
Abstract
Emetic toxin–producing Bacillus cereus group species are an important problem, because the staple food for Korean is grains such as rice. In this study, we determined the prevalence (24 of 129 isolates) of emetic B. cereus in 36,745 stool samples from sporadic food-poisoning cases in Korea between 2007 and 2008. The toxin gene profile, toxin production, and biofilm-forming ability of the emetic B. cereus isolates were investigated. Repetitive element sequence polymorphism polymerase chain reaction fingerprints (rep-PCR) were also used to assess the intraspecific biodiversity of these isolates. Emetic B. cereus was present in 0.07% of the sporadic food-poisoning cases. The 24 emetic isolates identified all carried the nheABC and entFM genes and produced NHE enterotoxin. However, they did not have hemolysin BL toxin or related genes. A relationship between biofilm formation and toxin production was not observed in this study. The rep-PCR fingerprints of the B. cereus isolates were not influenced by the presence of toxin genes, or biofilm-forming ability. The rep-PCR assay discriminated emetic B. cereus isolates from nonemetic isolates, even if this assay did not perfectly discriminate these isolates. Further study on emetic isolates possessing a high degree of diversity may be necessary to evaluate the performance of the subtyping assay to discriminate emetic and nonemetic B. cereus isolates and could provide a more accurate indication of the risk from B. cereus strains.
Introduction
B
In recent years, there has been increased awareness of the potential risk of emetic food poisoning caused by B. cereus in Korea, because rice and grains are consumed in large quantities and contaminated with emetic toxin–producing B. cereus (Kim et al., 2010b). However, only 2 cases of the emetic type of food poisoning (Kim et al., 2009, 2010b) were reported among 29 cases of food-poisoning outbreaks caused by B. cereus (
The persistent contamination of the industrial food processing system by B. cereus group bacteria (Kotiranta et al., 2000) may facilitate its involvement in gastroenteritis through contamination of food products. This persistence is due to spores, which may survive pasteurization and heating (Kamat et al., 1989; Larsen and Jorgensen, 1999), and to biofilms, which have been shown to be highly resistant to cleaning procedures (Peng et al., 2002). Many bacteria including Bacillus, Salmonella, Listeria, Staphylococcus, and Escherichia are capable of forming biofilms on the surface of foods, cooking utensils, metals, glass, plastic, or rubber (Ryu and Beuchat, 2005; Elhariry, 2008). Biofilms are also suspected to be involved in bacterial pathogenicity, because they may form on the host epithelia (Macfarlane and Dillon, 2007). Furthermore, biofilm formation might be enhanced through its effect on quorum sensing (cell-to-cell signaling; Auger et al., 2006; Hsueh et al., 2006). However, there have been no reports on whether biofilm formation by B. cereus could be connected to enterotoxin or emetic toxin produced by B. cereus.
A rapid and reliable molecular subtyping method to epidemiologically trace B. cereus is important in monitoring potential foodborne pathogens. Several methods have been used for subtyping emetic B. cereus for >10 years (Pirttijärvi et al., 1999; Ehling-Schulz et al., 2005a). Chon et al. (2012) have successfully distinguished emetic toxin–producing B. cereus strains from nonemetic strains using an automated repetitive-element-sequence polymorphism-based PCR (rep-PCR) system, DiversiLab™ (BioMérieux, Marcy l'Etoile, France), which is commercially available but expensive. Therefore, it is necessary to determine whether the conventional rep-PCR assay using general noncoding repetitive sequences, which is simple and more cost effective than the DiversiLab™ system, because it is able to use existing electrophoresis apparatus, can sufficiently differentiate subtypes of emetic B. cereus strains like the results from DiversiLab™.
In this study, we investigated the prevalence of emetic toxin–producing B. cereus in sporadic food-poisoning cases in Korea and performed in-depth characterization of the isolated emetic B. cereus strains based on genotypic and phenotypic traits, biofilm-forming ability, and concurrently assessed the intraspecific biodiversity of these isolates with a repetitive sequence-based PCR system.
