
Abstract
The objectives of this study were to characterize the phenotype and genotype of 36 non-O157 Shiga toxin–producing Escherichia coli (STEC) strains isolated from humans, ovines, or bovines, including the top 6 (O26, O45, O103, O111, O121, and O145) and three other serogroups implicated in serious illness (O91, O113, and O128). Biofilms were formed by all strains with intermediate to strong biofilm producers (n = 24) more common at 22°C than at 37°C (p < 0.001) and 48 and 72 h (p < 0.001) than 24 h of incubation time. Biofilm-forming potential differed by serogroup and origin with O113 and human strains exhibiting the highest potential (p < 0.001). Biofilm-associated genes, csgA/csgD/crl/fimH (100%), flu (94%), rpoS (92%), ehaAα (89%), and cah (72%), were most prevalent, while mlrA (22%) and ehaAβ (14%) were least prevalent, although there was no clear compliment of genes associated with strains exhibiting the greatest biofilm-forming capacity. Among 12 virulence genes screened, iha and ehxA were present in 92% of the strains. The occurrence of stx1 in the top 6 serogroups (8/12, 67%) did not differ (p = 0.8) from other serogroups (17/24, 71%), but stx2 was less likely (confidence interval [CI] = 0.14–1.12; p = 0.04) to be in the former (9/24, 38%) than the latter (9/12, 75%). Excluding serogroups, O91 and O121, at least one strain per serogroup was resistant to between three and six antimicrobials. Streptomycin (31%), sulfisoxazole (31%), and tetracycline (25%) resistance was most common and was 35–50% less likely (p < 0.05) in human than animal strains. All non-O157 STEC strains were able to form biofilms on an abiotic surface, with some exhibiting resistance to multiple antimicrobials. Potential as a reservoir of antimicrobial resistance genes may be another hazard of biofilms in food-processing plants. As a result, future strategies to control these pathogens may include measures to prevent biofilms.
Introduction
S
Non-O157 STEC outbreaks are typically associated with contaminated foods of bovine origin (Mathusa et al., 2010; Robbins et al., 2014) or vegetable products contaminated with bovine feces (Hussein and Bollinger, 2005; Smith et al., 2014). Specifically, six serogroups, O26, O45, O103, O111, O121, and O145 (top 6), were responsible for ∼70% of non-O157 infection from 1983 to 2002 in the United States (Brooks et al., 2005) and were declared as adulterants in raw beef products in the United States (USDA, 2012). Although other non-O157 serogroups such as O91, O113, and O128 are less likely to be associated with outbreaks, they can cause severe illness (Johnson et al., 2006; Bettelheim, 2007). The production of Shiga toxins (Stx) (Paton and Paton, 1998) and possession of mobile genetic elements (MGEs) (Tatsuno et al., 2001; Imamovic et al., 2010; Etcheverria and Padola, 2013) are the main virulence features of STEC-associated pathogenesis. Moreover, genetic diversity in STEC virulence varies among strains (Anjum et al., 2014; Tseng et al., 2014) and geographical locations (Mellor et al., 2013).
STEC may form biofilms on food or food contact surfaces (Silagyi et al., 2009; Dourou et al., 2011; Wang et al., 2012). Decontaminating food-processing equipment of biofilms is particularly difficult as biofilms frequently slough off, releasing cells into food products (Vogeleer et al., 2014). Biofilms can also form on food equipment surfaces, reducing the effectiveness of disinfectants (Vogeleer et al., 2014). Most STEC biofilms studies have focused on O157:H7 (Dourou et al., 2011; Uhlich et al., 2013; Wang et al., 2014) and few report biofilm formation by non-O157 STEC strains (Chen et al., 2013; Uhlich et al., 2014).
Differing evolutionarily from O157, non-O157 STEC are heterogeneous and information on their virulence, fitness, and stress responses is limited (Wang et al., 2013). This lack of knowledge and strain heterogeneity increase complexity of developing strategies to reduce food and water contamination with these pathogens. Therefore, the aim of this study was to investigate molecular and phenotypic features of non-O157 STEC strains from nine major serogroups isolated from Canada. Strains were characterized for the presence of biofilm-forming- and virulence-associated genes, biofilm-forming ability, and antimicrobial resistance (AMR) profiles.
