
Abstract
Group A rotavirus is responsible for inducing severe diarrhea in young children worldwide. Rotavirus vaccines are used to control the disease in many countries. In the current study, the sequences of human rotavirus G and P types in Saudi Arabia are reported and compared to different relevant published sequences. In addition, the VP4 and VP7 genes of the G1P[8] strains are compared to different antigenic epitopes of the rotavirus vaccines. Stool samples were collected from children under 2 years suffering from severe diarrhea. Screening of the rotavirus-positive samples was performed with rapid antigen detection kit. RNA was amplified from rotavirus-positive samples by reverse transcriptase polymerase chain reaction assay for both VP4 and VP7 genes. Direct sequencing of the VP4 and VP7 genes was conducted and the obtained sequences were compared to each other and to the rotavirus vaccines. Both G1P[8] G1P[4] genotypes were detected. Phylogenetic analysis revealed that the detected strains belong to G1 lineage 1 and 2, P[8] lineage 3, and to P[4] lineage 5. Multiple amino acid substitutions were detected between the Saudi RVA strains and the commonly used vaccines. The current findings emphasize the importance of the continuous surveillance of the circulating rotavirus strains, which is crucial for monitoring virus evolution and helping in predicting the protection level afforded by rotavirus vaccines.
Introduction
D
Rotavirus is among the major viral agents that cause human diarrhea (Desselberger, 2014). Nearly all children have experienced at least one rotavirus infection by age 5 (Parashar et al., 2003). Infection with rotavirus is more frequent in infants and children <2 years old, with a maximum incidence among children 0–12 months.
The virus is a member of the family Reoviridae, and possesses segmented double-stranded RNA (11 segments) and a triple-layered nonenveloped icosahedral symmetry. The virus encodes six structural (VP1, VP2, VP3, VP4, VP6, VP7) and five to six nonstructural proteins (NSP1–NSP5/6) (Estes and Greenberg, 2013).
Rotaviruses exist in eight different species (A–H), with the species A–C known to affect humans, with the A group being the most dominant worldwide (Diggle, 2007). The classification of group A rotavirus depends on the variation in its surface viral proteins: VP4 and VP7. The binary system of rotavirus classification is based on differentiating P-types (protease-sensitive protein of the VP4) and G-types (G for VP7 glycoprotein). Although there are at least 27 G-types and 37 P-types existing worldwide (Matthijnssens et al., 2011), the G1P[8], G2P[4], G3P[8], G4P[8], G9P[8], and G12P[8] combinations are the most common types (Banyai et al., 2012; Estes and Greenberg, 2013; Desselberger, 2014). The differences in the surface and backbone gene configurations suggest differences in the antigenic structure of the whole protein composition and might have implications for vaccination programs.
VP7 and VP4 gene segments undergo extensive genetic variation due to accumulated point mutations and gene rearrangement (Gouvea et al., 1990; Gentsch et al., 1992). Variation may also arise from reassortment among different rotaviruses, including animal rotaviruses (Kirkwood, 2010).
Two licensed rotavirus vaccines have been commercially available worldwide since 2006 including Rotarix, human G1P[8] monovalent rotavirus vaccine, and pentavalent RotaTeq vaccine (bovine reassortant chimeric virus strains expressing human G1–G4, and P[8]) (Dennehy, 2008). Although both vaccines elicit protective immunity to homotypic and heterotypic rotavirus strains (Leshem et al., 2014), there is a continuous need for a better understanding of the changing molecular characteristics of rotavirus strains and their degree of homology to the vaccine strains (Gentsch et al., 1992; Iturriza-Gomara et al., 2009, 2011; Braeckman et al., 2012).
In Saudi Arabia, the prevalence of rotavirus ranges from 10% to 46% and is common in children below 2 years of age (Kheyami et al., 2006). Rotarix vaccine has recently been introduced in the obligatory national Saudi Arabia vaccination program in January 2013. Although rotavirus is the most common cause of diarrhea in children in Saudi Arabia, only a few studies have screened the molecular characterization of rotavirus strains circulating in different areas of Saudi Arabia (Kheyami et al., 2008a, b). The present study aimed to determine the distribution of G and P genotypes via direct gene sequencing and to compare their relatedness to different rotavirus vaccines.
Materials and Methods
Ethics statement
The study design was approved by the medical ethics board of both the College of Medicine, Taif University and by Taif Pediatric Hospital in accordance with the guidelines for the protection of human subjects. Informed written consent was provided by the next of kin of the participants involved in the study.
