA and estI. Rectal swabs were streaked onto MacConkey Agar plates (Britania, Buenos Aires, Argentina) and incubated at 37°C for 18 h. Each sample was homogenized by inoculation of a loopful of the confluent growth zone in 3 mL of tryptic soy broth (TSB) (Britania). From each bacterial suspension, 700 μL was frozen at −70°C in 30.0% glycerol. DNA extraction was performed using 150 μL of bacterial suspension, as previously described (Leotta et al., 2005). The PCR protocols have been previously described (Toma et al., 2003). The strains E. coli ATCC 33965 (eltA/estI) and E. coli ATCC 25922 were used as positive and negative controls, respectively.
Positive ETEC isolation, biochemical characteristics, and hemolysin activity
Frozen stocks of samples positive for the eltA, estI, or both were inoculated in TSB and incubated at 42°C for 24 h. Isolates of positive samples were made by streaking the enrichment culture onto one MacConkey Agar plate (Britania) and three Eosin Methylene Blue (EMB) Agar plates (Britania). A total of 60 individual colonies per sample, with E. coli morphology, were tested for the presence of virulence genes. One positive isolate per sample for eltA, estI, or both was confirmed as E. coli by standard biochemical tests (Brusa et al., 2013).
Conventional serotyping
The presence of O and H antigens was determined by a previously described method (Guinée et al., 1981) with slight modifications (Blanco et al., 1997) in which all available O (O1-O181) and H (H1-H56) antisera were used (Orskov et al., 1984). Nonspecific agglutinins were removed by adsorption of each antiserum with the corresponding cross-reacting antigens. The O and H antisera were produced in the Laboratorio de Referencia de E. coli (LREC), Universidade de Santiago de Compostela (Lugo, Spain).
Antimicrobial susceptibility testing
Antimicrobial susceptibility was tested using the disk-diffusion method following the Clinical and Laboratory Standards Institute standard M31-A3 and M100-S23 (CLSI, 2008, 2013), for the following antimicrobial agents: ampicillin (AMP), cephalothin (CET), cefotaxime (CTX), cefoxitin (CXT), amoxicillin/clavulanic acid (AMC), gentamicin (GEN), amikacin (AMK), streptomycin (STR), tetracycline (TET), nalidixic acid (NAL), ciprofloxacin (CIP), chloramphenicol (CHL), florfenicol (FFN), trimethoprim-sulfamethoxazole (SXT), nitrofurantoin (NIT), fosfomycin (FOF), and colistin (CST) disk (Britania). The manufacture guidelines were used for CST interpretation. E. coli ATCC 25922 was used as reference strain. Multiple antibiotic resistance (MAR) was defined as ETEC isolates showing three or more antimicrobial classes resistance (Schwarz et al., 2010).
Genotypic characterization
According to references included in Table 1, PCR testing was performed to detect the presence of genes coding for LT, STa, STb, Stx1, Stx2, Stx2e, and EAST1 toxins, as well as Class 1 and Class 2 integrons (intI1 and intI2), and F4, F5, F6, F18, F41, intimin, Paa and AIDA-I adhesins, and the rfb O157 gene. Reference strains E. coli EDL933 (eae/paa/rfb O157), E. coli 7805 (eltA/estI/estII/faeG/east1), E. coli 81-603 A (fasA), E. coli 1073 B44 (fanC/F41), and E. coli 88-1199 (fedA) were used as positive controls. ETEC isolates were further characterized by enterobacterial repetitive intergenic consensus (ERIC)–PCR (Versalovic et al., 1991). ERIC fingerprints of amplified DNA fragments were obtained by agarose gel electrophoresis and analyzed using the software BioNumerics Version 6.6 (Applied Maths, Sint-Martens-Latem, Belgium).
Statistical analysis
Within-herd prevalence for each genotype in different production stages was calculated by dividing the number of ETEC-positive samples by each genotype identified. Ninety-five percent confidence intervals were computed for all of these estimates.