Materials and Methods
Bacterial strains and growth conditions
All strains used (Table 1 and Supplementary Table S1; Supplementary Data are available online at
Non-O157 STEC strains isolated from a bovine (B) and human (H).
Biofilm-forming ability was scored based on biofilm formation under optimal experimental conditions: +, weak; ++, intermediate; +++, strong.
Intact indicates detection of amplicon A/B only; Variant-L indicates detection of amplicons A/B and A/E; Variant-R indicates detection of amplicons A/B and B/F.
STEC, Shiga toxin–producing Escherichia coli.
Biofilm formation assay
Biofilm formation was assessed in 96-well polystyrene microplates (Nunc, Edmonton, AB) using a modification of Uhlich et al. (2013). Briefly, LB broth with no salt (LB-NS) only (negative control, 200 μL) or LB-NS diluted overnight culture of each strain (200 μL) was dispensed into four replicate wells of a microplate. The plates were incubated at 22°C or 37°C for 24, 48, or 72 h. Following incubation, supernatants were removed and wells were washed thrice. Remaining attached bacteria were fixed with 250 μL of absolute methanol (Sigma-Aldrich, Okaville, ON) per well for 15 min. Plates with biofilms were then emptied, air-dried, and stained with 1% (w/v) crystal violet (Sigma-Aldrich) solution for 15 min, followed by three water washes and air-drying. The dye bound to the biofilm was then dissolved with 33% glacial acetic acid (200 μL; Sigma-Aldrich) per well and OD590nm values were measured.
Based on the OD590nm produced by biofilms, strains were classified as no biofilm, weak, intermediate, or strong biofilm producers, as previously described (Stepanovic et al., 2000). Briefly, cutoff optical density value (ODc) of 0.071 was three standard deviations above mean OD of negative controls. Strains were classified as OD ≤ ODc, no biofilm producer; ODc < OD ≤2 × ODc, weak biofilm producers; 2 × ODc < OD ≤4 × ODc, intermediate biofilm producers; and 4 × ODc < OD, strong biofilm producers.
Polymerase chain reaction assay
All strains were screened by polymerase chain reaction (PCR) for the presence of major genes associated with biofilm formation and virulence (Tables 1 and 2). Bacterial DNA was isolated from 18-h cultures using the NucleoSpin® Tissue Kit (Macherey-Nagel, Bethlehem, PA). All PCR assays excepting bacteriophage insertions in mlrA were conducted individually using HotStar Plus MasterMix (Qiagen, Mississauga, ON) and 0.2 μM of each primer at annealing temperatures as indicated (Supplementary Table S2). The bacteriophage insertions in mlrA (yehV) were investigated using multiplex PCR and primers (Supplementary Table S2) complementary to regions flanking bacteriophage insertion site in mlrA (primers A and B) and to ends of predominant bacteriophage occupying mlrA insertion site (primers E and F) (Shaikh and Tarr, 2003). Multiplex PCR assays used QuantiFast Multiplex Master Mix (Qiagen) and 0.5 μM of each primer at annealing temperature of 57°C. Each PCR contained positive and negative controls. Samples were then electrophoresed on 2% agarose gels (w/v) stained with ethidium bromide and visualized with a UV transilluminator (Alpha Innotech, San Leandro, CA).
PCR, polymerase chain reaction; STEC, Shiga toxin–producing E. coli.
Antimicrobial resistance
AMR was determined against 12 antimicrobials using the disc diffusion method (CLSI, 2014). Antimicrobial discs (BD, Mississauga, ON) were used: ampicillin (AMP; 10 μg), amoxicillin–clavulanate (AMC; 20/10 μg), ceftazidime (CAZ; 30 μg), tetracycline (TET; 30 μg), kanamycin (KAN; 30 μg), nalidixic acid (NAL; 30 μg), streptomycin (STR; 10 μg), chloramphenicol (CHL; 30 μg), neomycin (NEO; 30 μg), enrofloxacin (ENR; 5 μg), trimethoprim–sulfamethoxazole (TMS; 1.25/23.75 μg), and sulfisoxazole (SUL; 250 μg). Inoculum of each strain was streaked on Mueller–Hinton agar (Dalynn Biologicals, Calgary, AB), and the appropriate drug-impregnated discs were placed on the agar surface. Plates were inverted and incubated (37°C, 18 h), and zones of inhibition measured.
