
Abstract
Foodborne disease outbreaks involving fresh produce have increased in recent years. The risk of infection from contaminated food is worsened by the increased prevalence of antibiotic-resistant strains. This study evaluated the prevalence of antibiotic resistance in Salmonella isolates (n = 263) from agricultural production systems through to the final packed product. Salmonella isolates were preliminarily identified by matrix-assisted laser desorption ionization–time-of-flight mass spectroscopy (MALDI-TOF MS) and API 20E and identities confirmed by invA gene polymerase chain reaction. Antimicrobial susceptibility was performed with 15 antimicrobial agents using the Kirby–Bauer disk diffusion test. Of the 263 Salmonella isolates assessed, 59.3% were resistant to one or more antimicrobials. The most frequently detected resistance was against chloramphenicol and kanamycin (46.7%), trimethoprim–sulfamethoxazole (28%), and streptomycin (14%), and the less frequently detected resistance was toward ampicillin (1.14%), amikacin (0.76%), and amoxicillin–clavulanic acid (0.38%). Multiple antimicrobial resistance (MAR) (resistance to ≥3 antibiotics) was found in 48.7% (76/156) isolates. The most common MAR phenotype was to chloramphenicol and trimethoprim/sulfamethoxazole–kanamycin (43.6%). Resistance to chloramphenicol, kanamycin, or trimethoprim/sulfamethoxazole was only observed in MAR phenotypes. All isolates were susceptible to ceftiofur, cefoxitin, ceftriaxone, ciprofloxacin, nalidixic acid, gentamicin, and tetracycline. This study confirms the importance of fresh produce production environments as potential reservoirs and fresh produce as carriers of antibiotic-resistant Salmonella spp. with significant clinical importance. Further studies to evaluate the actual level of health risk from these pathogens should include characterization of the antibiotic resistance determinant genes among the isolates.
Introduction
I
Studies to evaluate antibiotic resistance in fresh produce and pre- and postharvest environments are limited, and the results are often conflicting. Some studies report that Salmonella isolates of plant origin food are generally susceptible to the commonly used antibiotics (Sivapalasingam et al., 2003; Greene et al., 2008; Patchanee et al., 2010; Gorski et al., 2011). Conversely, Salmonella-resistant strains are gradually being reported in fresh produce production systems (Zhao et al., 2003; Singh et al; 2007; Learn-Han et al., 2009; Li et al., 2014; Abakpa et al., 2015). Any efforts to control antibiotic resistance in the fresh produce chain should therefore systematically evaluate the role of production environments to act as reservoirs or carriers of antibiotic-resistant bacteria (Chidamba and Korsten, 2015). In light of the health implications associated with drug-resistant Salmonella strains, the emergence of antibiotic-resistant Salmonella in the horticultural system is a new and emerging food safety challenge, which deserves further study. This study was undertaken to assess antimicrobial resistance (AR) profiles of Salmonella isolates from commercial horticultural environments.
Materials and Methods
Sampling sites
Three large commercial farms located in three different regions/provinces were selected for this study. The farms were export oriented and GLOBALG.A.P. certified and had their own packhouses. The types of fruit produced on these farms as well as the exact locations are not given due to confidentiality reasons. Moreover, it was not the purpose of this study to investigate a specific fruit type but rather a horticultural production system. The selected sites represent the major fruit production regions with no nearby large-scale cattle farm, feedlot, or battery poultry farming. The water used on these farms is extracted from large nearby river systems. The quality of this water is at times compromised due to nearby informal settlements or municipal wastewater (Fatoki et al., 2001; De Villiers, 2007).
Sample collection
A total of 491 samples comprising fruit (225), agricultural water (140), and packline and hand swabs (126) were collected from the three commercial farms (A, B, and C) and associated packhouses during June to September 2011. Water samples were collected from both pre- and postharvest stages at each farm and at the associated packhouses. Preharvest water samples were collected from river, dam, and/or storage tanks. Irrigation water samples were collected as described in ISO 5667-10 (1992) from pipes along the drip line in the same orchards, where fruit samples were collected.
