
Abstract
The aim of the study was to survey three foodborne pathogens in kiwifruit orchards as a continuous monitoring program. A total of 193 samples were collected from 11 kiwifruit orchards in Shaanxi province in October 2013. Among the 193 samples, 68 Escherichia coli isolates were recovered, while no Staphylococcus aureus and Salmonella was recovered. All E. coli isolates were characterized by antimicrobial susceptibility testing, detection of virulence genes, and the ability to produce biofilm formation. The isolates were further examined by random amplified polymorphic DNA (RAPD) analysis. E. coli isolates displayed resistance most frequently to tetracycline (48.5%). Two E. coli isolates (2.9%) were positive for the eae gene (the intimin gene). All E. coli isolates lacked the ability to make biofilm formation. Multilocus sequence typing analysis demonstrated that one isolate in kiwifruit orchards shared the same sequence type with a human clinical isolate. RAPD results showed a close relationship among E. coli isolates from fresh fruit, fallen fruit, soil, air, and irrigation water. This study could provide a further understanding of microbial contamination in kiwifruit orchards based on our previous study and help growers take appropriate measures for prevention.
Introduction
T
Pathogenic Escherichia coli, Salmonella, and Staphylococcus aureus are three of the most common foodborne pathogens that contaminate fresh food, which can result in foodborne illness (Mansur and Oh, 2015). In May 2011, a large outbreak of the hemolytic-uremic syndrome caused by Shiga toxin–producing E. coli O104:H4, which occurred in Germany, was associated with sprouts (Buchholz et al., 2011). There was a total of 33,353 illnesses in South Korea during 2001–2005, among which 10.8% became sick due to enterotoxins produced by S. aureus (Yoon et al., 2011). In 2008, a Salmonella Saintpaul outbreak in the United States implicated jalapeño pepper and Serrano pepper, which highlighted the importance of preventing contamination in the field (Barton Behravesh et al., 2011). The consumption of fruit continues to rise all around the world due to healthy-lifestyle recommendations. Meanwhile, the rate of foodborne illness caused by the consumption of fresh fruit remains high in the United States and European Union, representing a significant public health and financial issue (Callejón et al., 2015).
Kiwifruit, Actinidia chinensis, is a nutritious fruit mainly grown in China, New Zealand, and Italy. Since China is the biggest kiwifruit exporter in the world, ensuring safe kiwifruit consumption in China is important both domestically and internationally (Testolin and Ferguson, 2009). Fresh produce has been shown to harbor various levels of bacterial loads (aerobic bacteria, coliform bacteria, and yeast and molds), ranging from <1 to 8 log10 colony-forming units (CFU) per gram, with most samples ranging between 3 and 4 log10 CFU per gram (Badosa et al., 2008). Regular monitoring of microbial hygiene status in kiwifruit orchards helps growers take approaches to prevent preharvest contamination. This study followed the previous survey carried out in 2012 (Feng et al., 2014) to continue the rolling program of monitoring E. coli, S. aureus, and Salmonella contamination of kiwifruit orchards. The study aimed to understand the microbial status in kiwifruit orchards in 2013 and see whether any difference of contamination rates existed in different years.
Materials and Methods
Experimental design and sample collection
Eleven commercial orchards in the main kiwifruit production region were selected in two cities (Zhouzhi [n = 6], Meixian [n = 5]) in Shaanxi province just before harvest. The region is near east longitude 108°, north latitude 34°, and 500 m above sea level. Orchards were sampled once in October 2013.
Gloves and boot covers were changed between each orchard, and gloves were disinfected with 75% ethanol prior to sample collection in order to avoid cross-contamination during the sampling. A total of 193 various types of samples were collected. The samples in kiwifruit orchards included kiwifruits on trees (n = 54) and fallen kiwifruits (n = 48) on the ground, soil (n = 55), irrigation water (n = 3), and air (n = 33). The air samples were collected by exposing the Luria Bertani agar (LB agar; Beijing Land Bridge Technology Ltd., Beijing, China) plates for 5 min in the air. After sampling, about 25 g of each soil sample were weighed, and 25 mL of each water sample was used for isolation. Each whole fruit was used for isolation directly without weighing.