Statistical analysis
Results from biofilm formation were compiled from two independent experiments. No biofilm and weak biofilm formation were scored as negative (designated as low biofilm-forming potential) and intermediate or strong biofilm formation (designated as high biofilm-forming potential) was scored as positive. Influence of origin (animal vs. human), serogroups, incubation temperature, and time on biofilm-forming potential was analyzed using GLIMMIX with random measures. Odds ratios were calculated for the percentage of high biofilm-forming potential with cohorts of animal origin, serogroup O26, at 37°C and 24 h showing a low percentage of high biofilm-forming ability as referent. Within each serogroup, OD590nm values were transformed and then analyzed using MIXED and least-squares differentiated means (p < 0.05). Correlations between biofilm-forming potential and biofilm-forming genes and between serogroup or origin and virulence-associated genes were assessed using tetrachoric correlation and Cochran–Mantel–Haenszel statistics of FREQ. All analyses were conducted with SAS (version 9.3; SAS Institute, Cary, NC).
Results
Biofilm formation
Generally, biofilm-forming potential differed by serogroup and depended on incubation temperature and time (p < 0.001) with human strains nearly twice as likely (p < 0.001) to form strong biofilms than animal strains (Table 3). Among serogroups, O113 exhibited the overall highest biofilm-forming potential (p < 0.001), followed by O91 (p < 0.01), O103 and O121 (p < 0.05), and O45 and O128 (p < 0.05), respectively. Strains of O111, O145, and O26 formed the least biofilms. Across all serogroups, intermediate to strong biofilms were more likely at 22°C than 37°C (p < 0.001) and with 48 and 72 h (p < 0.001) than 24-h incubation times. Bofilms developed at 37°C, but became less established with longer incubation times in some O45, O91, O113, and O128 strains.
Odds ratio for serogroups, temperature, time, and origin were generated, respectively, with cohorts of serogroup O26, at 37°C, 24 h, and animal origin as the referent.
CI, confidence interval.
Although the ability to form biofilms varied among strains within serogroup (p < 0.001; Fig. 1), all strains produced biofilms under some conditions with 24/36 strains showing intermediate or strong biofilm (Table 1).
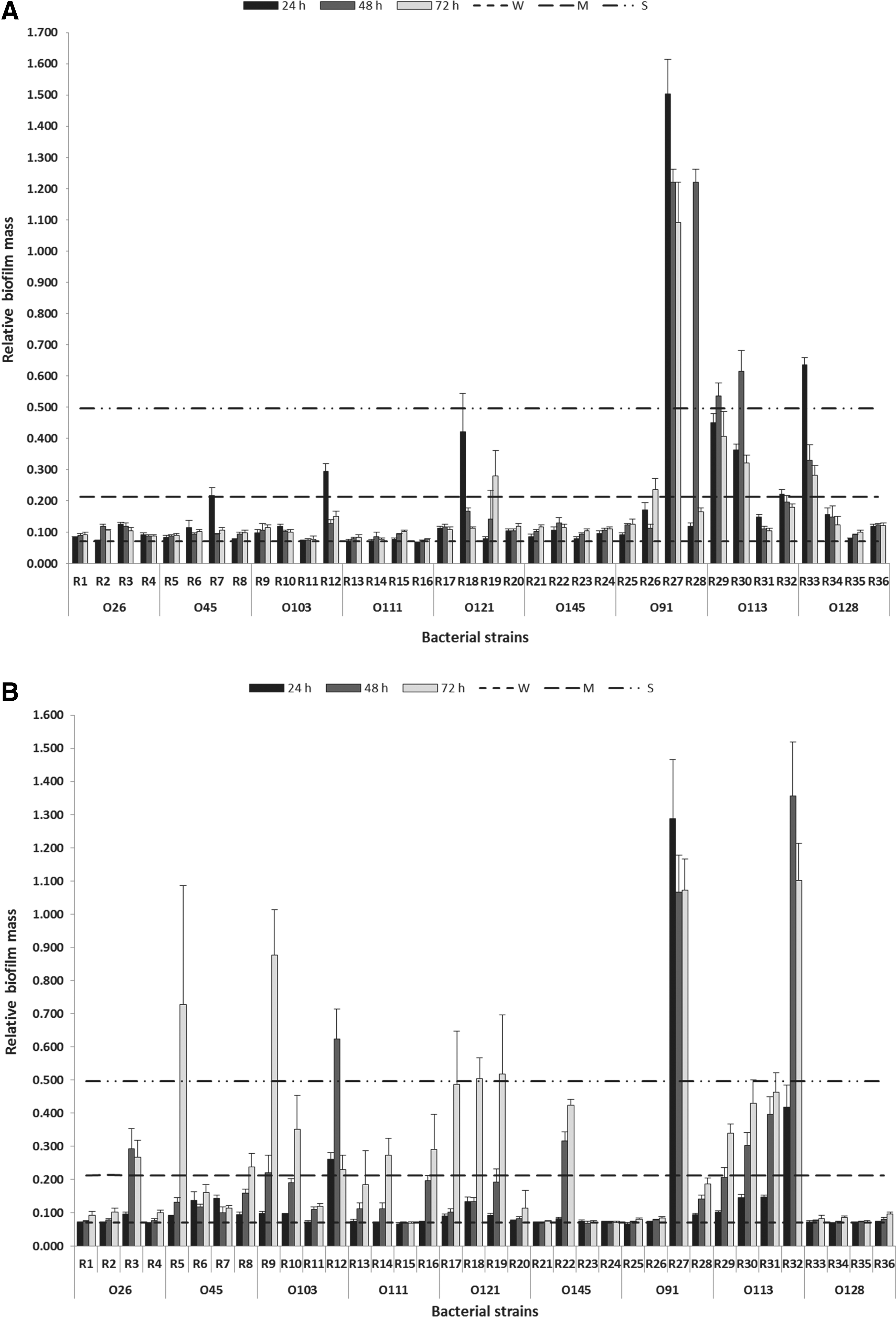
Biofilm formation of non-O157 STEC on polystyrene surface at 37°C
Detection of biofilm-associated genes