Materials and Methods
Bacterial isolates and culture conditions
We used a total of 36,745 clinical stool samples that were referred to the Korea Center for Disease and Prevention (KCDC; Jeollabuk-do, South Korea) between 2007 and 2008. All samples were streaked onto the surface of B. cereus–selective mannitol–egg yolk–polymyxin (Difco, Becton Dickinson, Sparks, MD) agar plates and incubated at 37°C for 24 h. A pink colony was selected for culture on tryptone soya agar (Oxoid, Denka Seiken Ltd, Tokyo, Japan) and blood agar (BA; 5% sheep blood, Komed, Seoul, Korea) at 37°C for 24 h. Cultures of presumptive colonies that developed β-hemolysis on BA were confirmed with Gram staining. The identities of Gram-positive isolates were determined with a VITEK BCL card (bioMérieux, Marcy l'Etoile, France). All confirmed clinical strains were then screened for emetic genes using PCR amplification described below. According to previous studies, B. cereus ATCC 14579 (diarrheal) and B. cereus KFDA 229 (emetic) were used as reference strains (Hansen and Hendriksen, 2001; Lee et al., 2008).
PCR amplification of enterotoxin and emetic toxin genes
The genomic DNA for the PCR assays was extracted with a DNeasy Tissue kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Extracted genomic DNA was quantified with the NanoDrop 2000 (Thermo Scientific, Wilmington, DE). For detection of B. cereus toxin genes, the primer pairs and PCR reaction conditions were in accordance with a previous report (Lee et al., 2012). Each PCR reaction was conducted in triplicate, and the amplified products were separated by electrophoresis on 2% agarose gel at 100 V for 30 min. The gels were stained with 0.5 μg/mL ethidium bromide solution and visualized with a UV transilluminator (Gel Doc XR system, Bio-Rad, Hercules, CA).
Toxin production analysis
To determine emetic toxin production, the Hep-2 (human larynx carcinoma cell) vacuolation assay was performed. Each isolate was grown in 10% skim milk medium (SMM; Difco). Overnight cultures of B. cereus were added to 50 mL of each medium (10% SMM) in 500-mL conical flasks and incubated at 30°C with orbital shaking (200 rpm) for 18 h. Samples (50 mL) were centrifuged at 8000 × g at 4°C for 20 min. Supernatants were autoclaved at 121°C for 15 min to sterilize the samples and to denature heat-labile toxins (enterotoxins). The serial logarithmic dilution of the heat-treated supernatant fluids was added to the Hep-2 cells in 96-well microtiter plates. The appearance of vacuolation was monitored after incubation for 8 h at 37°C in 5% CO2 (Finlay et al., 1999). The emetic toxin assay was conducted only in strains harboring an emetic toxin gene.
To characterize enterotoxin-producing emetic B. cereus isolates, the Bacillus Diarrheal Enterotoxin Visual Immunoassay (BDE-VIA) kit (Tecra Diagnostics, Roseville, Australia) and the B. cereus Enterotoxin Reversed Passive Latex Agglutination (BCET-RPLA) kit (Oxoid, Denka Seiken Ltd, Japan) were used for the detection of NHE and HBL enterotoxins, respectively. The VIA kit measures the NheA component of the NHE complex, whereas RPLA measures the L2 component of the HBL complex (Granum et al., 1996). The tests were done according to the manufacturers' instructions.