Fruit samples included low-hanging fruit regularly watered with the microirrigation system in the orchard and fruit (from the same orchard) collected from the harvesting crates before wash and at various points further down the packline, including after washing, fungicide treatment, wax application, and in the final packed box. Swab samples (Medical Wire and Equipment) were collected from hands of packhouse personnel handling the fruit and ∼25-cm2 conveyor belt surfaces that came into contact with the same batch of fruit by thoroughly swabbing with at least 10 passes vertically and horizontally. All samples were collected aseptically and kept chilled at 4°C for no longer than 48 h before analysis.
Salmonella detection, isolation, and identification
All microbiological media were purchased from Merck (Johannesburg, South Africa), unless otherwise stated. All samples (n = 491) were analyzed for Salmonella following the U.S. Food and Drug Administration (U.S. FDA) Bacteriological Analytical Manual (BAM) protocol for fresh produce (Andrews and Hammack, 2007) with minor modifications. Fruit samples (three fruits per sample) were submerged in 1 L sterile 0.1% buffered peptone water supplemented with 0.02% (v/v) Tween 80 (Associated Chemical Enterprises). Microbial epiphytes on fruit surfaces were dislodged by sonication in a digital heated ultrasonic cleaner (Eumax, UD200SH-6L; Labotec) for 5 min at 200 W and 50 Hz. The fruit microflora washings and 1 L water samples were concentrated by filtration through a 0.45-μm pore size nitrocellulose membrane (Sartorius).
Subsequently, membranes and swabs were aseptically transferred into 9 mL tryptone soy broth and incubated with agitation for 24 h at 37°C. An aliquot (0.1 mL) of the preenriched broth was inoculated into 10 mL Rappaport medium for selective enrichment and isolation of Salmonella as previously described (Gorski et al., 2011). A total of 263 Salmonella were isolated and preliminarily identified by matrix-assisted laser desorption ionization–time-of-flight mass spectroscopy (MALDI-TOF MS) (Dieckmann and Malorny, 2011) and API 20E (Holmes et al., 1978) and identities confirmed by invA gene polymerase chain reaction (PCR) as previously described (Rahn et al., 1992).
Antimicrobial susceptibility testing
Isolates were tested for antimicrobial susceptibility using the Kirby–Bauer disk diffusion method on Muller–Hinton agar (Oxoid) as previously described (Gorski et al., 2011) in accordance with the Clinical and Laboratory Standards Institute (CLSI, 2007) guidelines. Fifteen antimicrobial agents (Mast Diagnostics, supplied by Davies Diagnostics) representing eight classes of drugs were tested in this study (Table 1).
Number of antibiotics to which isolates were resistant.
Other antibiotics to which all isolates were susceptible to included tetracycline, nalidixic acid, ciprofloxacin, cefotaxime, cefoxitin, ceftazidime, ceftriaxone, and gentamicin.
AK, amikacin; AP, ampicillin; AUG, amoxicillin–clavulanic acid; K, kanamycin; C, chloramphenicol; S, streptomycin; TS, trimethoprim–sulfamethoxazole.
Genotypic determination and serotyping of Salmonella isolates
All 263 isolates confirmed as Salmonella spp. were typed with the primer sets BOX-A1R and (GTG)-5 as previously described (Versalovic et al., 1994; Rademaker and de Bruijn, 1997). A total of 39 representative isolates were systematically selected to represent various AR and rep-PCR (BOX-A1R and (GTG-5)) profiles, sources, and sample types. Selected isolates were serotyped at the Onderstepoort Veterinary Institute Bacteriology Laboratory, Pretoria, as previously described (Grimont and Weill, 2007).