Sample collection and processing
Rotavirus surveillance was conducted at the Taif Pediatric Hospital in 2013. Children <2 years of age who were admitted with acute gastroenteritis, including episodes of diarrhea, watery stools/mucus in stools, and vomiting were included in the study. Rectal swabs from 80 affected children were collected from the Governmental Pediatric Hospital, Al-Taif, Saudi Arabia. The children's ages ranged from 2 months to 2 years. Clinical manifestations and case histories were recorded. Individual swabs were kept in 1-mL sterile saline containing gentamicin sulfate. Swabs were routinely processed and kept at −80°C until further analysis.
Rotavirus A antigen detection
Rectal swab specimens were screened first for rotavirus group A antigen using a Rota/Adeno rapid chromatographic antigen detection kit (RapiCard™ 176067-1; Diagnostic Automation/Cortez Diagnostics, Inc., USA).
RNA extraction and reverse transcriptase polymerase chain reaction (RT-PCR)
Viral RNA was extracted from 10% (wt/vol) rectal swab supernatants using the viral RNA extraction kit (Qiagen, Germany), according to the manufacturer's instructions. Viral RNA was extracted from samples that reacted positively to a rapid chromatographic antigen detection test. The extracted RNA was used as a template for conventional RT-PCR using one-step RT-PCR kit (Koma Bioteck, Korea). Amplification of the partial VP4 gene (VP8* region: 663 bp) was carried out using the oligonucleotide primers: VP4F 5′-TATGCTCCAGTNAATTGG-3′ (nucleotide [nt] position 132–149) and VP4R 5′-ATTGCATTTCTTTCCATAATG-3′ (nt 775–795), as described previously (Simmonds et al., 2008). The partial VP7 gene (884 bp) was amplified using oligonucleotide primers specific for conserved regions of the VP7 gene: VP7-F 5′-ATGTATGGTATTGAATATACCAC-3′ (nt 49-71) and VP7-R 5′-AACTTGCCACCATTTTTTCC-3′ (nt 914-933), as described previously (Iturriza-Gomara et al., 2001). The RT-PCR amplicons were subjected to gel electrophoresis (1.5%).
Direct sequencing and gene sequence analysis
RT-PCR amplicons were purified using a gel-extraction kit (Koma Biotek, Korea) and used as templates for direct sequencing. Different gene sequences were analyzed using MEGA 5.1. Homology BLASTn searches for individual gene sequences were performed using highly similar sequences (megablast) against available published rotavirus sequences in the GenBank databases in January 2015 using default algorithm parameters. CLUSTAL W multisequence analysis was performed and phylogenetic trees constructed using the neighbor-joining (NJ) method with 1000 bootstrap replicates. NJ was chosen since it is among the trees that were shown to be least erroneous in inferring topology (Woolley et al., 2008). The sequence identity percentage was performed using the free online tool available at
Accession numbers
The nucleotide sequences for genes described in this study have been deposited in GenBank under the following accession numbers: RVA/Human/SA/Taif-1/2013/G1P[8] (KP281273, KP281279), RVA/Human/SA/Taif-2/2013/G1P[8] (KP281274, KP281280), RVA/Human /SA /Taif-3/2013/G1P[8] (KP281275, KP281281), RVA/Human/SA/Taif-4/2013/G1P[8] (KP281276, KP281282), RVA/Human/SA/Taif-5/2013/G1P[8] (KP281277, KP281283), RVA/Human/A/Taif-6/2013/G1P[4](KP281278, KP281284), RVA/ Human/SA/Taif-7/2013/GxP[8] (KP281285) and RVA/Human/SA/Taif-8/2013/GxP[8] (KP281286).
Results and Discussion
The rotavirus epidemiology in Saudi Arabia from 1982 to 2003 was reviewed, with infection prevalence in children being found to range from 10% to 46% with a median of 30% (Kheyami et al., 2006); however, lower rates of prevalence were also recorded (elAssouli et al., 1996; Ghazi et al., 2005). Effective control depends upon an accurate understanding of the burden of the disease and the relative importance of circulating genotypes. In four studies where strains had been G-typed, the G1 genotype was found to be predominant, followed by G4, G3, and G2, but no sequence data are available for the circulating G/P types (Kheyami et al., 2006).
