
Abstract
The emergence and spread of bacteriocin-resistant bacteria threaten the efficiency of bacteriocin usage as food preservatives. In this experiment, 19 selected Enterococcus durans strains acquired resistance after exposure to durancin GL, and the mutants had similar intermediate levels of resistance. One wild-type E. durans KLDS 6.0603 and its two resistant mutants, E. durans KLDS 6.0603-2 and E. durans KLDS 6.0603-3, were used to characterize phenotypic and genotypic differences. Approximately 100 μg/mL of durancin GL can penetrate the cytoplasmic membrane of E. durans KLDS 6.0603, causing damage to bacterial cells, but cannot penetrate E. durans KLDS 6.0603-2 and KLDS 6.0603-3 membranes. Unsaturated fatty acid content in resistant strains was significantly increased compared with wild-type strains, indicating that the former has more fluidity of cell membrane than the latter. Decreased mannose phosphotransferase system gene expression (mptD) was observed in the two resistant strains. Results showed that the factors, including the increased unsaturated fatty acid and decreased mptD expression, could contribute to durancin GL resistance.
Introduction
C
Enterococcus strains, usually found in human and animal digestive tracks, may either be harmless commensal microorganisms or opportunistic pathogens that act as causative agents for infectious diseases (Kayser, 2003; Lempiäinen et al., 2005; Golińska et al., 2013; Liu et al., 2013). Some Enterococcus strains even emerged as antibiotic-resistant pathogens in hospitals (Pusch et al., 2013; Shokoohizadeh et al., 2013; Kafil and Asgharzadeh, 2014; Jahan et al., 2015). Meanwhile, class IIa bacteriocins may possibly be used as food preservatives and to treat diseases caused by antibiotic-resistant pathogens (Yang et al., 2014). Regarding new antimicrobial agents, the potential risk of resistance development upon prolonged exposure should be determined to prove its safety (Gravesen et al., 2002a; Bastos et al., 2009, 2014). Bacterial resistance toward pediocin-like class IIa bacteriocins had been studied in L. monocytogenes strains previously (Dalet et al., 2001; Tessema et al., 2009; Opsata et al., 2010), still the cell membrane and gene expression alterations that are responsible for resistance in Enterococcus strains need to be further studied (Bastos et al., 2014).
Durancin GL is a novel class IIa bacteriocin produced by Enterococcus durans 41D, which was isolated from a Mexican-style artisanal cheese sample (Du et al., 2012a, 2012b). Durancin GL is active only against L. monocytogenes and some Enterococcus strains. The purpose of this study was to examine whether E. durans strains can acquire resistance after exposure to durancin GL and to characterize the phenotypic and genotypic differences between E. durans wild-type and resistant strains.
Materials and Methods
Bacterial strains
E. durans wild-type strains, KLDS 6.0603, 6.0315, 6.0314, 6.0318, 6.0607, 6.0605, and 6.0313, were obtained from Key Laboratory of Dairy Science (KLDS), Northeast Agricultural University, Ministry of Education. All the strains were isolated from traditional dairy products of Inner Mongolia and are sensitive to durancin GL. E. durans strains were cultured in M17 broth (Oxoid Ltd., Basingstoke, England).
Production of durancin GL
Durancin GL was synthesized (Wuhan Moon Bosciences Co. Ltd.) according to its amino acid sequence, which had been published in our previous article (Du et al., 2012b). The purity of the synthesized peptide was at least 95% and had a molecular mass of 4986 Da.
Bacteriocin antimicrobial activity assay
The antimicrobial activity of different concentrations of durancin GL solutions in sterilized ddH2O was examined using the spot-on-the-lawn antimicrobial method (Henderson et al., 1992; Du et al., 2012a). Approximately 5 μL of the solution was spotted on M17 agar plates that had been inoculated with an overnight culture of E. durans (0.5%, v/v). The plates were stored at 4°C for 2 h and then incubated for 12 h to 16 h at 37°C. The clear inhibition zone around each spot indicated antimicrobial activity.
Spontaneous mutation of E. durans strains with resistance to durancin GL
Spontaneous mutants of E. durans that are resistant to durancin GL were selected after a single exposure to durancin GL of 100 μg/mL. About two to four E. durans colonies grown in each clear zone of inhibition were separately further inoculated into M17 broth with 100 μg/mL durancin GL added to obtain the durancin GL-resistant strains. The selected durancin GL-resistant strains were named by using the suffixes following the wild-type strain designations.