Statistical analysis
Statistica 10 (StatSoft) was used for all the statistical analyses. Data on antibiotic resistance of each bacterial isolate were reported either as the inhibition diameter (in millimeters) or as susceptible or resistant (based on CLSI, 2011 breakpoints). To avoid overestimation of resistance, all isolates that showed intermediate resistance were reclassified as susceptible (Ta et al., 2014). Multiple antimicrobial resistance (MAR) index defined as a/b, where “a” is the number of antibiotics to which a particular isolate is resistant to and “b” is the total number of antimicrobials tested (Krumperman, 1983), was determined for each of the isolates tested. Electrophoretic gels were analyzed as previously described (Chidamba and Bezuidenhout, 2012).
Results
A total of 26 (5.3%) of the 491 samples analyzed were positive for Salmonella, and 263 isolates were recovered. At site A, Salmonella was detected from three orchard water (33 isolates), two orchard fruit (20 isolates), and nine packhouse conveyor belt swab (54 isolates) samples. At site B, Salmonella was detected from four packhouse fruit samples: after wash (16 isolates), warm bath (25 isolates), waxing and packed fruit (11 isolates each), and two packhouse wash water (21 isolates) samples. At site C, Salmonella was detected from one dam water (23 isolates), four orchard fruit (35 isolates), and one packed fruit (14 isolates) samples. All isolates were preliminarily identified with MALDI-TOF MS as Salmonella spp., with score values ranging from 2.292 to 2.537 and harbored the invA gene. The data on antibiotic resistance profiles to a panel of 15 antibiotics are shown in Table 1. Antibiotic resistance was observed against three aminoglycosides (amikacin [0.76%], kanamycin [47.1%], and streptomycin [13.7%]), two β-lactams (ampicillin [0.76%] and amoxicillin–clavulanic acid [0.38%]), phenicols (chloramphenicol [47%]), and folate pathway inhibitors (trimethoprim–sulfamethoxazole [28.1%]). All the isolates tested were susceptible to tetracyclines (tetracycline), quinolones (nalidixic acid), fluoroquinolones (ciprofloxacin), cephalosporins (cefotaxime, cefoxitin, ceftazidime, and ceftriaxone), and one aminoglycoside (gentamicin).
On average, 59.3% (156/263) of the Salmonella isolates were resistant to at least one or more antimicrobials tested in this study. Resistance toward ampicillin, amikacin, and amoxicillin–clavulanic acid was observed in small proportions (1.14%, 0.76%, and 0.38%, respectively).
A comparison of resistance levels of isolates from different types/sources of samples across sampling sites revealed that isolates from conveyor belt swabs for site A and fruit from the orchard and final packs at site C were the most resistant and exhibited 100% resistance to chloramphenicol and kanamycin. Although resistance to chloramphenicol, trimethoprim–sulfamethoxazole, kanamycin, and ampicillin was observed for isolates from sites A and C, no such resistance was observed for isolates from site B. Resistance to amoxicillin–clavulanic acid was only observed in one isolate from the final packed fruit at site C. Similarly, ampicillin resistance was only detected in conveyor swabs at site A.
All chloramphenicol-resistant isolates also exhibited resistance to kanamycin. Antibiotic resistance phenotypes observed in the current study included single, dual, and multiple antibiotic resistances (up to four different antibiotics) (Table 1). Of the 156 antibiotic-resistant isolates, 48.7% (76/156) exhibited MAR (≥3) distributed among six different patterns (AP-C-K, C-K-AUG, C-TS-K, C-AP-TS-K, C-TS-K-AK, and C-TS-S-K), with MAR indices ranging from 0.2 (3/15) to 0.27 (4/15) (Table 1). The most common MAR phenotypes included combinations of three antimicrobials (44.9%; 70/156) and to a lesser extent four antimicrobials (3.8%; 6/156). Single AR phenotypes were limited to S and AK, whereas the only dual AR phenotype observed was C-K.