Escherichia coli isolation and identification
Isolation of E. coli was performed according to procedures described previously (Feng et al., 2014). Each sample (25 g or mL) or the whole fruit was homogenized in 225 mL of buffered peptone water (BPW; Beijing Land Bridge Technology Ltd.) and homogenized for 2 min at 320 rpm. The fruit was placed into 225 mL BPW without removing the skin. The plate, exposed to air, was rinsed with 25 mL of BPW. After incubation for 10 min at room temperature with periodic shaking, the culture was transferred into a sterile container. After incubating for 3 h at 35°C, the culture was transferred to 225 mL of double-strength tryptone phosphate broth (TP broth; Oxoid, Basingstoke, England). After incubation at 44°C for 20 h, a loopful of the culture was streaked onto eosin–methylene blue agar (EMB; Beijing Land Bridge Technology Ltd.) and MacConkey agar (Beijing Land Bridge Technology Ltd.). After incubation for 20 h at 35°C, the plates were examined for morphologically typical (with metallic green sheen on EMB plates and pink on MacConkey plates) E. coli colonies. For each sample, one presumptive E. coli isolate was picked and confirmed by polymerase chain reaction (PCR) detection of the uidA gene (Feng et al., 2014). These E. coli isolates were stored at −80°C in Luria Bertani (LB) broth (Beijing Land Bridge Technology Ltd.) containing 30% glycerol for further analysis.
Staphylococcus aureus isolation and identification
Isolation of Staphylococcus aureus was performed according to procedures described previously (Feng et al., 2014). Each sample (25 g or mL) was placed in 225 mL of BPW and incubated at 37°C for 24 h. After pre-enrichment, 5 mL of the culture was transferred to 50 mL of trypticase soy broth (Beijing Land Bridge Technology Ltd.) containing 7.5% NaCl. After 18–24 h incubation at 35°C, a loopful of the culture was inoculated onto Baird-Parker agar (BPA, Beijing Land Bridge Technology Ltd.) plates with 5% egg yolk and tellurite. After incubation for 20 h at 35°C, plates were examined for morphologically typical (black colonies surrounded by a pale halo) S. aureus colonies. For each positive sample, one presumptive S. aureus isolate was picked and confirmed by PCR detection of the thermonuclease gene (Brakstad et al., 1992).
Salmonella isolation and identification
Isolation of Salmonella was performed according to procedures described in the National Standards of the People's Republic of China (GB 4789.1-2010). Briefly, each sample (25 g or mL) was placed in 225 mL of BPW and incubated at 37°C for 8–18 h. Ten mL aliquots of pre-enriched cultures were transferred to 100 mL each of tetrathionate broth base (TT, Beijing Land Bridge Technology Ltd.) and selenite cystine broth (SC, Beijing Land Bridge Technology Ltd.), respectively. After incubation at 42°C for 18–24 h in TT broth and in SC broth, one loopful of each broth was streaked onto bismuth sulfite (BS, Beijing Land Bridge Technology Ltd.) and xylose lysine desoxycholate agar (XLD, Beijing Land Bridge Technology Ltd.). The BS plates were incubated at 37°C for 40–48 h and XLD plates were incubated at 37°C for 18–24 h. After incubation, plates were examined for morphologically typical Salmonella colonies. For positive samples, one presumptive Salmonella isolate was picked and confirmed by PCR detection of the invA gene (Yang et al., 2010).
PCR detection of virulence genes
Six virulence genes specific for each type of diarrheagenic E. coli were tested by PCR: the eae gene (Oswald et al., 2000) for enteropathogenic E. coli (EPEC), the ipaH gene (Sethabutr et al., 1993) for enteroinvasive E. coli, the elt gene (Tamanai-Shacoori et al., 1994), and the est gene (Hornes et al., 1991) for enterotoxigenic E. coli, the aggR gene (Ratchtrachenchai et al., 2012) for enteroaggregative E. coli, and the stx-genes stx1/stx2 (Yamasaki et al., 1996) for Shiga toxin–producing E. coli (STEC). The PCR products were resolved by 1.0% (wt/vol) agarose gel electrophoresis in 0.5 × Tris-borate EDTA (TBE) buffer.
Serotyping
E. coli isolates positive for virulence genes were serotyped in Henan Center for Disease Control and Prevention, China. O and H antigens were characterized using slide agglutination with hyperimmune sera (S&A Company, Bangna, Thailand) and the serotype was assigned following the manufacturer's instructions.
Antimicrobial susceptibility testing
Antimicrobial susceptibility test of E. coli isolates were performed by the agar dilution method for ciprofloxacin, nalidixic acid, gatifloxacin, ampicillin, gentamicin, kanamycin, amikacin, cefoxitin, ceftriaxone, trimethoprim/sulfamethoxazole, chloramphenicol, and tetracycline. E. coli ATCC 25922 and Enterococcus faecalis ATCC 29212 were used as quality control strains in each run. The breakpoints used were taken according to guidelines developed by the Clinical Laboratory Standard Institute (CLSI, 2013).