RAPD-PCR analysis
The wild-type E. durans strains and their corresponding mutants were cultured in 5 mL of M17 broth at 37°C for 16 h. The genomic DNA of each strain was then separately extracted using a TIANamp bacteria genomic DNA extraction kit (Tiangen) according to the manufacturer's procedure. Amplification reactions were performed in a 25 μL reaction volume containing 50 ng of bacterial total DNA genome, 2 μL of dNTP, 0.5 μL of PrimeSTAR HS DNA polymerase (Takara), 1 μL of primer D8635 (5′-GAG CGG CCA AAG GGA GCA GAC-3′, 10 μm), 5 μL of 5× polymerase chain reaction (PCR) buffer (Mg2+-free), and 1.5 μL of MgCl2 (25 mM). The reaction was performed by a Thermal Cycler Dice (Takara) under a 94°C temperature profile for 2 min. This reaction was followed by 35 cycles at 94°C for 1 min; 42°C for 1 min; 72°C for 1.5 min; and 72°C for 10 min during the final stage (Akopyanz et al., 1992; Suzzi et al., 2000). Amplification products were separated by electrophoresis on 2% (w/v) agarose gels in 1% TAE buffer.
Time-kill assay
Bacterial growth inhibition was tested by inoculating ca. 104 CFUs of E. durans cells into 20 mL of M17 broth with various concentrations (0, 0.01, 0.10, 0.39, 12.5, 25, and 100 μg/mL) of durancin GL. The cultures were incubated for 24 h with the OD600 value determined every 2 h.
Membrane integrity assessment
Membrane integrity was assessed using the Dead/Live BacLight™ kit (Invitrogen) according to a previous study (Liu et al., 2015). E. durans strains of KLDS 6.0603, 6.0603-2, and 6.0603-3 were cultured to the early logarithmic phase (OD 660 = 0.6) in 20 mL of M17 broth at 37°C. The bacterial cells were harvested by centrifugation (5000 g at 4°C for 10 min) and washed with saline solution (9 g/L NaCl), respectively. The bacterial cells were resuspended in saline solution with an OD600 value of 0.6. This bacterial suspension was divided into three portions. Antimicrobials were added to two of the portions at the concentrations of 10 μg/mL durancin GL and 125 μg/mL nisin, respectively. The remaining portion was left untreated as a control. The resuspension was incubated at 25°C for 60 min. Two fluorescent probes, that is, SYTO-9 and propidium iodide (PI), were added to the treated cells at final concentrations of 5 and 30 μM, respectively. The mixtures were incubated in the dark for 15 min at 25°C. Afterward, 3 μL of stained cells was spotted on a microscope slide and covered with a glass coverslip. Digital images were obtained from a fluorescence microscope (DFM-20C; Zeiss) by using the following settings: excitation/emission of 480/500 and 490/635 nm for SYTO-9 and PI, respectively.
Examination of cell morphology by scanning electron microscope
E. durans strains of KLDS 6.0603, 6.0603-2, and 6.0603-3 were cultured to the early logarithmic phase (OD 660 = 0.6) in 20 mL of M17 broth at 37°C. The cells were harvested by centrifugation at 5000 g for 10 min and resuspended in 10 mL saline solution (9 g/L NaCl). The resuspension of each strain was divided into four portions. Antimicrobials were added to three of the portions at the following concentrations: 100 μg/mL durancin GL, 10 μg/mL durancin GL, and 125 μg/mL nisin. The remaining portion was left untreated as a control. The resuspension was incubated at 37°C for 2 h, and subsequently, the cells from all four tubes were harvested through centrifugation at 5000 g for 10 min and washed with 0.1 M phosphate buffer. The following treatment was carried out according to the previous article (Liu et al., 2015). Digital images of the treated and untreated E. durans cells were acquired through scanning electron microscopy (SEM; EVOMA10/LS10; Zeiss) at an accelerating voltage of 15 kV and instrumental magnifications of ×10,000.