Biofilm assays
The biofilm assay was performed as described by O'Toole and Kolter (1998) with a few modifications. Pre-cultures in the exponential phase of growth were inoculated at an optical density (600 nm; OD600) of 0.01 into fresh Luria-Bertani medium (Difco) in 96-well microtiter plates and allowed to incubate at 30°C for 24 h. The microtiter plates were gently washed twice with sterile distilled water and dried at room temperature. Each well was stained with 200 μL of 1% (wt/vol) crystal violet solution for 20 min and rinsed three times with sterile water. The dye was solubilized by the addition of 200 μL of 95% ethanol. The absorbance at 595 nm of the solubilized dye was subsequently determined. All the experiments were performed five times and the results were expressed as means ± standard deviation (SD). The differences between control and test were analyzed using Student t-test. P values < 0.05 were considered to be statistically significant.
rep-PCR
rep-PCR fingerprinting was used for chromosomal comparisons of B. cereus isolates. The (GTG)5 primer (5′-GTGGTGGTGGTGGTG-3′) was performed at the following amplification conditions: an initial denaturation step at 95°C for 2 min was followed by 30 cycles (30 s at 94°C + 1 min at 40°C + 1 min at 60°C) and a final elongation step for 5 min at 65°C. The obtained PCR products were separated with 1.5% agarose gel electrophoresis in 1× Tris-acetate-EDTA (TAE) buffer at 50 V for 5 h. The rep-PCR profiles were visualized after staining with ethidium bromide under UV light, followed by digital image capturing with a charge-coupled device camera (Gel Doc XR system; Bio-Rad). The resulting fingerprints were analyzed with the FPQuest software package (Bio-Rad). Similarities between the digitized profiles were calculated with the Dice coefficient, and the average linkage (unweighted-pair group method with arithmetic mean dendrogram was obtained.
Results and Discussion
Prevalence of emetic B. cereus isolates in sporadic food-poisoning cases
A total of 129 B. cereus strains (0.35%) were isolated from 36,745 sporadic food poisoning cases that were referred to the KCDC over 2 years (from 2007 to 2008). We isolated a total of 35 B. cereus strains (27%) that carried the ces gene. Twenty-four of those isolates were B. cereus emetic isolates (24 of 129, 18.6%) that were positive for the ces gene and Hep-2 cell culture (Table 1). This result indicates that not all of the B. cereus isolates harboring the ces gene produced a toxin associated with emetic symptoms under the experimental conditions tested, but the potential risk for the emetic type of food poisoning associated with B. cereus existed. The Korea Food and Drug Administration (KFDA) has reported in 2011 that only 6.9% (2 of 29) of all B. cereus outbreak cases from 2001 to 2010 were the emetic type of food poisoning (Kim et al., 2009, 2010b). It was slightly higher than the reports of the KFDA in 2009 (
B. cereus enterotoxin reversed passive latex agglutination (BCET-RPLA) was used to detect hemolysin BL enterotoxin of B. cereus.
Bacillus diarrheal enterotoxin visual immunoassay kit was used to detect nonhemolytic enterotoxin of B. cereus.
All the experiments were performed five times and the results were expressed as means ± standard deviation (SD). The differences between control and test were analyzed using Student t-test. P values < 0.05 were considered to be statistically significant.
OD, optical density.
Recently in Korea, Chon et al. (2012) reported on seven B. cereus emetic isolates (7 of 206, 3.4%) carrying genes and toxin associated with the emetic syndrome. Lee et al. (2012) also found that 36 (92%) of 39 B. cereus isolates from Sunsik (a powdered mixture of roasted grains and other foodstuffs) carried the ces gene. These results are alarming because B. cereus isolates that have the emetic gene or emetic toxin are increasing, although the emetic type of food-poisoning outbreaks caused by B. cereus is low in Korea.
Enterotoxin production and profiling of related genes
Toxin gene profiling and toxin production by isolates in this study are summarized in Table 1. Various genotypic and phenotypic traits were investigated in the 24 emetic B. cereus isolates that have the emetic toxin gene and produce emetic toxin and the 11 nonemetic B. cereus isolates harboring only the emetic toxin gene. All emetic strains possessed both the nhe toxin-producing gene and entFM gene and also produced the NHE toxin determined with the Tecra BDE-VIA kit. Among them, 95.8% (23 out of 24) produced high levels of the toxin (production at an index of ≥4). However, these emetic strains were negative for the hbl toxin genes and cytK gene. Additionally, all of the nonemetic strains harbored the nheABC (100%) and entFM (100%) genes and were positive for NHE toxin. Of them, 5 nonemetic isolates (45.4%; 5 out of 11) had the hbl toxin-producing gene and cytK gene and were positive for the HBL toxin production reaction (with an index of ≥32) determined with the Oxoid BCET-RPLA kit.