Results
Prevalence of eltA and estI genes in nondiarrheic pigs
Of the 990 samples screened, 150 (15.2%; 95.0% CI±2.24) samples were positive for the presence of eltA, estI, or both. Of these ETEC+ pigs, 82.7% (95.0% CI±6.05) were positive for estI, 11.3% (95.0% CI±5.07) were positive for eltA, and 6.0% (95.0% CI±3.8) were positive for estI/eltA (Fig. 1). The overall percentage of ETEC+ animals increased from 16.6% (95.0% CI±5.95) in the farrowing phase to 66.0% (95.0% CI±7.58) in the nursery phase. The percentage of carrier pigs abruptly declined to 17.3% (95.0% CI±6.05) in the finisher population. ETEC+ pigs were present in 10 of the 11 evaluated farms. Among the 150 ETEC+ pigs, 40 ETEC strains (26.7%) (95.0% CI±13.71) were isolated. ETEC strains were isolated from 30 nursery samples (75.0%) (95.0% CI±13.42), 8 finishing samples (20.0%) (95.0% CI±12.4), and 2 farrowing samples (5.0%) (95.0% CI±6.75).
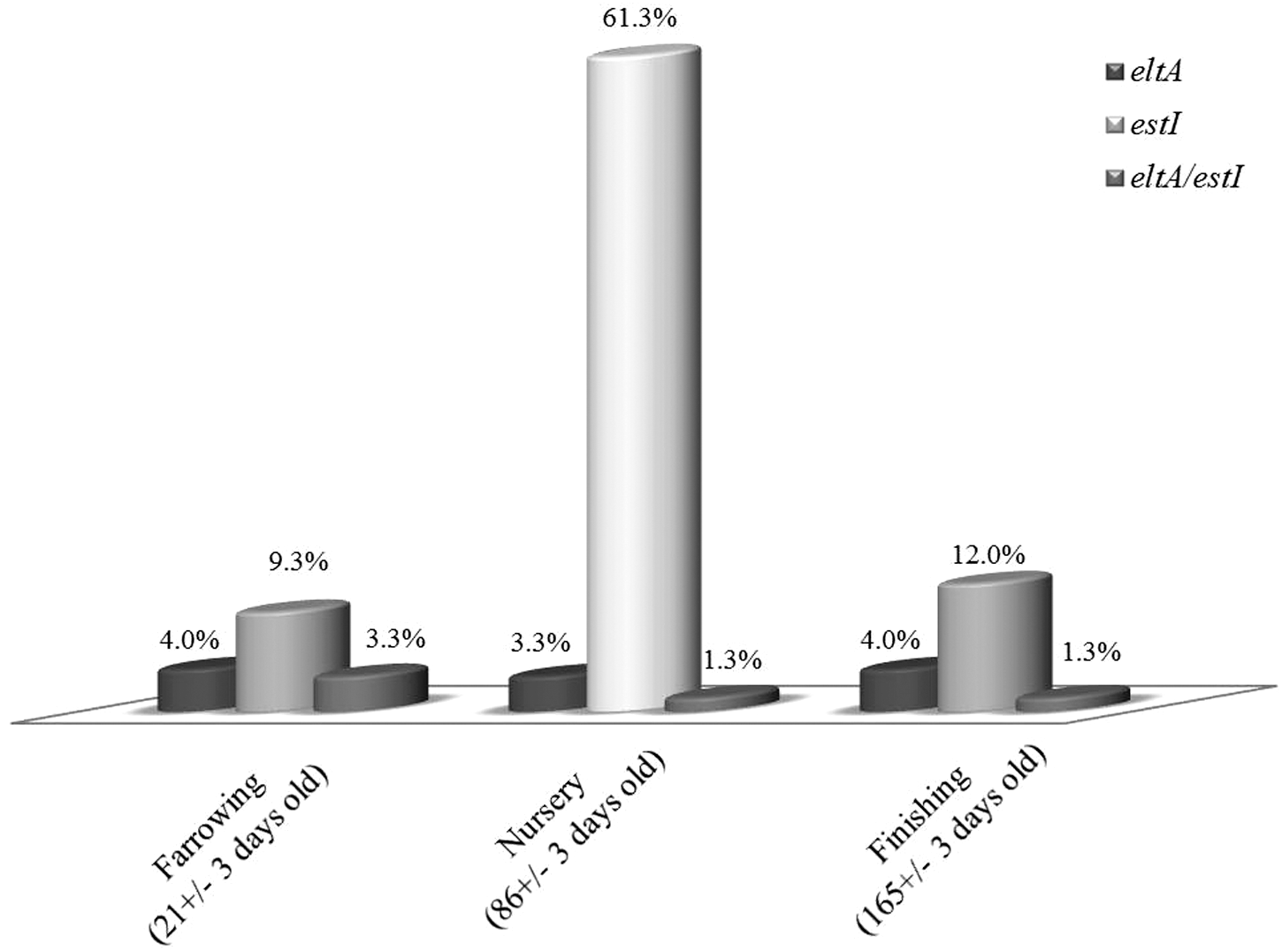
Prevalence of eltA (toxin LT) and estI (toxin STa) genes obtained from the confluent growth zone in tryptic soy broth (TSB). A total of 90 rectal swabs were collected from each farm to determine the presence of eltA (toxin LT) and estI (toxin STa) genes and the subsequent prevalence of different genotypes in each production stage. Thirty randomly selected pigs, from three different stratums based upon the age and stage of production, were sampled.