Biofilm-associated genes, csgA/csgD/crl/fimH (100%), flu (94%), rpoS (92%), ehaAα (89%), and cah (72%), were most prevalent, while mlrA (22%) and ehaAβ (14%) were least prevalent (Table 1), although no correlation (p > 0.1) was found between biofilm-forming ability and the presence of these genes. Most strains (33/36, 92%) carried the rpoS gene, but this gene was lacking in R29, R30, and R35. Of 36 strains, mlrA-intact (amplicon A/B only) was detected in 28 (78%), indicating that these strains did not carry the prophage stx1 in mlrA.
Detection of virulence-associated genes
Prevalence of virulence genes in the non-O157 STEC strains ranked as iha/ehxA (92%), ureA (81%), adfO (78%), efa1-5′ (72%), stx1 /eae (70%), efa1-3′ (67%), sodCF (58%), stx2 /toxB (50%), saa (14%), and tccP (8%; Table 2). Of the strains, 18 contained only stx1 , 11 were only positive for stx2 , and 7 carried both stx genes. Prevalence of stx1 from the top 6 serogroups (8/12, 67%) did not differ (p = 0.8) from that of other serogroups (17/24, 71%), but detection of stx2 was 40% less likely (confidence interval [CI] = 0.14–1.12; p = 0.04) in the top 6 (9/24, 38%) than in the latter (9/12, 75%).
Other genes associated with virulence, including eae, adfO, efa1-5′, efa1-3′, toxB, ureA, and sodCF, were more (p < 0.05) common in top 6 strains than other serogroups. All strains from the top 6 were positive for eae, adfO, and efa1-5′, whereas these genes were only present in 8–33% of the 12 strains from O91, O113, and O128. In contrast to the top 6 strains, O91, O113, and O128 were negative for both toxB and tccP. Additionally, most O91, O112, and O128 strains lacked an intact efa1 gene, even though four strains possessed either efa1-5′ or efa1-3′. A lower prevalence of ureA (50%) was also observed in O91, O113, and O128 serogroups than in the top 6 serogroups (96%). No correlation (p > 0.1) was found between any virulence genes and strain origin.
Antimicrobial susceptibility
Excluding serogroups, O91 and O121, 12 strains from other serogroups were resistant to three to six antimicrobials and concurrent STR-SUL-TET resistance was most common (8/12, 67%; Table 4). Moreover, STR (31%), SUL (31%), and TET (25%) resistance was 35–50% less likely (p < 0.05) to be detected from strains with human origin than those from bovine origin. Of the 12 multidrug-resistant (MDR) strains, 8 possessed an intermediate or strong ability to form a biofilm, and all strains but one (R21, O145:NM) possessed stx1 or both stx genes (Table 4). In addition, all strains tested were susceptible to AMC, CAZ, CHL, ENR, and NAL.