Genotyping of the 263 Salmonella isolates with rep-PCR (BOX-A1R and (GTG)-5 primers) resulted in a diversity of profiles (Fig. 1). These together with the observed AR for isolates from different sources and sample types resulted in the systematic selection of 39 representative isolates (Fig. 2). Serotyping showed these isolates to belong to the serovars Salmonella Muenchen (13/39; 33.3%), Salmonella Typhimurium (12/39; 30.8%), Salmonella Heidelberg (8/39; 20.5%), Salmonella Bsilla (3/39; 7.7%), Salm IIb: 17: r: z (2/39; 5.1%), and one untypable rough biotype. Cluster analysis showed the isolates to group relative to their serovars, although other clusters were composed of different serovars. The genotype grouping showed the detection of similar strains among different packhouse samples, even though no similar strains could be detected between packhouse and field samples (Fig. 2). Although similar serovars were detected between orchard water and packhouse samples, consideration of their antibiotic resistance showed them to be different strains.
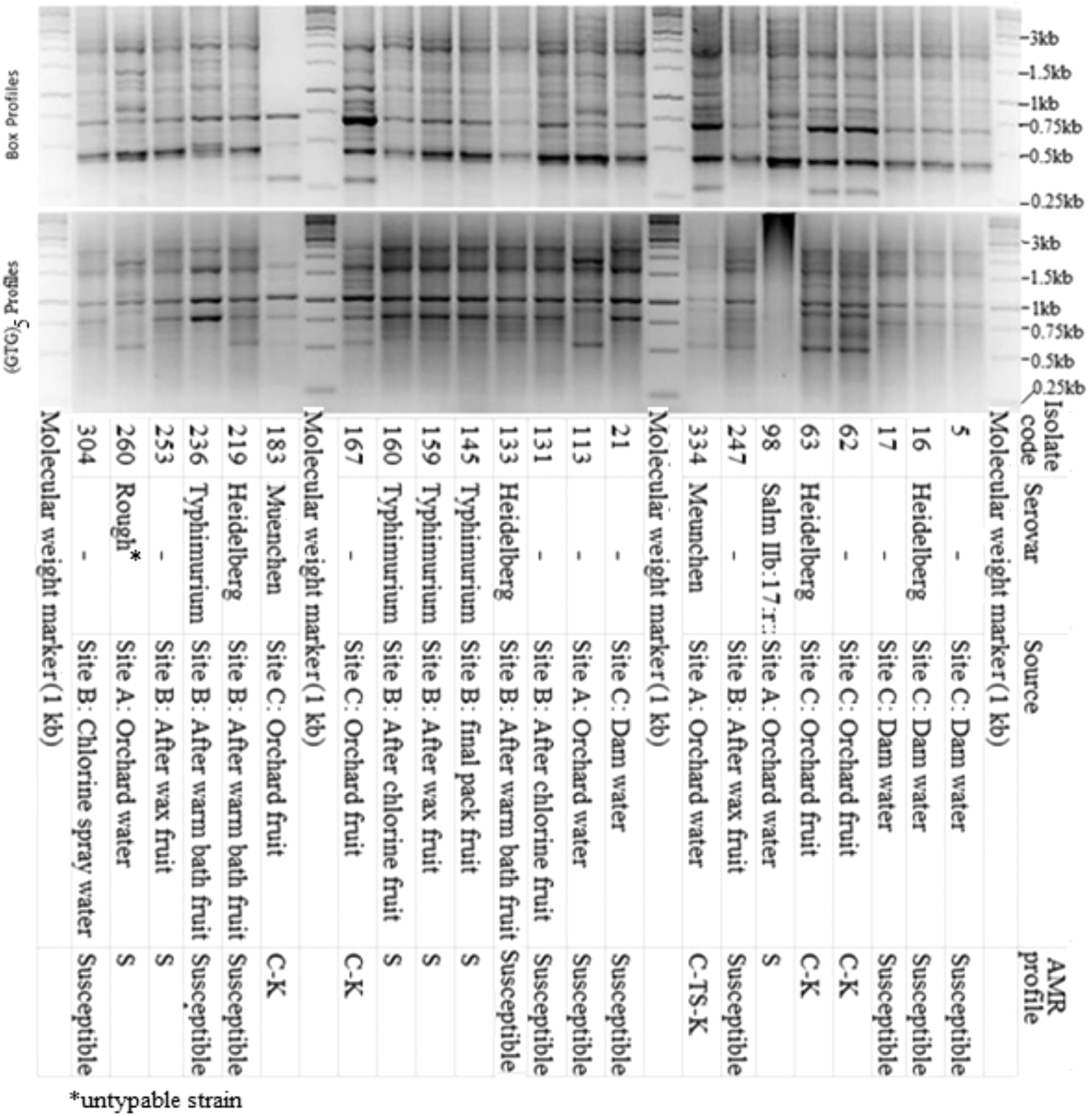
Representative agarose gel images for Salmonella genotyping with BOX and (GTG)5 primers showing the selected representative isolates with respect to source, antibiotic resistance, and serotypes.