Fatty acid analysis
Total bacterial lipids were extracted according to the previous method with small modifications (Folch et al., 1957). E. durans strains of KLDS 6.0603, 6.0603-2, and 6.0603-3 were cultured to the early logarithmic phase (OD 660 = 0.6) in 1000 mL M17 broth at 37°C. The cells were harvested by centrifugation at 5000 g for 10 min. Then, 3.0 g of bacterial pellets was added to 18 mL of chloroform/methanol (2/1, v/v) solution at 1500 rpm by using an Ultra Turrax. The homogenate was allowed to stand for 1 h before passing through two layers of filters. A solution, 0.2-fold of its volume, containing 7.3 g/L NaCl and 0.5 g/L CaCl2, was added to the filtrate. The mixture was centrifuged for 15 min at 3000 rpm (Allegra 64R; Beckman), and the lower phase was dried under vacuum on a rotary evaporator (RE-85C; Yarong) in a 44°C water bath and stored at −40°C afterward. Methylated fatty acids were analyzed using a gas chromatograph (GC-2010 Plus; Shimadzu) equipped with a flame ionization detector and a split injector according to the previous article (Gandemer, 2002). The relative percentages of fatty acids were determined by the peak areas.
RNA preparation and Quantitative reverse transcription polymerase chain reaction analysis of gene expression
To prepare total cellular RNA, 5 mL of the E. durans cultures was grown to the early log phase (OD600 = 0.6). The cells were harvested from 3 mL cultures by centrifugation under 4°C for 3 min at 10,000 g. RNA was extracted using the TIANamp RNAprep pure Cell/Bacteria Kit (Tiangen). RNA integrity, concentration, and purity were checked by electrophoresis on a 2% (w/v) agarose gel and by measuring the OD260/OD280 ratio. Contaminated DNA removal and cDNA synthesis were carried out using the PrimeScript® RT reagent Kit with gDNA Eraser (Takara) following the manufacturer's recommendations.
Gene expression was analyzed by real-time PCR according to the previous article (Hew et al., 2007). The primers used for the analysis of gene expression were designed using Primer5.0 software based on the genome sequences of E. durans 41D (not published) and are listed in Table 1. The gdhA gene (Table 1) was used as reference for data normalization. Gdh1 and gdh2 housekeeping genes (Table 1) were also included as control samples to ensure data reliability. cDNA quantification was conducted using the SYBR Green PCR master mix (Applied Biosystems) in a 7500 Fast Real-Time PCR System (Applied Biosystems). The reactions consisted of the following components: 12.5 μL of SYBR Premix Ex Taq™ II (Takara), 8.5 μL of ddH2O, 2 μL of primer mix (containing 5 pmol of each primer), and 2 μL of each cDNA sample. The amplification program was set at 95°C for 30 s, followed by 40 cycles at 95°C for 5 s and 60°C for 30 s. All the samples, including no-RT and no-template controls, were analyzed in triplicate. Data analysis was performed using the 2−ΔΔCT method, where ΔΔCT = ΔCT (treated sample) − ΔCT (untreated sample), ΔCT = CT (target gene) − CT (gdhA), and CT is the threshold cycle value for the amplified gene. A fold change of <−2 or >2 was considered a real difference in the transcript level.
PTS, phosphotransferase; RT-qPCR, Quantitative reverse transcription polymerase chain reaction.
Statistical analysis of the data
The differences between groups were evaluated by one-way analysis of variance, using SPSS software and the Pearson correlation coefficient option (version 18.0; IBM-SPSS, Inc.). Differences were regarded as significant at p < 0.05. All data are expressed as mean ± standard error.
Results
Development of durancin GL resistance
The antimicrobial activities of durancin GL for E. durans wild-type strains and spontaneous mutants are shown in Table 2. The natural minimum inhibitory concentration of durancin GL for all the E. durans wild-type strains used in this study were about 10 μg/mL. While the durancin GL-resistant strains can be resistant to 100 μg/mL durancin GL, they are not resistant to 1 mg/mL durancin GL. The resistant mutants have the same RAPD profiles as the wild-type strains from which they were derived (Fig. 1).