Biofilm formation
All E. coli isolates were tested for biofilm formation by using the Congo red agar method (Kaiser et al., 2013) with modification. In brief, isolates were cultivated on Brain Heart Infusion Medium (Beijing Land Bridge Technology Ltd.) with 1.5% agar, 0.08% (wt/vol) Congo Red (Sanland-Chem International Inc., Xiamen, China), 1.5% of NaCl and 2% (wt/vol) of glucose per liter. The plates were incubated at 37°C for 24 h. The biofilm producer strains were expected to form black colonies, while the non–biofilm producer strains formed red colonies. The results were checked by two observers, and experiments with the Congo red agar method were performed at least twice.
Random amplified polymorphic DNA (RAPD) analysis
Random amplified polymorphic DNA (RAPD) analysis was performed as described by Pacheco et al. (1997). Bacteria were grown overnight at 37°C in LB broth. An aliquot of the culture was diluted 1:10 in water immediately before use to determine the optical density at 600 nm. After being adjusted to an optical density value of 0.4, a 200-μL aliquot of the stationary-phase culture was centrifuged (12,000 × g for 2 min) and re-suspended in 900 mL of sterile distilled water. The suspension was then boiled for 10 min, briefly centrifuged, and used as the template (Schroeder, 2003). Three primers were selected: 1252 (5'-GCGGAAATAG-3'), 1254 (5'-CCGCAGCCAA-3'), and 1290 (5'-GTGGATCGA-3'). The PCR programs were as follows: 4 cycles of 94°C for 5 min, annealing temperature for 5 min, and 72°C for 5 min, followed by 30 cycles of 94°C for 1 min, annealing temperature for 1 min, and 72°C for 2 min, and a final extension step at 72°C for 10 min. The optimized annealing temperatures of the three primers were 36°C, 37°C, and 38°C, respectively. Each product was analyzed by electrophoresis on 1.5% agarose gels. Gels were photographed under ultraviolet light after being stained with ethidium bromide. The band patterns were characterized visually. The same reaction was performed three times to confirm each band in a certain position. The bands were identified as long as two repetitions were shown. At the same time, ATCC 25922 was used as control strain. The bands were coded as binary data (absent = 0 or present = 1). For each tested strain, the amplification profiles obtained with the three primers were combined for comparison. Data recording and calculations were performed by using NTsys (version 2.10e; Exeter Software, New York, NY). The resulting matrix of pairwise distances was used to generate a phenogram based on the unweighted pairwise distance with arithmetic mean (UPGMA) method, included in the NTsys software.
Multilocus sequence typing (MLST)
MLST was performed by using seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) (Wirth et al., 2006). Alleles and sequence types (STs) were determined following the instructions given elsewhere (Wirth et al., 2006). Then the STs of the selected strains were compared with the MLST database of E. coli provided by the website.
Results
Isolation of E. coli, S. aureus, and Salmonella
Of the 193 samples, 68 (35.2%) samples were positive for E. coli (Table 1), including 7 (13.0%) of 54 fresh fruit samples, 28 (58.3%) of 48 fallen fruits samples, 28 (50.9%) of 55 soil samples, 3 (9.1%) of 33 air samples, and 2 (66.7%) of 3 samples from irrigation water in the kiwifruit orchards. S. aureus and Salmonella were not detected in the present study.
PCR detection of virulence genes
Among 68 E. coli isolates, 2 isolates, recovered from fallen fruits in the orchards in Zhouzhi and Meixian, were positive for the eae gene. No other virulence gene specific for diarrheagenic E. coli was detected among the isolates.
Serotyping
The serotypes of two E. coli isolates containing the eae gene were O169: MH, and O158: H21.
Antimicrobial susceptibility testing
Of the 68 E. coli isolates, resistance was most frequently observed to tetracycline (48.5%), followed by ampicillin (30.9%), nalidixic acid (22.1%), ciprofloxacin (22.1%), cefoxitin (13.2%), gentamicin (10.3%), chloramphenicol (10.3%), gatifloxacin (8.8%), ceftriaxone (7.4%), and kanamycin (1.5%). All E. coli isolates were susceptible to trimethoprim–sulfamethoxazole and amikacin (Table 2). Forty E. coli (58.8%) isolates were resistant to at least one antimicrobial, 18 (26.5%) to three or more antimicrobials. In the eae-positive E. coli isolates, one isolate from the fallen fruit in Meixian exhibited resistance to four antimicrobials, while the other one in Zhouzhi was susceptible to all antimicrobials tested.