Serotype profiles of the ETEC strains
The ETEC strains were grouped in 12 different serotypes, with O157:H19 being the most prevalent (n=15; 37.5%), followed by serotypes O8:H- (n=7; 17.5%), ONT:H- (n=5; 12.5%), O8:H31 (n=4; 10.0%), O8:H26 (n=2; 5.0%), and individual cases of serotypes O7:H-, O7:H15, O7:H16, O54:H21, O60:H-, O149:H-, and O157:H-. Of the nontypeable somatic strains, all were nonmotile (Fig. 2).
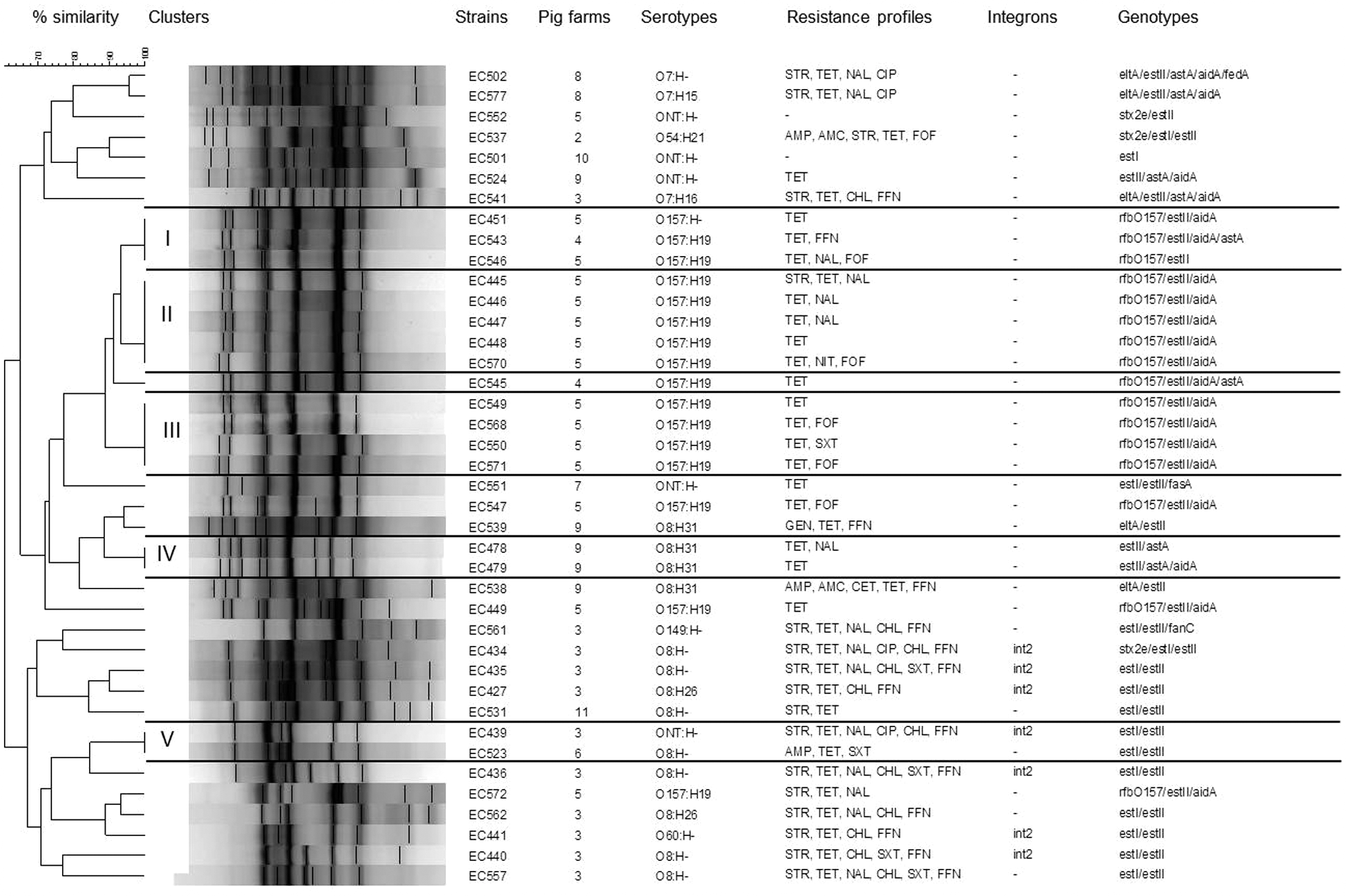
Enterobacterial repetitive intergenic consensus–polymerase chain reaction (ERIC-PCR) dendrogram showing the genetic relationship, serotypes, antimicrobial resistance profiles (for antimicrobial abbreviations, see text), and virulence genes of Enterotoxigenic Escherichia coli (ETEC) isolated from nondiarrheic pigs. The dendrogram, generated by BioNumerics software (Applied Maths, Belgium), shows distances calculated by the Dice similarity index of ERIC-PCR among 40 ETEC strains. The degree of similarity (%) is shown on the scale. Roman numbers indicate cluster number. The dendrogram was generated based on the unweighted-pair group method with the arithmetic mean.
Antimicrobial susceptibility of ETEC strains
Two of the 40 ETEC strains were susceptible to all antimicrobial agents tested. All of the strains were susceptible to AMK, CST, CTX, and CXT. Thirty-eight (95%) strains were resistant to at least 1 antimicrobial tested, and all of these strains showed resistance to TET. Nineteen different antimicrobial resistance profiles were observed. MAR to more than 3 antibiotics was observed in 52.5% of the strains (Table 2).
Antimicrobial: ampicillin (AMP), cephalothin (CET), amoxicillin/clavulanic acid (AMC), gentamicin (GEN), streptomycin (STR), tetracycline (TET), nalidixic acid (NAL), ciprofloxacin (CIP), chloramphenicol (CHL), florfenicol (FFN), trimethoprim-sulfamethoxazole (SXT), nitrofurantoin (NIT), fosfomycin (FOF).