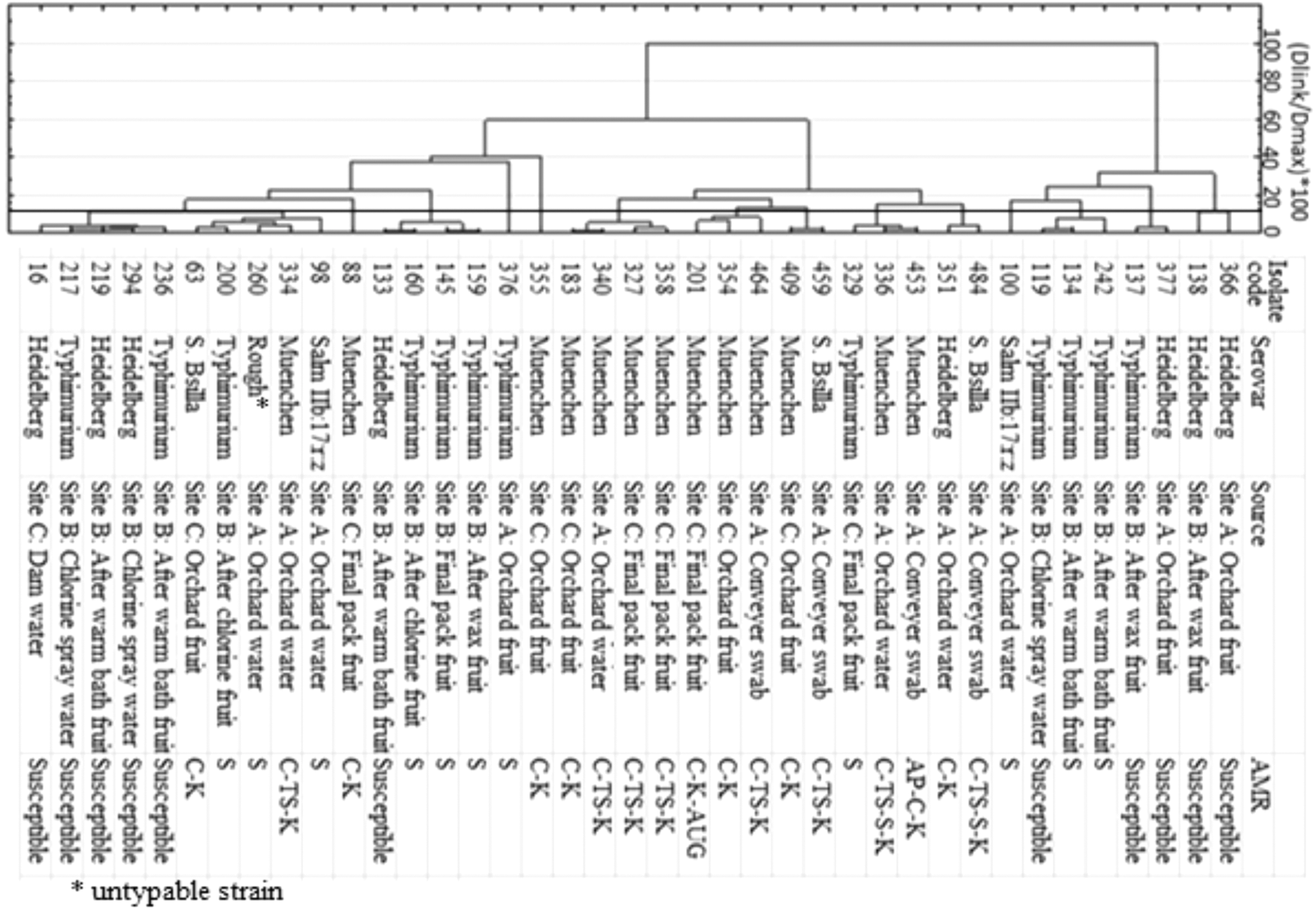
Ward and Euclidean distance clustering of selected Salmonella representative isolates based on BOX and (GTG)5 rep-PCR profiles and their associated serotypes, sources, and antibiotic resistance profiles. PCR, polymerase chain reaction.