Minimum inhibitory concentrations (μg/mL) determined via agar dilution in accordance with Clinical and Laboratory Standards Institute (CLSI, 2013).
Biofilm formation
The Congo red agar method showed that all 68 E. coli isolates were in red or white, and none of them showed black colonies, suggesting that none of them have the ability to produce biofilm formation under this condition.
Random amplified polymorphic DNA (RAPD)
The UPGMA dendrogram based on RAPD profile (Supplementary Fig. S1; Supplementary Data are available online at
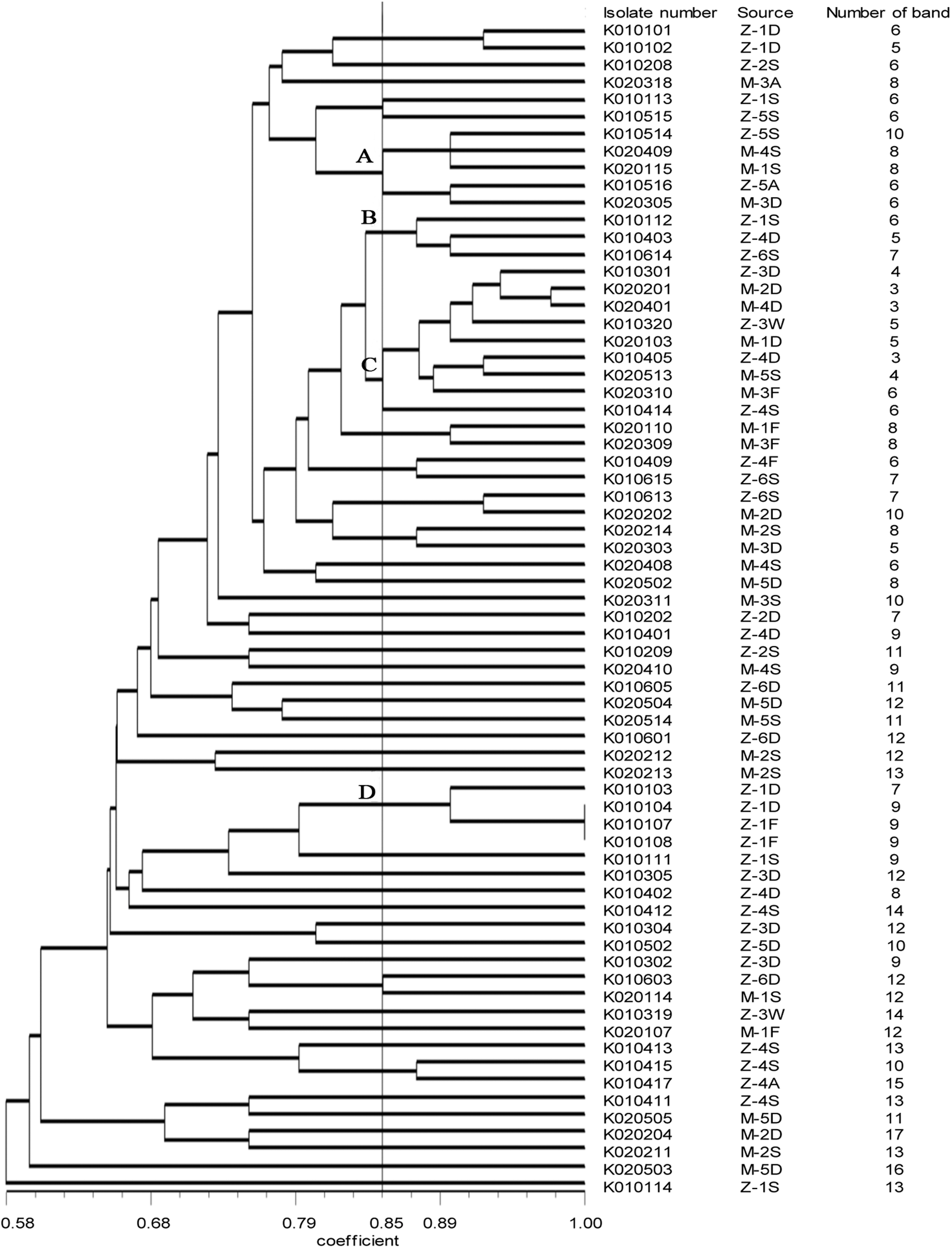
Dendrogram of random amplified polymorphic DNA patterns showing the relatedness of Escherichia coli isolates recovered from orchards.