The percentage of isolates carrying nheABC in this study is in good agreement with previous studies (Anderson et al., 2001; Kim et al., 2010a; Lee et al., 2012), which demonstrated that 90–100% of the emetic and nonemetic isolates possessed these genes. Guinebretière et al. (2002) also showed that almost all the strains from food poisoning and foodborne ecosystems harbored the nhe genes. In common with the nheABC genes, the presence of the entFM gene was observed in all B. cereus strains (Chon et al., 2012; Lee et al., 2012). The genes nheABC and entFM appeared to be broadly distributed among the emetic B. cereus strains (Ehling-Schulz et al., 2005b; Chon et al., 2012), regardless of the source. Tran et al. (2010) reported that entFM, cell wall peptidases, is related to bacterial biofilm formation and virulence of human intestinal bacteria besides foods. These findings suggest that exposure to emetic B. cereus strains infection has much higher risk potential.
Results of this study differed from those of our earlier studies with regard to hblACD in the emetic strains (Lee et al., 2012). In a previous study, the occurrence of the hblACD gene among emetic B. cereus strains isolated from Sunsik was 75%, which was much higher than the present study. However, Ehling-Schulz et al. (2005a) reported that none of the B. cereus strains in that study were positive for the hblACD gene or the HBL toxin, consistent with our study. Kim et al. (2010a) and Chon et al. (2012) reported hblACD-harboring emetic B. cereus strains, not consistent with our study. It appears that emetic strains are not related to the presence of hblACD genes.
The cytK gene was found in five (14.3%) of the B. cereus group isolates. This is consistent with previous results identifying the cytK gene among emetic isolates as ranging from 12.1% to 27.5% (Ehling-Schulz et al., 2005a; Kim et al., 2010a). Granum et al. (1996) reported that bceT was present in two of the seven food-poisoning B. cereus isolates. Mäntynen and Lindström (1998) tested 80 strains and found that only 1 strain was bceT-PCR positive. In contrast, the widespread occurrence (58.3%) of bceT among emetic B. cereus strains tested in this study is in agreement with the findings of Hsieh et al. (49%, 1999), and Ombui et al. (41.1%, 1997).
All the B. cereus isolates tested in this study contained at least one enterotoxin and its gene as well emetic toxin. It suggests that these isolates have the potential to cause diarrheal and emetic food poisoning. Also, this could be the reason that only two cases of the emetic food poisoning occurred in Korea.
Biofilm formation by B. cereus
Biofilm formation by the B. cereus isolates is shown in Table 1. We arbitrarily classified the formation of a biofilm by the B. cereus isolates as strong (OD595 > 0.5), intermediate (0.2 < OD595 < 0.5), and weak (OD595 < 0.2) (Auger et al., 2006; Lee et al., 2012). First, the ability of the reference strains B. cereus ATCC14579 and B. cereus KFDA250 to form biofilms on polystyrene 96-well microtiter plates was tested. We confirmed that KFDA250 formed strong biofilms while ATCC14579 formed weak biofilms on the polystyrene surfaces, which is in agreement with previously published reports (Auger et al., 2006; Hsueh et al., 2006). This microtiter plate assay detected differences in the biofilm-forming ability among the B. cereus strains. In 16 B. cereus isolates, the optical density at 595 nm (OD595) ranged from 0.094 to 0.176. Among the B. cereus isolates, 12 showed intermediate biofilm formation with an OD595 of 0.208–0.432. Of the remaining B. cereus isolates, seven strains showed strong biofilm formation with an OD of 0.527∼0.706. Interestingly, 11 isolates among the 16 weak biofilm formers (OD595 < 0.2) were the nonemetic B. cereus isolates. Five of these isolates were also positive for the hblACD gene and the HBL toxin, as was the ATCC14579 reference strain, which was associated with weak biofilm formation. However, grouping isolates according to emetic toxin production did not reveal any significant differences in biofilm production between the groups. It suggests that the biofilm-forming ability could rather be related to enterotoxin production than emetic toxin production. The ubiquitous B. cereus can easily contaminate food production or processing systems and forms biofilms that are highly resistant to cleaning procedures. Thus, use of human stools isolates harboring biofilm-forming ability may be more useful for understanding the interaction between human pathogens and foods.