In the current study, antigen detection was used to screen the rotavirus and successfully detect rotavirus in 12/80 samples. Rotavirus group A antigen detection followed by genotyping of positive samples was used in the diagnosis of rotavirus (Ramani et al., 2010; Arakawa et al., 2011). This approach broadens the detection level, a theory that has been confirmed by Maes et al., who successfully used human group A rotavirus assay for the detection of bovine rotavirus with a sensitivity and specificity of 87.0% and 93.6%, respectively (Maes et al., 2003). Upon testing positive samples using RT-PCR for both G/P types, only 8/12 and 6/12 showed positive reactions for P and G genotypes, respectively. Taif-7 and Taif-8 strains could not be assigned to a G-type specificity. However, the fact that some samples that were positive on the antigen detection test were negative on RT-PCR does not indicate the relative sensitivity of the two tests. The negative reaction in RT-PCR might be related to either the presence of point mutations not recognized by the primers used for the detection of P and G genotyping, or as yet unidentified G/P genotypes. Furthermore, earlier studies have demonstrated a reduced sensitivity of the PCR for the detection of G/P genotypes due to primer mismatch, since 53.6% and 8% were not successfully genotyped for G and P genotypes, respectively (Bonkoungou et al., 2011). In addition, a high percentage of the G1 strains worldwide could not have been molecularly typed using a single RT-PCR strategy (Parra and Espinola, 2006). RT-PCR is a highly