MLST
MLST analysis was performed for two EPEC strains isolated in the present study and three STEC isolated in 2012. Two EPEC belonged to two different sequence types (STs) (Table 3). One is ST3068 and the other has a new combination of allele types and was given a novel ST-type as ST4614. Three STEC of 2012 have two ST types. Two of them are ST201 and another one has a new combination of allele types and was given a novel ST-type as ST4613.
New ST type.
Discussion
This study aimed to continue a rolling program to monitor the contamination of three foodborne bacteria in the kiwifruit orchards. The rate of samples positive for E. coli in kiwifruit orchards is close to the rate (37.4%, 130/348) in the year 2012 (Feng et al., 2014). However, no S. aureus was found in this study, while 5 (5/348, 1.4%) samples were contaminated by S. aureus in 2012. This is in contrast to other studies showing that the irrigation water, soil, and air are possible sources of S. aureus and Salmonella (Nakamura et al., 1997; Lapidotand and Yaron, 2009; Ijabadeniyi et al., 2011; Micallef et al., 2012).The present study indicates that E. coli is one of the major microbial contamination in kiwifruit orchards, which may be due to the use of fecal materials as fertilizers. In China, some farmers prefer to use the feces of livestock as fertilizer to reduce fertilizer expenses. This practice greatly increases the chance of E. coli contamination and should be discouraged. Only two (2.9%) E. coli isolates were found to carry the eae gene, and other virulence genes for diarrheagenic E. coli were not detected. We lack the evidence for confirming further that if the feces of animals or birds are the source of these EPEC isolates (Fukushima et al., 1999). Another possible source of EPEC is fruit flies. Fallen fruit is susceptible to spoilage and becomes attractive to fruit flies, which were easily contaminated externally and internally with E. coli after contact with the bacterium source (Janisiewicz et al., 1999). The MLST results showed that one EPEC isolate from fallen fruit shared the same ST type with an human clinical isolate in Switzerland in the MLST databases (
The antimicrobial resistance rates of E. coli isolates in this study were a little higher than those reported for E. coli isolates recovered from kiwifruit orchards in 2012 (Feng et al., 2014), except that the resistance to trimethoprim–sulfamethoxazole was lower than in the previous study. Isolates from identical clusters showed similar resistance patterns (data not shown). None of E. coli showed the ability to produce biofilm formation, which was different from isolates recovered from dairy, fish processing, poultry, meat, and the ready-to-eat food industry (Srey et al., 2013). It has been confirmed that the surface properties of tissue, nutrient, and water availability strongly influence the resulting biofilm structure (Ramey et al., 2004). The environments in kiwifruit orchards may be not suitable for biofilm formation, and this makes it less difficult for growers to maintain hygiene in orchards.
RAPD typing is a fast, high-resolution, and inexpensive method to subtype microbial pathogens compared with other typing methods, such as pulsed-field gel electrophoresis (Nielsen et al., 2014). The RAPD patterns revealed genetic diversity between different isolates. Several identical patterns were found from different types of samples in the orchard. Cluster A, containing isolates from the same orchard, revealed the possibility of cross-contamination between air and soil. In particular, the air closer to the ground is more likely to be contaminated by E. coli. This is a new contamination route compared with the last study. Cluster C, containing isolates from the same orchard, revealed the close connection between E. coli from soil and E. coli from irrigation water, and the close connection between E. coli from fallen fruit and E. coli from soil, which has been indicated in our last study. These indicated that irrigation water might be a reservoir and a transmission pathway for contamination. In addition, fallen fruit may be contaminated by soil, which is a natural habitat of E. coli.
Conclusions
This study demonstrated the frequent presence of E. coli in the kiwifruit orchards in Shaanxi Province, China, while no S. aureus and Salmonella were detected. E. coli was widely distributed such as in fresh fruit, fallen fruit, soil, irrigation water, and even air. Irrigation water and soil could be potential sources of microbial contamination in the kiwifruit orchard. These findings provide useful information for kiwifruit growers to establish proper management practices that help minimize the chance of microbial contamination during the growth stage. For instance, reduction of the use of raw manure and use of good-quality irrigation water are useful to reduce contamination in the field.
Footnotes
Acknowledgments
This work was supported in part by the Twelve-Five Science and Technology Support Program (No. 2012BAH30F03 and 2012BAK17B06), New Century Excellent Talent Support Plan (NCET-13-0488), International Collaboration Partner Plan (No. A213021203) in Northwest A&F University, and Science and Technology Development Plan Program of Shaanxi Province (No. 2013KJXX-16).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.