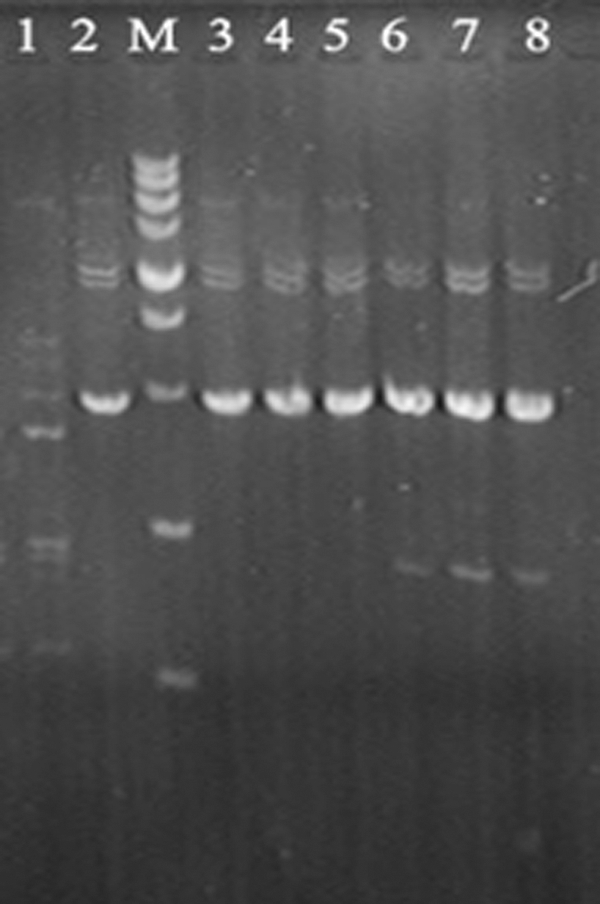
RAPD profiles of Enterococcus strains. 1, Enterococcus faecalis R612-Z1; 2, Enterococcus durans KLDS 6.0603; M, DNA marker with DNA fragments ranging from 100 to 5000 bp (5000\3000\2000\1500\1000\750\500\250\100 bp); 3, E. durans KLDS 6.0603; 4, E. durans KLDS 6.0603-2; 5, E. durans KLDS 6.0603-3; 6, E. durans KLDS 6.0315; 7, E. durans KLDS 6.0315-1; 8, E. durans KLDS 6.0315-2.
“+” means bacterial inhibition; “−” means bacterial growth.
Bacterial growth in liquid culture
Although the growth of E. durans KLDS 6.0603 at the early growth phase was inhibited by durancin GL in the concentrations of 0.10, 0.39, 12.5, 25, and 100 μg/mL compared with the control group, the bacteria continued to grow after 8 h. The final OD values of the treated group were the same with the control group. The final E. durans strains grown in the broth with durancin GL were resistant to durancin GL, indicating that these strains became durancin GL resistant. However, the growth of E. durans KLDS 6.0603-2 in the broth was not affected by adding durancin GL at these concentrations.
Effects of antimicrobials on the cell morphology of wild-type and resistant strains
Changes in the cell morphology of E. durans wild-type KLDS 6.0603 and two resistant strains were further examined by SEM. The bacterial cells of KLDS 6.0603-2 and 6.0603-3 had irregular cell surfaces and were larger than those of control KLDS 6.0603 (Fig. 2A, E, I). In contrast to control samples, marked changes were evident on the outer membranes of KLDS 6.0603 and resistant strains treated with nisin (Fig. 2B, 2F, 2). However, durancin GL can only cause extensive damage to the bacterial cells of KLDS 6.0603 (Fig. 2C, D, G, H, K, L) after 2 h of treatment.