Considering the observed antibiotic resistance profiles, the serotype Salmonella Muenchen had the highest antibiotic resistance observed with five AR profile types (Fig. 2). Among these, the AR profiles C-K and C-TS-K were the most prevalent. The serovar Salmonella Bsilla had three AMR profile types. The rough type and Salm IIb: 17: r: z had a single AR profile type, while Salmonella Typhimurium and Salmonella Heidelberg having two phenotypes each. The highest MAR was observed for the C-TS-S-K combination in Salmonella Bsilla and Salmonella Muenchen. The serovars Salmonella Heidelberg and Salmonella Typhimurium were the only serovars with isolates susceptible to all antibiotics tested.
Discussion
AR among Salmonella spp. isolates from fresh produce is a serious public health concern given that these products are mostly consumed raw or just minimally processed. Although the finding of AR in Salmonella (59.3%) in this study is high, it is relatively lower than previous findings, which reported high levels (82.9%) of AR in Salmonella isolated from market vegetables in northern India (Singh et al., 2007). However, the Salmonella isolates were collected at the market end and did not include any isolates from the primary production environments. In contrast, the majority of the Salmonella isolates recovered from tomato farm environments in the Mid-Atlantic region of the United States (Micallef et al., 2012) and crop production watersheds in North Carolina (Patchanee et al., 2010) were susceptible to most of the antibiotics tested. Differences in antimicrobial use patterns and associated selection pressures could possibly explain the observed variation in Salmonella resistance profiles reported from different countries or regions.
Frequent resistance observed for chloramphenicol, kanamycin, and trimethoprim/sulfamethoxazole in this study could be due to their use in human and veterinary medicine for many years (Li et al., 2014). Our findings were inconsistent with results from Duffy et al. (2005), Learn-Han et al. (2009), Gorski et al. (2011), and Micallef et al. (2012), who reported low or no resistance to these drugs. In contrast, Osterblad et al. (1999) noted a chloramphenicol resistance rate of 12% among Enterobacteriaceae isolated from vegetables. Although the use of chloramphenicol as a growth promoter in animal production has been prohibited in developed countries owing to induce anemia in humans, this drug has for long been on the list of first-line drugs to treat numerous human infections in sub-Saharan Africa (Okeke and Sosa, 2003). Furthermore, the presence of trimethoprim/sulfamethoxazole-resistant Salmonella isolates is a cause of concern since this drug is widely used to treat systemic Salmonella infections and typhoid fever in humans (WHO/UNAIDS, 2000).
Similar to our findings, Micallef et al. (2012) also reported very low resistance percentages for amoxicillin/clavulanic acid and ampicillin. According to the Veterinary Drug Directorate, Health Canada, amoxicillin/clavulanic acid is classified in category 1 (very high importance in human medicine) (Mainali et al., 2014). All Salmonella isolates were susceptible to cefoxitin, ceftriaxone, ceftazidime, cefotaxime, nalidixic acid, ciprofloxacin, gentamicin, and tetracycline. These findings are similar to those reported by Metcallf et al. (2012), who found no resistance toward ciprofloxacin, ceftriaxone, gentamicin, and nalidixic acid but observed resistance toward cefoxitin and tetracycline in Salmonella isolates from the Mid-Atlantic tomato farms. It is important to note that most of their Salmonella isolates were recovered from irrigation ponds at large-scale farms located close to poultry production facilities.