sensitive and specific assay used for rotavirus genotyping (Fischer and Gentsch, 2004); however, continuous gene mutations in the virus may reduce the sensitivity of the assay over time. Consequently, the primers used for genotyping should be changed regularly (Simmonds et al., 2008).
BLAST analysis of the different Saudi RVA strains showed that five strains belong to G1P[8], one strain to G1 P[4], and two strains belong to GxP[8]. Phylogenetic analysis revealed that the 7/8 VP4 sequences belong to P[8] lineage 3, while the 1/8 sequence, RVA/Human/SA/Taif-6/2013, belongs to the P[4] lineage 5 (Fig. 1). Genotype P[8] strains circulate globally in four distinct lineages, although several studies have revealed that in the 2000s the predominant circulation was of the P[8]-3 lineage (Phan et al., 2007; Espinola et al., 2008; Le et al., 2010; Cashman et al., 2012). In the present study the P[8] lineage 3 was found in 7/8 isolates, thus indicating the circulation of this sublineage in Saudi Arabia.
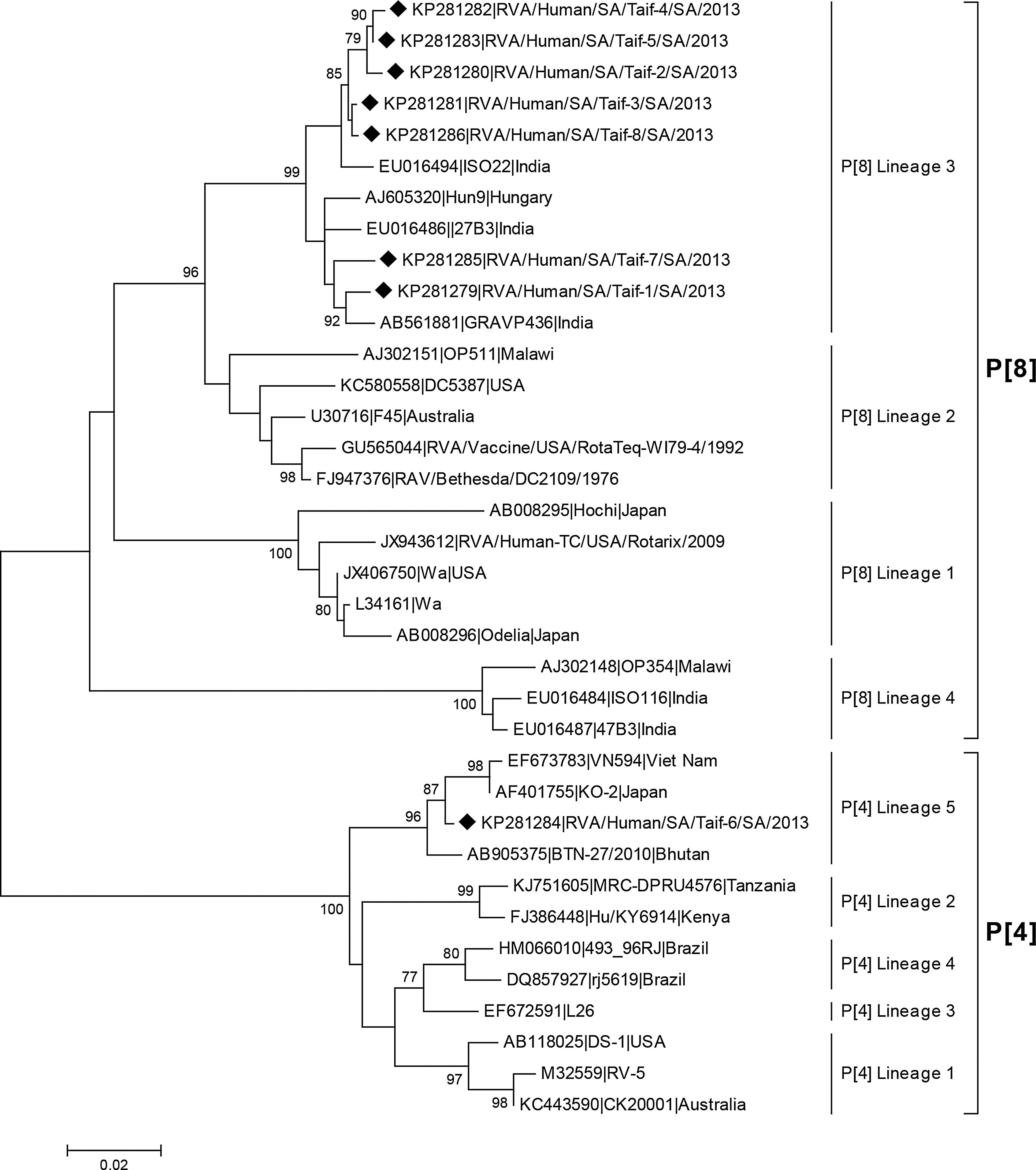
Phylogenetic tree of nucleotide sequences of VP4 gene of Saudi RVA strains in comparison to relevant published strains. Neighbor joining (NJ) tree was constructed using MEGA 5.1 freeware. Bootstrap analysis with 1000 replicates was performed. Only bootstrap values above 70% are shown. The symbol (♦) was used to denote strains isolated in the current study.