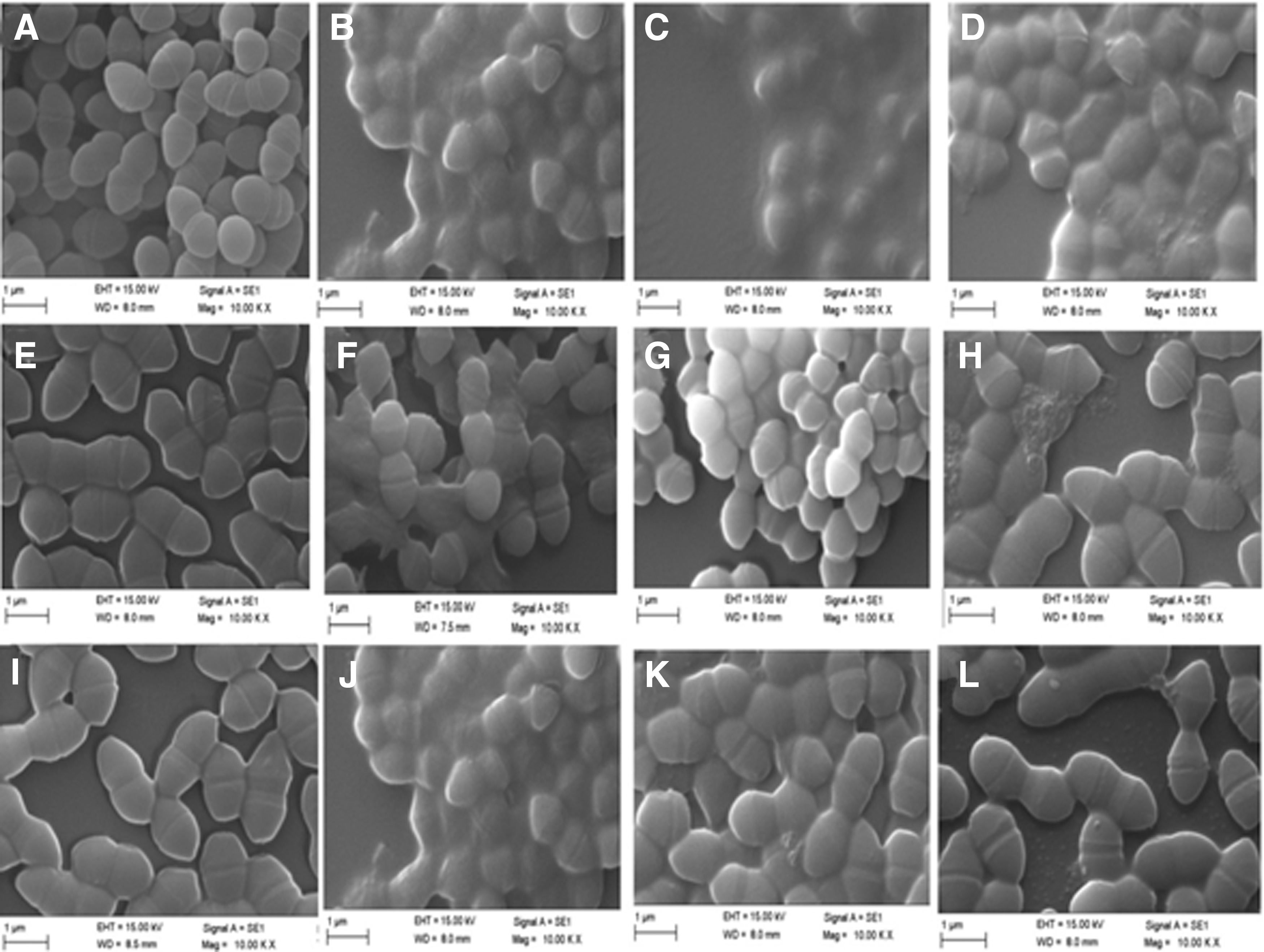
Scanning electron micrographs of E. durans strains after durancin GL and nisin treatment for 2 h.
Effects of antimicrobials on cell membrane permeability of wild-type and resistant strains
Cytoplasmic membrane permeability was observed after being stained with two fluorescent nucleic acid stains, SYTO-9 and PI. The untreated bacterial cells of KLDS 6.0603, 6.0603-2, and 6.0603-3 were stained green (Fig. 3A, D, G). The majority of bacterial cells of KLDS 6.0603 treated by durancin GL for 60 min became red (Fig. 3B), whereas the bacterial cells of KLDS 6.0603-2 and 6.0603-3 treated by durancin GL for 60 min remained green (Fig. 3E, H). These results show that durancin GL can penetrate the cytoplasmic membrane of KLDS 6.0603, but cannot penetrate the cytoplasmic membranes of KLDS 6.0603-2 and 6.0603-3. The bacterial cells of KLDS 6.0603, 6.0603-2, and 6.0603-3 treated by nisin for 60 min became red (Fig. 3C, F, I), indicating that nisin can penetrate all the cytoplasmic membranes of KLDS 6.0603, 6.0603-2, and 6.0603-3.