Rep-PCR fingerprinting
A total of 37 B. cereus isolates including the 2 reference strains described earlier were characterized by rep-PCR assay of the genomic DNA to examine the genetic relationships among these strains. A total of 20 band patterns were obtained from the rep-PCR assay, 12 of which were represented by only 1 isolate (Fig. 1).
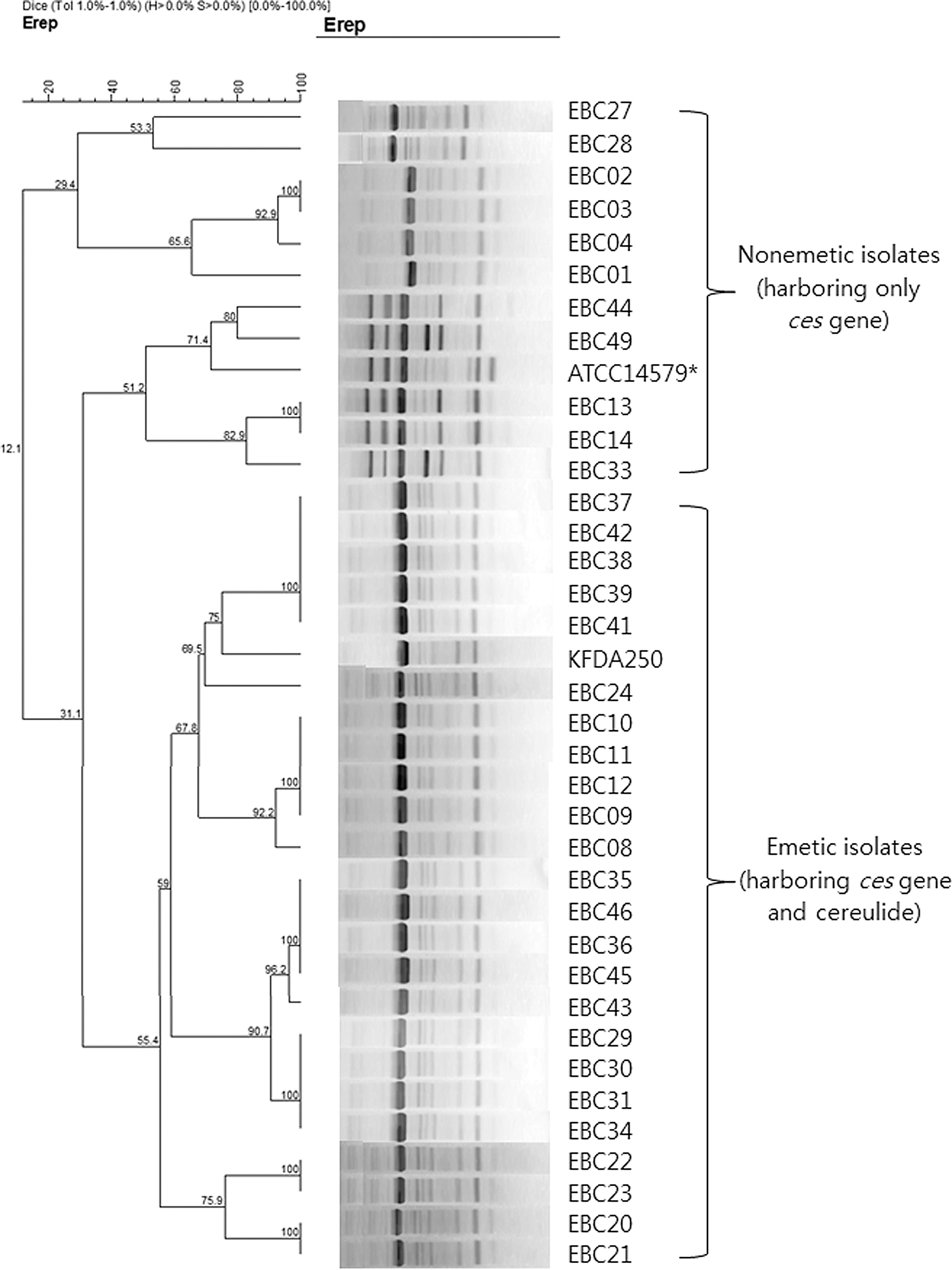
Dendrogram of the repetitive-element-sequence polymorphism-based polymerase chain reaction fingerprints obtained for the Bacillus cereus isolates from human stools. A pairwise percent similarity matrix was generated with the Dice coefficient, which was used to generate the unweighted-pair group method with arithmetic mean dendrogram. *Strain ATCC14579 (enterotoxin-producing B. cereus) was used as nonemetic B. cereus reference strain, and did not have the ces gene.
Although B. cereus isolates tested in this study were classified into various pulsotypes based on the rep-PCR banding patterns and are genetically unrelated, the dendrogram in Figure 1 shows that the rep-PCR assay discriminates emetic B. cereus isolates from nonemetic isolates. This result is agreement with a previous study by Chon et al. (2012) using DiversiLab™. All 25 emetic B. cereus isolates including the KFDA250 emetic reference strain produced identical patterns regardless of their origins, exhibiting a similarity level of 59–100%. The nonemetic isolates including the enterotoxin-producing reference strain ATCC14579, were scattered throughout various subgroups that had <31.1% similarity to the emetic isolates. Pirttijärvi et al. (1999) reported that emetic strains possess very similar ribotypes. Ehling-Schulz et al. (2005a) reported that emetic B. cereus shows a single evolutionary lineage of closely related strains using various genetic and phonetic typing methods. All emetic toxin–producing B. cereus in those studies did not have the HBL toxin or related genes (Pirttijärvi et