Multiple antimicrobial resistances.
Genotypic characterization and molecular subtyping of ETEC isolates
We observed 16 different virulence profiles among the 40 isolated ETEC strains (Table 3). ERIC-PCR analysis was able to subtype the 40 ETEC strains of different serotypes, generating 29 distinct ERIC-PCR patterns with 6–12 discernible fragments, ranging from 300 to 1500 bp. Sixteen isolates were grouped in 5 clusters (I–V), and 24 isolates presented unique ERIC-PCR patterns (Fig. 2). Cluster I included three ETEC strains, two of them belonging to serotype O157:H19 and one to serotype O157:H-. The Cluster I strains presented three different antimicrobial resistance patterns (TET, TET/FFN, and TET/NAL/FOF) and had three different genotypes (rfb O157/estII/aidA, rfb O157/estII/aidA/astA, and rfb O157/estII). Cluster II grouped five O157:H19 strains that had four different antimicrobial resistance patterns (TET, TET/NAL, STR/TET/NAL, and TET/NIT/FOF) and one genotypic profile (rfb O157/estII/aidA). Cluster III grouped four O157:H19 strains that had three different antimicrobial resistance profiles (TET, TET/FOF, and TET/SXT) and one genotype (rfb O157/estII/aidA). Of the 12 strains grouped within Clusters I, II, and III, 11 strains were isolated from Farm 5 and 1 from Farm 4. Cluster IV included two O8:H31 strains from Farm 9 that had two antimicrobial resistance patterns (TET and TET/NAL) and presented two different genotypic patterns (estII/astA and estII/astA/aidA). Cluster V included two strains isolated from Farms 3 and 6, O8:H- and ONT:H-. These strains had two antimicrobial resistance patterns (AMP/TET/SXT and STR/TET/NAL/CIP/CHL/FFN) and two genotypic profiles (estI/estII/intI2 and estI/estII).