Meanwhile, all the VP7 sequences showed that the Saudi RVA strains belong to G1 genotypes, 5/6, to G1 lineage 2 and 1/6, RVA/Human/SA/Taif-6/2013, to G1 lineage 1 (Fig. 2). Previous findings from Saudi Arabia and also from worldwide reported that the G1P[8] is the commonly isolated RVA genotype in human populations (Kheyami et al., 2006; O'Ryan, 2009). The current data also suggest that sequence variations do exist among different rotavirus strains in both VP4 and VP7 (Figs. 3 and 4).
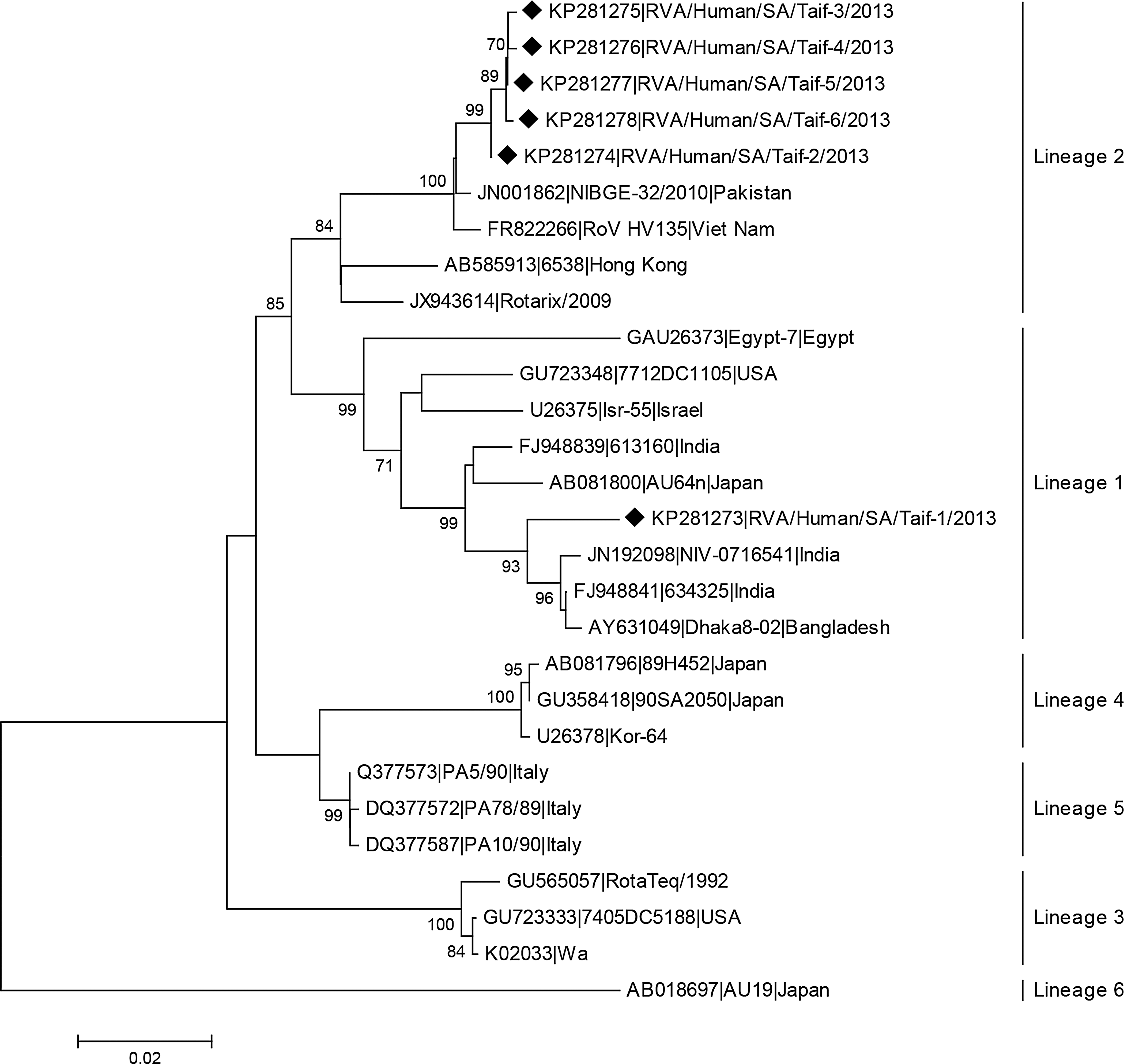
Phylogenetic tree of nucleotide sequence of VP7 proteins of Saudi RVA strains in comparison to RVA of different G1 lineages. Neighbor joining (NJ) tree was constructed using MEGA 5.1 freeware. Bootstrap analysis with 1000 replicates was performed. Only bootstrap values above 70% are shown. The symbol (♦) was used to denote strains isolated in the current study.
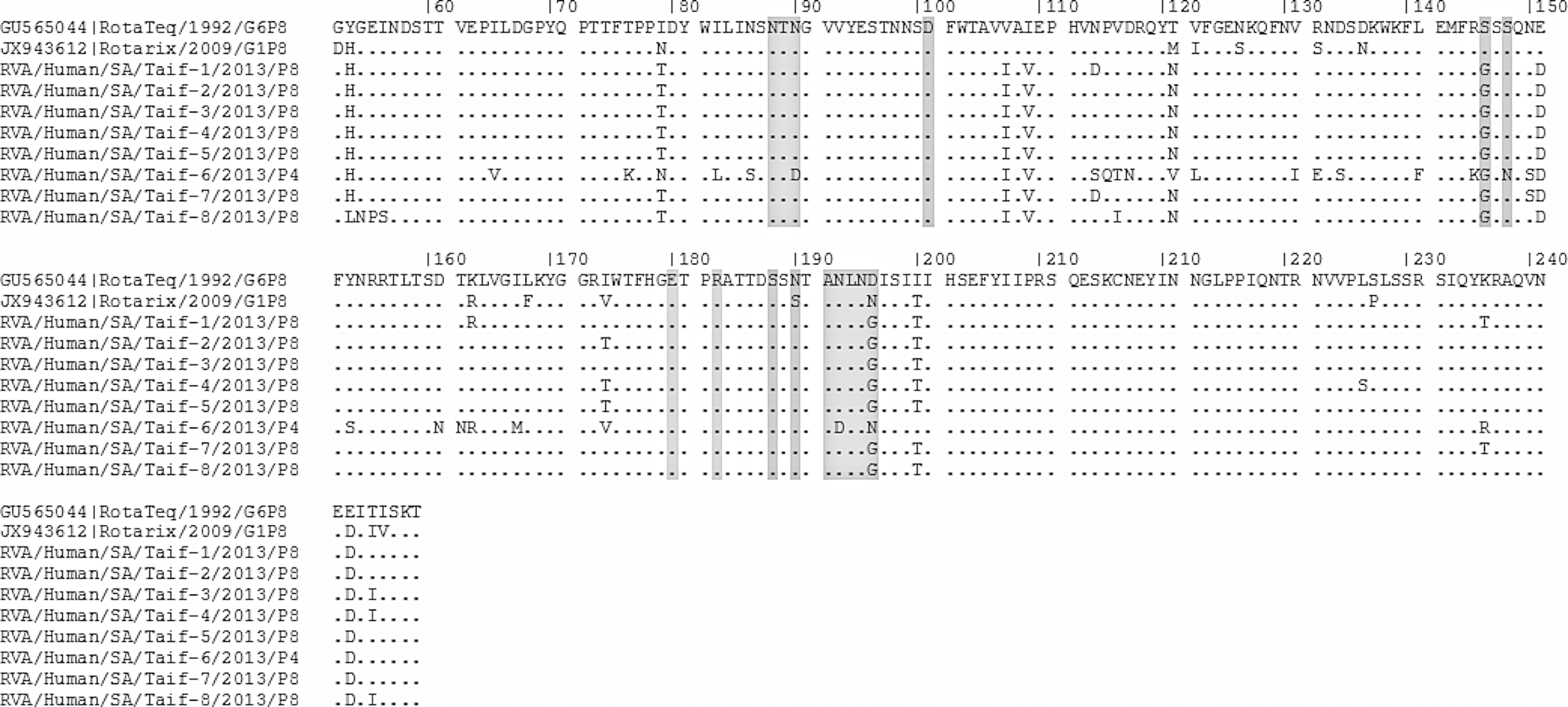
Deduced amino acid sequences of VP4 proteins of the different rotavirus A strains in comparison to the rotavirus vaccines. Amino acid residues in epitopes 1–4 are highlighted in gray.

Deduced amino acid sequences of VP7 proteins of the different rotavirus A strains in comparison to the rotavirus vaccines. Amino acid residues in the following epitopes: 1a (87, 91, 94, 96–100, 104, 123, 125, and 129–130), 1b (201, 211–213, 238, and 242), and 2 (143, 145–148, 190, 217, and 221) are highlighted in gray.
Two live attenuated RVA vaccines, Rotarix and RotaTeq, have been licensed for use in many countries worldwide. Rotarix is an attenuated human G1P[8] rotavirus strain while RotaTeq is a live attenuated human–bovine reassortant RVA that includes five major genotypes: G1, G2, G3, G4, and P[8]. In the current study, we investigated the sequence diversity between the VP4 and VP7 of the Saudi RVA strains, and the relevant proteins in the Rotarix (G1 lineage 2, P[8] lineage 1) and RotaTeq (G1 lineage 3, P[8] lineage 2).