Although tetracycline and ampicillin are widely used both in veterinary and human medicine, none of our isolates presented resistance to tetracycline, while only one isolate was resistant to ampicillin. A lack of resistance to tetracycline and ampicillin in the present study may imply that the isolates were not of animal origin since these drugs are widely used in livestock farming as feed additives in many countries, and no animal farm was situated near the sampling sites in this study (Geonaras et al., 2001; Henton et al., 2011; Mainali et al., 2014). With the exception of streptomycin and kanamycin, resistance toward other members of the aminoglycosides tested in this study (gentamicin and amikacin) was not observed or insignificant. Use of antibiotics in this class, apart from streptomycin that is mostly used to treat tuberculosis patients, has been reduced in many countries (Goni-Urriza et al., 2000), and this may account for the low resistance levels observed in this study.
Resistance to chloramphenicol, kanamycin, and trimethoprim/sulfamethoxazole was only observed in multiple resistant isolates and never as single antibiotics, mostly at sites A and C. High resistances observed for isolates from sites A and C may be because the isolates were from preharvest stages (storage dam water, orchard irrigation water, and orchard fruit), which are exposed to the outside environment compared to site B isolates, which all came from the packhouse environment with limited pathogen dispersal. Associations among antibiotic resistances in particular isolates can be ascribed to the presence of linked genes in mobile genetic elements, such as plasmids, transposons, or integrons, that harbor one or more resistant genes, each encoding a single antibiotic resistance phenotype (Kelly et al., 2009; Mather et al., 2013).
Due to the nature of horticultural production systems assessed in this study, large-scale commercial farms are mostly in rural areas away from poultry plants or feedlots. In certain cases, informal settlements can be found around upstream areas of the river catchment, which serve as a source of irrigation water. It has been well documented that municipalities in these areas are the major contributor to agricultural water pollution (CSIR, 2010). Consequently, the most likely source of AR in Salmonella in this scenario is municipal polluted rivers used by the farmers in this study. Future studies should focus on source tracking to show the human, animal, sewage, river, crop, and food link.
Although antibiotic inhibition patterns between isolates from water, fruit, and swab samples suggest irrigation water to be the source of Salmonella contamination in packhouses, consideration of rep-PCR genotypic profiles suggests otherwise. There is evidence of spread of the same Salmonella strains within the packhouses, but no link could be made to the Salmonella observed in the field or irrigation water samples. However, the absence of some genotypes and AR phenotypes in some samples at the same farm and packhouse may be a result of failing to pick the representative isolates during the random selection. Hence, the results of this study point to contamination of agricultural production environments by a wide diversity of AR Salmonella spp. most likely contributed by polluted river water used for irrigation or packers in the packhouses.
Conclusions
This study confirms the importance of fresh produce production environments as potential reservoirs and fresh produce as carriers of antibiotic-resistant Salmonella spp. However, there was no link between production site and packhouse/fruit contamination in the study. The AR toward the commonly used antimicrobials demonstrated in our study is a cause for concern. However, it is encouraging that all isolates were susceptible to the more clinically important drugs (third-generation cephalosporins, quinolones, and fluoroquinolones) used for treating life-threatening infections caused by Salmonella. Nevertheless, ongoing monitoring of AR is necessary to evaluate overall resistance pools and trends and explore risks and risk mitigation strategies in horticultural supply chains.
Footnotes
Acknowledgments
The authors are thankful for funding from the National Research Foundation (NRF) for providing funds to purchase the MALDI-TOF MS used in the preliminary identification of the isolates. Special recognition goes to all farmers and packhouse managers for giving us access to their orchards and packhouses.
Disclosure Statement
No competing financial interests exist.