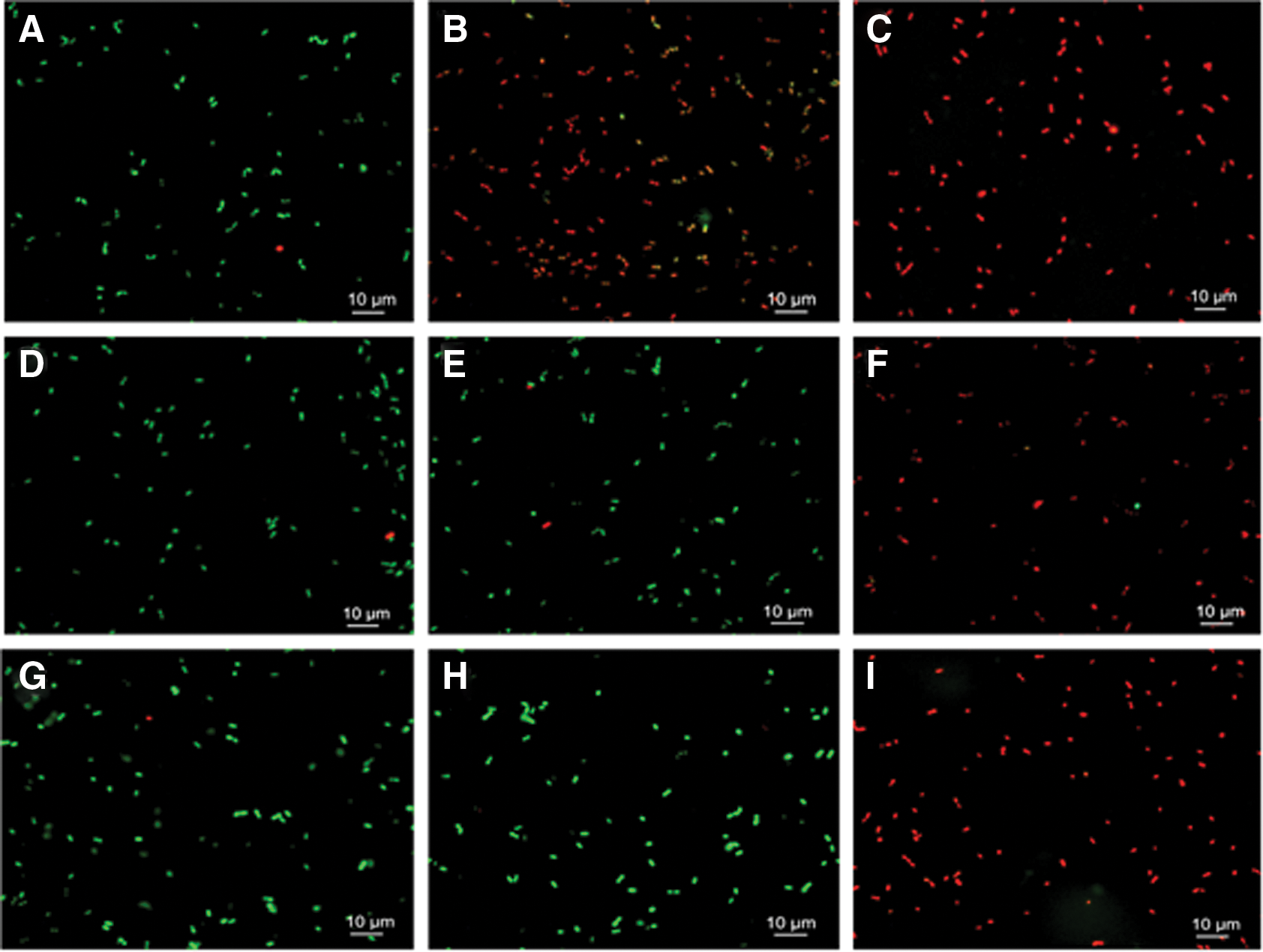
Changes in bacterial cell membrane permeability after durancin GL and nisin treatment for 60 min. Intact membranes exhibit green fluorescence, and the compromised membranes exhibit red fluorescence.