al., 1999; Ehling-Schulz et al., 2005a). Our results are consistent with these findings. However, the emetic isolates in this study showed genetic diversity similar to enterotoxin-producing B. cereus and appear as nontypical traits that have lower genetic homogeneity than typical emetic strains in terms of constituting three distinct clusters of emetic B. cereus strains with a similarity level of 65%. Various researchers have reported that the emetic toxin synthetase (ces) gene is encoded on a large virulence plasmid, and the ces gene related to emetic toxin production can be transferred to different Bacillus strains (Pirttijärvi et al., 1999; Ehling-Schulz et al., 2005a; Hoton et al., 2005). Thus, our emetic B. cereus may be the result of the transfer of emetic toxin synthetase genes between enterotoxin-producing B. cereus.
Herein, we have reported that emetic B. cereus isolates isolated from sporadic food-poisoning cases in Korea with similar phenotypic and genotypic traits and biofilm-forming ability between emetic B. cereus isolates are not related to the genetic background for the toxin produced by B. cereus. In addition, it is worth noting that the rep-PCR assay tested in this study can be used to discriminate emetic strains from nonemetic isolates, even if not perfect. However, a much more detailed study on uncommon emetic strains or emetic strains harboring a high degree of genetic diversity may be necessary to evaluate the performance of the subtyping assay to discriminate between emetic and nonemetic strains of B. cereus.
Footnotes
Acknowledgment
This research was supported by Bio-industry Technology Development Program, Ministry of Agriculture, Food and Rural Affairs and the IT R&D program of MOTIE (Ministry of Trade, Industry, and Energy)/MISP (Ministry of Science, ICT, and Future Planning)/KEIT (Korea Evaluation Institute of Industrial Technology).
Disclosure Statement
No competing financial interests exist.