Non-O157 STEC strains isolated from a bovine (B) and human (H).
Biofilm-forming ability was scored based on biofilm formation under optimal experimental condition: +, weak; ++, intermediate; +++, strong.
AMP, ampicillin; KAN, kanamycin; NEO, neomycin; STEC, Shiga toxin–producing E. coli; STR, streptomycin; SUL, sulfisoxazole; TET, tetracycline; TMS, trimethoprim–sulfamethoxazole.
Discussion
This study was the first to evaluate potential molecular and biological hazards of STEC strains representing nine important non-O157 serogroups from Canada.
Biofilm formation and its associated genes
All non-O157 STEC strains were able to form biofilms with the majority showing high biofilm-forming potential, although there was variation in biofilm formation among serogroups. This variation may be attributed to the relatively limited number of strains studied, but biofilm formation of STEC O157, O26, O91, O103, O111, and O113 on polystyrene surfaces has been recognized to be strain dependent (Biscola et al., 2011; Wang et al., 2012). In the present study, all strains of serogroup O113 readily formed biofilms. Across all serogroups and times, biofilm formation on polystyrene surfaces occurred most readily at room temperature. This agrees with the observation that 19 non-O157 STEC strains from O26, O45, O103, O111, O113, O121, and O145 formed more extensive biofilms at 25°C than at 37°C (Uhlich et al., 2014). Similar results were also obtained by Nesse et al. (2014) as strains of O103:H2 produced less biofilm at 37°C than at 20°C. Additionally, biofilm mass was generally increased with exposure time, confirming the results of Fouladkhah et al. (2013) where biofilm formation was more extensive after 7 days compared with on day 0 at 15°C and 25°C. No reports, to our knowledge, have compared the biofilm-forming ability between human and animal strains of STEC, but Vijay et al. (2015) found that human isolates of Enteroaggregative E. coli produce comparatively more biofilm than did animal isolates.
To identify biofilm-associated genes, a panel of adhesin genes and autotransporter protein-associated genes was screened by PCR. For STEC, biofilm formation in most strains depended on the expression of curli fimbriae (Biscola et al., 2011; Wang et al., 2012). In the present study, type 1 fimbriae-encoding gene fimH, curli gene csgA, and crl, as well as central biofilm-regulating gene csgD, were present in all strains examined. However, biofilm formation potential of these strains was highly variable, likely reflecting divergence in the expression of these genes.
Previous studies have shown that variations in rpoS and prophage insertions in mlrA limited CsgD-dependent biofilm formation in O157:H7 and non-O157 strains (Chen et al., 2013; Uhlich et al., 2013). Even though mutations of the rpos were not assessed in the present study, two of three non-O157 strains lacked the rpoS, but surprisingly, still exhibited high biofilm-forming capacity. To date, frequency of RpoS in STEC strains is unknown, but Franz et al. (2015) reported that two strains from O128:H2 and O107:H2 were rpoS negative. Presumably, another sigma factor gene may play an rpoS-like role in regulating the transcription of csgD in response to environmental stress. In addition, fewer non-O157 strains studied harbored prophage stx1 in mlrA, consistent with the previous findings that mlrA prophage insertions in strains of non-O157 serogroups are not as common as in O157:H7 strains (Shaikh and Tarr, 2003; Chen et al., 2013). Interestingly, stx1 was detected in 25 strains, suggesting that stx1 -encoding phages may have a different chromosomal insertion site in some non-O157 STEC strains (Shaikh and Tarr, 2003; Kondo et al., 2010).
Autotransporters are other important factors for biofilm formation of non-O157 STEC (Jaglic et al., 2014). In the present study, autotransporter protein-associated genes, cah and flu, were detected in most non-O157 STEC strains. It is interesting that among the strains lacking cah in the present study, 60% exhibited reduced biofilm formation with increasing incubation time at 37°C. As the presence of none of the genes was statistically related to biofilm formation, further investigation of expression of these genes is required to determine their relative importance in biofilm formation.