Fatty acid analysis
The relative percentages of fatty acids of KLDS 6.0603, 6.0603-2, and 6.0603-3 are shown in Table 3. The content of monounsaturated fatty acid and polyunsaturated fatty acid in the two resistant strains was significantly increased compared with the wild-type strain KLDS 6.0603.
Experiments were conducted in triplicate and their means ± SDs are presented. Different lowercase letters in a row indicate a significant difference (p < 0.05).
FA, fatty acid; MUFA, monounsaturated fatty acid; PUFA, Polyunsaturated fatty acid; SFA, saturated fatty acid.
Quantitative reverse transcription polymerase chain reaction
Based on the sequenced genome sequence of E. durans (date had not been published), two gene clusters encoding the function of mannose/fructose/sorbose-specific PTS system were predicated. However, which Man-PTS system has the docking molecule of durancin GL had not been identified yet. IID component genes corresponding to these two Man-PTS systems were designated as mptD18 and mptD23. Both the expression levels of the mptD18 and mptD23 genes in the two resistant strains of KLDS 6.0603-2 and 6.0603-3 were downregulated about twofold compared with the expression levels in the corresponding wild-type strain (Fig. 4).
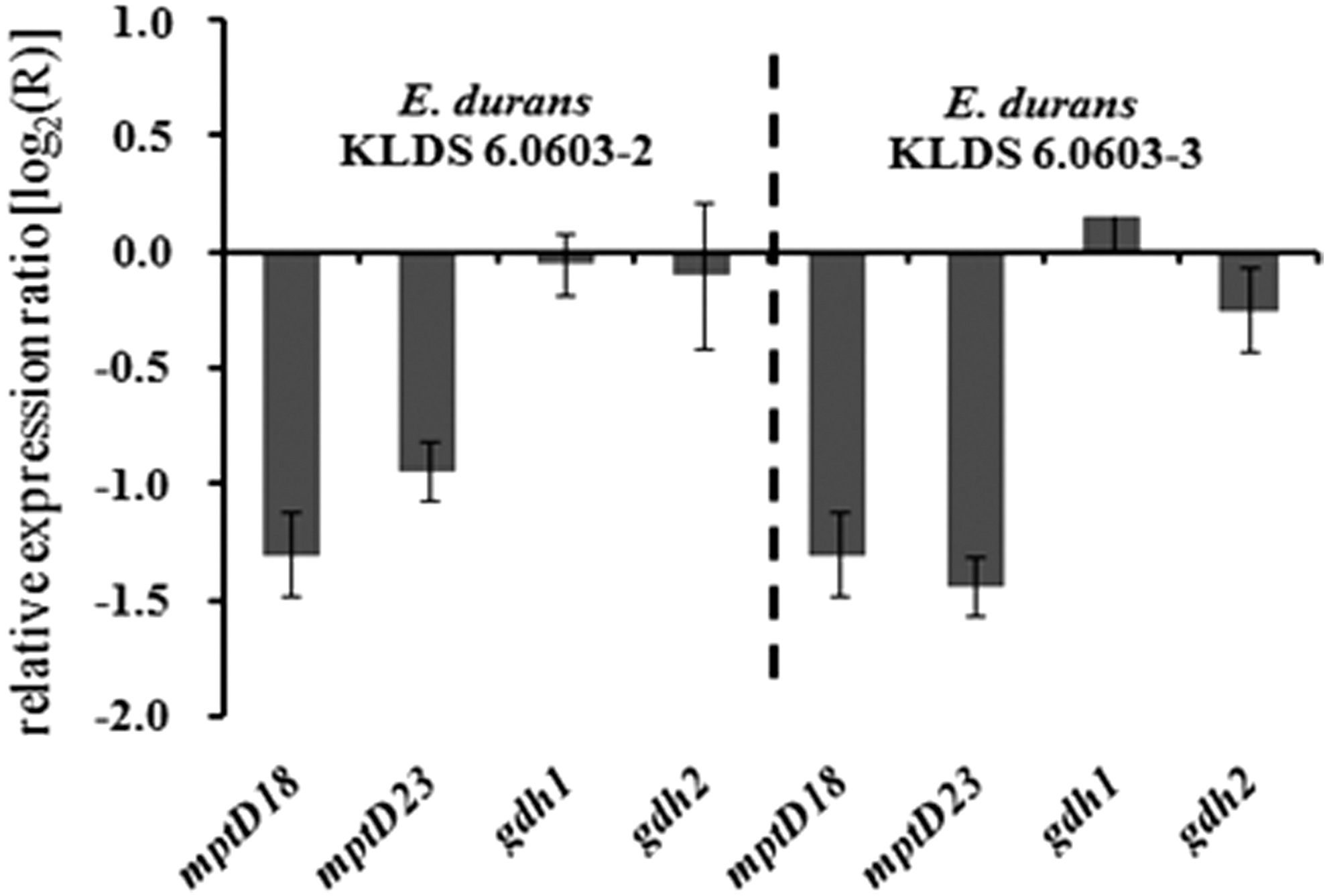
Changes in mptD gene expression in durancin GL-resistant strains of E. durans KLDS 6.0603-2 and KLDS 6.0603-3 compared with that in the wild-type strain of E. durans KLDS 6.0603. The 0-line represents the expression level of E. durans KLDS 6.0603.