ETEC/Shiga toxin–producing Escherichia coli hybrids* (n): number of strain.
Discussion
In pigs, ETEC is normally associated with ND and PWD; however, this pathogen can also be shed in feces from healthy animals (Osek, 1999). This study revealed the presence of ETEC in nondiarrheic pigs in different production stages. The overall prevalence of animals carrying eltA, estI, or both was 15.2%, which appears to be lower than that reported in animals with clinical diarrhea (Vidotto et al., 2009; Zajacova et al., 2012). In this study, the percentage of ETEC+ nondiarrheic piglets during the lactation period was 16.6%. Reported prevalence values in clinically affected neonates are higher, varying from 25.7% to 43.0% (Nakazawa et al., 1987; Do et al., 2006). However, the prevalence of ETEC+ nursery pigs observed during this study does not differ from other reports involving animals with clinical diarrhea (Fairbrother et al., 2005; Do et al., 2006). The prevalence of ETEC+ pigs observed during the finisher period is consistent with the prevalence of ETEC previously reported among animals at slaughter (Martins et al., 2010). ETEC clinical manifestation is a complex process that not only requires the presence of ETEC strains (Dewey et al., 1995) but also environmental changes, considered risk factors for the presence of clinical disease (Amezcua et al., 2002; Laine et al., 2008). Therefore, detection of subclinical carriers should be considered in order to prevent clinical manifestation and further dissemination of ETEC strains.
ETEC-STa strains are believed to be responsible for most cases of diarrhea in animals less than a week old; in animals more than 4 weeks old, diarrhea is more likely the result of ETEC-STb+ or ETEC-STb/a+ (Moon et al., 1986). Although the estII gene was not evaluated in this study's screening stage, the genotypic characterization of ETEC strains showed estII's prevalence at 39/40 (97.5%). Several studies have demonstrated that bacteria encoding estII, either alone or in combination with other toxins, are highly prevalent in animals with clinical diarrhea (Dubreuil, 1997). In this study, the most frequent gene combination was rfb O157/estII/aidA (32.5%). Previous reports proposed that ETEC-STb strains might lack an adherence factor, or there might be dysregulation of the amount of toxin produced. ETEC-STb is also commonly associated with the presence of F4 fimbria (Dubreuil 1997). In this study, the estII gene was present in ETEC isolates carrying F5, F6, and F18 genes, but not F4. Although F5 and F6 are commonly associated with ND, and F18 with PWD, we observed strains carrying genes encoding estII and fimbrial adhesins F5, F6, and F18 in nondiarrheic pigs. In this study, 8 strains (20.0%) harboring the astA gene were observed in association with ETEC-F6+ and ETEC-F18+ strains. The role of the EAST1 toxin in swine colibacillosis has not been fully demonstrated. However, the astA gene encoding the EAST1 toxin is commonly found in ETEC isolates associated with PWD (Vu Khac et al., 2006; Moredo et al., 2012).