CLUSTAL-W alignments of the VP4 (VP8* of the P[8]) amino acid sequences of Saudi strains with those of vaccine strains showed that the RotaTeq vaccine is more related to the Saudi strains than the Rotarix vaccine: RotaTeq showed 92–94% identity to all Saudi strains except Taif-6, which showed only an 83% homology. Rotarix, meanwhile, showed 88–91% identity to all Saudi strains except Taif-6 (P[4]), which showed only an 82% homology (data not shown). This finding is expected since the P[8] strain of the RotaTeq vaccine belongs to a different genetic lineage (lineage 2). CLUSTAL-W alignments of the VP7 (G1) amino acids, meanwhile, showed that Rotarix had higher amino acid identities to the Saudi strains than the RotaTeq vaccine, with the former showing (96–97%) except for RVA/Human /SA/Taif-1/2013/G1P[8], where it showed a 92% identity, while the latter showed a 94–95% identity, except for RVA/Human /SA/Taif-1/2013/G1P[8], which showed a 91% identity.
Amino acid variations at the neutralization escape mutation sites within the VP7 and VP4 antigenic epitopes have been previously determined (Dormitzer et al., 2004; Aoki et al., 2009; Kulkarni et al., 2014). Any amino acid substitutions in these antigenic regions are highly likely to enhance immune escape, thereby reducing the protection afforded by the rotavirus vaccination (Simmonds et al., 2008; Kulkarni et al., 2014).