Discussion
The emergence and spread of bacteriocin resistance among food spoilage and pathogenic bacteria threaten the safety and availability of using bacteriocins as food preservatives and new antibiotic substances (Vignolo et al., 2000). In the present study, the spontaneous mutants of E. durans existed in clear inhibition zones indicating that this was a consistent phenomenon for E. durans strains to acquire resistance. Investigation of the physiological characteristics of bacteriocin-resistant strains should help us to understand the relationship between bacteriocin and its target bacteria. Tessema et al. (2009) grouped the resistant mutants of L. monocytogenes to class IIa bacteriocins into mutants with high level of resistance (500 times to 106 times more resistant than the wild-type strain) and mutants with intermediate levels of resistance (2–8 times more resistant than the wild-type strain). According to these criteria, our isolated resistant strains were intermediate levels of resistance. Different mechanisms are associated with resistance development to class IIa bacteriocins. In many cases, high-level resistance of Gram-positive bacteria to class IIa bacteriocins results from either loss or reduction of expression of Man-PTS (Héchard et al., 2001; Gravesen et al., 2002b; Tessema et al., 2009; Opsata et al., 2010; Kjos et al., 2011). Vadyvaloo et al. (2004) reported that the transcription levels of the mptA gene in the highly resistant spontaneous mutants, B73-MR1 and 412P, were decreased more than 1000-fold than the wild-type strains. Although the expression levels of Man-PTS in intermediate resistant strains to class IIa bacteriocins were also decreased, this decrease is only approximately fourfold (Vadyvaloo et al., 2004; Tessema et al., 2009). A correlation between transcription levels of the mpt gene and the intermediate levels of resistance was also found in our study.
Although the downregulation of Man-PTS expression is the main resistance mechanism, other mechanisms may be also involved in mutants with resistance to various class IIa bacteriocins (Ran et al., 2013; Roces et al., 2012). A significant increase in unsaturated fatty acids was also found in the resistant strains, which is an indication of more fluidity of cell membrane, and the increase of unsaturated fatty acids may prevent the oligomerization of the bacteriocin monomers to make a stable pore in the phospholipid bilayer (Limonet et al., 2002). Meanwhile, a decrease in cell wall negative charge caused by an increase in D-Ala content in teichoic acids was found to be responsible for the resistance to class IIa bacteriocins (Vadyvaloo et al., 2004). Since the antibacterial mechanism of nisin is different from that of Class IIa bacteriocin, the resistant strains with the increased unsaturated fatty acid and decreased mptD expression in this study were still sensitive to nisin.
Conclusions
E. durans strains can acquire resistance to durancin GL, and multiple factors, including the increased unsaturated fatty acid content and decreased mptD expression level, could contribute to durancin GL resistance.
Footnotes
Acknowledgments
The authors thank Dr. Guicheng Huo from KLDS for providing some of the Enterococcus strains used in this study. This study was funded by the National Key Technology Support Program (2015BAD16B04), the National Natural Science Foundation of China (31271930), the Jiangsu Province Science and Technology Support Program (BE2014711), and a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Disclosure Statement
No competing financial interests exist.