Correlation among virulence determinants and non-O157 STEC strains
Karmali et al. (2003) proposed grouping STEC strains into five seropathotypes (SPTs), from A through E, according to their reported frequencies in outbreaks, human illness, and the presence of MGEs such as locus of enterocyte effacement (LEE) and OI-122. In our study, all top 6 strains were classified as SPT-B (associated with outbreaks and Hemolytic uremic syndrom (HUS), but less commonly than serotype O157:H7) with strains from O91, O113, and O128 classified as SPT-C (associated with sporadic HUS, but not typically with outbreaks). Previous studies demonstrated a relationship between the presence of stx2 and the severity of human disease, including the development of HUS and bloody diarrhea with O26, O103, O111, and O145 (Boerlin et al., 1999; Friedrich et al., 2002). However, in the present study, we found that stx2 genes were more prevalent in the SPT-C. A low prevalence of stx2 in top 6 strains in the present study aligned with previous findings where a higher percentage of stx1 than stx2 was observed in strains from serogroups, O26, O103, and O111 (Tayzar et al., 2013; Anjum et al., 2014).
The eae in the LEE is also thought to be a significant determinant of STEC virulence (Etcheverria and Padola, 2013). In this study, eae was detected in all strains from top 6 serogroups and in only one O128:NM strain, an observation that agrees with previous reports (Girardeau et al., 2005; Kobayashi et al., 2013).
Other virulence genes, including toxB, adfO, sodCF, ureA, and efa1, were primarily associated with top 6 strains. Anjum et al. (2014) showed that the top 6 STEC were likely to harbor adfO, sodCF, ureA, and efa1. All of these virulence-associated genes are located on MGEs in STEC, including OI-57 (adfO) (Imamovic et al., 2010), lambdoid prophage (sodCF) (D'Orazio et al., 2008), OI-43 or OI-48 (urea) (Yin et al., 2009), and OI-122 (efa1) (Karmali et al., 2003). Overall, these data suggest that the MGEs substantially contribute to the virulence of STEC strains.
MDR of non-O157 STEC
AMR among foodborne bacteria has been rising since the early 1990s (Walsh and Fanning, 2008). One-third of non-O157 STEC strains evaluated were MDR, with the highest resistance rates observed for STR, SUL, and TET. Other studies from United States (Ju et al., 2012), Spain (Cabal et al., 2013), Belgium (Buvens et al., 2010), and India (Rajkhowa and Sarma, 2014) have made similar observations. Interestingly, STR-SUL-TET resistance was more likely to be detected from animal than human strains. The high frequency of STR-SUL-TET resistance reported among non-O157 strains may, in part, reflect the common use of these antibiotics for growth promotion and disease prevention in food animals (McEwen and Fedorka-Cray, 2002; USDA, 2009)
In the present study, MDR was found in 12 non-O157 strains with 67% possessing a strong or intermediate ability to form biofilms. Previously, Ito et al. (2009) indicated that generic E. coli in mature biofilms were highly resistant to antimicrobial agents, an outcome that could complicate the disinfection of food contact surfaces (Giaouris et al., 2014). STEC strains forming biofilms with resistance to antimicrobials highlight current challenges of antimicrobial-based sanitation measures for biofilm removal. This is especially true as strains exhibiting AMR often exhibit cross-resistance to many common disinfectants (Ryu et al., 2004; Fouladkhah et al., 2013).
Conclusions
All non-O157 STEC strains evaluated were able to form biofilms, with the majority exhibiting high biofilm-forming potential. Biofilm formation varied with serogroup and origin was generally enhanced at room temperature with prolonged incubation times. Of the genes screened for biofilm formation, no specific combination appeared to be associated with enhanced biofilm formation. In addition, eae, toxB, adfO, sodCF, ureA, and efa1 were the dominant virulence profile in top 6 strains. However, a high percentage of strains from serogroups, O91, O113, and O128, carried stx2 , highlighting that serogroups other than top 6 can carry genes with implications for human health. Furthermore, MDR was found in non-O157 STEC strains and most displayed intermediate or strong ability to produce biofilms. Consequently, future strategies to control non-O157 STEC may include measures that effectively control biofilms.
Footnotes
Acknowledgments
Financial support was provided by the Alberta Livestock and Meat Agency, Agriculture and Agri-Food Canada, and China Scholarship Council. The authors gratefully acknowledge R. Barbieri, S. Cook, and R. Zaheer from AAFC and T. Rueter, J. Peters, S. Trapp, Y. Graham, and C. Conrad from AF for technical assistance and support.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.