Rotavirus VP4 protein is subject to proteolytic cleavage into two subunits: VP8 (aa 1–247) and VP5 (aa 248–776) (Estes and Greenberg, 2013). Saudi strains differ from RotaTeq by at least 10 amino acids (Fig. 3), with two of them (S146G and D196G/N) being in the epitope 1 antigenic site (Table 1). In respect to the Rotarix vaccine, however, they showed at least 17 different amino acids (Fig. 3), with three of them in the epitope 1 antigenic site (S146G, S190N, and N196G) and three in the epitope 3 (S126N, S132R, and N136D) (Table 1).
Different amino acid residues are highlighted in gray.
Taif-6 strain, meanwhile, belongs to P[4], which showed at least five amino acid substitutions in the epitope 1 site compared to both the RotaTeq and Rotarix vaccines, while in the epitope 3 site there were six amino acid substitutions from the RotaTeq vaccine and eight from the Rotarix vaccine, as well as a single amino acid substitution in epitope 4 (Table 1).
The Taif-7 strain also showed N150S substitution in the epitope 1 site compared to both the Rotarix and the RotaTeq vaccines (Table 1). In the epitope 3 site, 3/7 P [8] Saudi strains showed a single individual amino acid substitution: Taif-1 and Taif-7 showed (N114D), while Taif-8 showed V116I substitution from both the RotaTeq and Rotarix vaccines.
McDonald et al. (2012) reported that three amino acid residues (I79, T121, and K163) of RotaTeq VP4 differed from nearly every P[8] strain isolated since 2000. These amino acid residues are located in or near neutralization domains 8-1, 8-3, or 8-4 of VP8* (McDonald et al., 2012). The authors suggested that the location of these three residues influences the antigenicity of VP4. In the Saudi RVA strains, I79T/N and T121N/V were detected in all strains, while K163R substitution was detected in only two of them (Fig. 3).
The VP7 protein possesses 7-1a, 7-1b, and 7-2 as antigenic epitopes (Aoki et al., 2009). The Saudi strains did not show any amino acid substitutions in different epitopes of the VP7 protein, either from each other or from the Rotarix vaccine; however, they did show two amino acid substitutions in epitope D97E in epitope 1a and S147N substitution in epitope 2 from the RotaTeq vaccine (Fig. 4). Interestingly, D97E and S147N amino acid substitutions are critical in retaining the neutralization epitopes (Coulson and Kirkwood, 1991). D97E is a critical substitution of a negatively charged polar amino acid to an uncharged polar amino acid that may help in the emergence new rotavirus strains (Doan et al., 2011).
In contrast, N94S amino acid substitution was detected only in the Taif-1 strain that belongs to the G1 sublineage 1. Such residue is essential for the discrimination of G1 monotypes (Coulson and Kirkwood, 1991) and also for distinguishing lineages 1 and 2 (Diwakarla and Palombo, 1999). N94S is also correlated with resistance against the virulent G1 strain (Green and Kapikian, 1992).
A considerable variability in amino acids was detected in the VP7 sequences of the G1 rotaviruses analyzed in this study in comparison to the vaccine strains. Apart from the variability detected in the three antigenic epitopes sites, Taif-2, 3, 5, and 6 strains (G1 lineage 2) showed 13 and 7 amino acid substitutions from RotaTeq and Rotarix vaccines, respectively, while Taif-4 (G1 lineage 2) showed an additional T277S amino acid substitution (14 aa substitutions) from both the Rotarix and RotaTeq vaccines (Fig. 4). In contrast, Taif-1 (G1 lineage 1) showed 18 and 17 amino acid substitutions from RotaTeq (G1 lineage 3) and Rotarix (G1 lineage 2) vaccines respectively (Fig. 4). One of the limitations of the current study was the inability to evaluate the impact of the amino acid substitutions on both virus virulence and protection level afforded by the rotavaccines.
In conclusion, the present study describes differences between the rotavirus strains circulating in Al-Taif, Saudi Arabia (mainly G1P[8]). Comparison of the antigenic differences between the Saudi strains with the Rotarix and RotaTeq vaccines revealed significant differences in different epitopes. Continuous monitoring of the rotavirus strains should be regularly conducted to screen the subgenotypic lineages of rotavirus strains.
Footnotes
Acknowledgments
This work was supported by Taif University, Grant No. 3331-435-1.
Disclosure Statement
No competing financial interests exist.