Two additional virulence factors, AIDA-I and Stx2e, are present in ETEC strains associated with diseased pigs (Ngeleka et al., 2003; Wang et al., 2011). The AIDA-I gene seems to be equally prevalent in fimbrial and nonfimbrial isolates (Zhang et al., 2007); in this study, 50.0% of isolates were ETEC-aidA +; however, only a single isolate (eltA/estII/astA/aidA/fedA) was associated with the presence of an F18 gene. Previous reports suggested that the STb/AIDA association could be the missing link in what we know about swine colibacillosis; the similarities between AIDA-I of human origin and that of porcine origin could represent a potential danger of cross-infection between humans and pigs (Dubreuil, 2010). We observed three ETEC strains carrying stx 2e/estI and stx 2e/estI/estII genes. Our findings concur with recent studies that have observed an ETEC strain carrying stx 2 genes (Wang et al., 2011). These strains carrying ETEC and Shiga toxin–producing Escherichia coli (STEC) genes have been proposed as ETEC/STEC hybrids carrying stx 2 gens (Tozzoli et al., 2014). The importance of these findings is due to the potential risk of human infection with ETEC/STEC hybrids (Wester et al., 2013).
Previous studies have observed, in varying frequency, the following serogroups associated with PWD and ED: O8, O45, O138, O139, O141, O149, and O157 (Frydendahl, 2002; Fairbrother et al., 2005). Fairbrother et al. (2005) have described serogroups O149 and O157 as the most frequent cause of ND, PWD, and ED. In the present study, serotypes O8:[H21,H26,H31], O149:H-, and O157:H19 were the most prevalent among ETEC isolates in nondiarrheic pigs. The serogroups detected in this study concur with those reported in the literature as causative agents of disease in pigs (Frydendahl, 2002; Fairbrother et al., 2005). Serotypes O157:H7 and O8:[H2,H8,H9,H19,H-] have been identified in human outbreak of STEC (Friesema et al., 2015; Kaper and O'Brien, 2014). Although only Shiga toxin–negative strains of E. coli O157 has been proved to be capable of acquiring stx genes (Wetzel and LeJeune, 2007), the potential role of Shiga toxin–negative strains of E. coli O8 cannot be ruled out, increasing their potential to emerge as new Shiga toxin–producing E. coli strains.
Of the 40 ETEC strains, 16 (40.0%) were characterized as O157:[H19/H-]. According to the subtyping analysis performed in this study, ETEC O157:[H19/H-] strains were grouped in three different clusters (I, II, and II); however, these strains differed by antimicrobial resistance patterns and genotypic profile. These results are consistent with previous reports in which ERIC-PCR had a low discriminatory rate among strains of E. coli O157:H7 (Giammanco et al., 2002). However, we observed that ERIC-PCR has better discriminatory power among ETEC O8:[H26/H31/H-] strains, the second most prevalent serotype in our study. These isolates were grouped into two clusters, and the technique was capable of differentiating nine strains with individual ERIC-PCR patterns.
Previous publications demonstrated that E. coli strains responsible for PWD could be resistant to multiple antibiotics (Fairbrother et al., 2005; Wang et al., 2011). In our study, 52.5% of the strains showed different MAR patterns. Prophylactic uses of antibiotics seem to play a role in MAR. A small number of studies have demonstrated that certain groups of drugs can have a prophylactic effect; however, when these antibiotics were withdrawn, clinical cases of PWD increased (Casewell et al., 2003). In this study, 7 (17.5%) of the ETEC strains were positive for intl2 and resistant to STR, TET, CHL, and FFN. Previous studies have shown an association between MAR strains, including STR and TET in their multiresistance profiles, and the presence of Class 2 integrons detected in E. coli strains isolated from swine (Kadlec and Schwarz, 2008; Lapierre et al., 2008). The presence of MARs ETEC strains suggest that there is a great need for surveillance programs to monitor MARs E. coli that can be potentially transmitted to humans from food animals.
This study demonstrates that numerous potentially virulent ETEC genotypes cohabit and circulate in swine populations without clinical manifestation of ND or PWD. The information generated is important not only for diagnostic and epidemiological purposes, but also for understanding the dynamics and ecology of ETEC in pigs in different production stages that can be potentially transmitted to humans from food animals.
Footnotes
Acknowledgments
We thank Dr. Kent J. Schwartz (Iowa State University Veterinary Diagnostic Laboratory) and Dr. Roberta Alvarez for their critical revisions of this article.
Disclosure Statement
No competing financial interests exist.