
Abstract
Increased enterococcal infections in hospitals and multidrug-resistant and vancomycin-resistant enterococci (VRE) isolated from humans, animals, and food sources raised public health concern on the presence of VRE in multiple sources. We performed a comparative analysis of the antimicrobial resistance and genetics of VRE isolates derived from fresh produce and human fecal samples. Of 389 Enterococcus isolates, 8 fecal and 3 produce isolates were resistant to vancomycin and teicoplanin; all harbored vanA gene. The VRE isolates showed multidrug-resistant properties. The isolates from fresh produce in this study showed to have the common shared characteristics with the isolates from humans by the results of antimicrobial resistance, multilocus sequence typing, and Tn 1546 transposon analysis. Therefore, VRE isolates from fresh produce are likely related to VRE derived from humans. The results suggested that VRE may contaminate vegetables through the environment, and the contaminated vegetables could then act as a vehicle for human infections. Ongoing nationwide surveillance of antibiotic resistance and the promotion of the proper use of antibiotics are necessary.
Introduction
S
The vanA gene cluster is related to the mobile genetic element Tn1546, which is a transposon that is commonly carried by a self-transferable plasmid. Plasmid transfer could be responsible for the spread of glycopeptide resistance among pathogens, especially Gram-positive cocci (Aarestrup et al., 1996; Nilsson et al., 2009).
Typically, VRE are reported in patients (Banerjee and Anupurba, 2015), but they have recently been confirmed in various other sources, such as broilers (Nilsson et al., 2009), animal-derived foods (Cetinkaya et al., 2013), river water, dental plaques, and food products (Creti et al., 2004). Although Korea banned the use of avoparcin and other antibiotics as feed additives for food animals, VRE have been continuously detected in both food animals and humans. However, little is known about VRE derived from fresh produce. Therefore, we characterized and compared VRE isolates derived from fresh produce and human feces based on analyses of antimicrobial resistance, multilocus sequence typing, and Tn1546 transposon properties.
Materials and Methods
Bacterial isolates
Enterococcus isolates were derived from 830 samples, 540 fresh produces and their environments and 290 human fecal samples. Fresh produce samples were collected from a surveillance project from fresh produces and their environments in 2011, and fecal samples of diarrheal patients were provided by Gyeonggi Institute of Health and Environment in 2012 in Korea. Enterococcus faecium (EFM) and Enterococcus faecalis (EFS) were isolated by using Azide dextrose broth with added 6.5% NaCl for enumeration and Enterococcosel agar for selection of colonies. VITEK 2 (bioMérieux, Marcy I'Etoile, France) was used for the identification of Enterococcus spp. The isolates from fecal samples were cultured on Enterococcosel agar without any enrichment step and identified.
Detection of vancomycin-resistant Enterococcus
To detect vancomycin-resistant isolates, antimicrobial susceptibility tests were performed on vancomycin (30 μg) and teicoplanin (30 μg) using the disk diffusion method. The minimum inhibitory concentrations (MICs) of vancomycin (>32 μg/mL) and teicoplanin (>32 μg/mL) were determined using the agar dilution method according to the Clinical Laboratory Standards Institute (CLSI) guidelines (2014). E. faecalis ATCC 29212 was used as the quality control strain. VanA gene was detected by polymerase chain reaction (PCR) (Miele et al., 1995).
Antimicrobial susceptibility testing
Antimicrobial susceptibility tests were performed for vancomycin-resistant isolates using the disk diffusion method according to CLSI guidelines (2014). The antimicrobial agents and their corresponding concentrations were as follows: ampicillin (10 μg), chloramphenicol (30 μg), erythromycin (15 μg), gentamicin (120 μg), linezolid (30 μg), penicillin (10 μg), rifampin (5 μg), quinupristin/dalfopristin (15 μg), streptomycin (300 μg), and tetracycline (30 μg). All antimicrobial discs were BBL products (Becton-Dickinson Microbiology Systems, Cockeysville, MD). S. aureus ATCC 25923 was used as a quality control strain.
Multilocus sequence typing
For the multilocus sequence typing (MLST) analysis, PCR was performed using seven housekeeping genes in E. faecium (gdh, purK, pstS, atpA, gyd, adk, and ddl) (Homan et al., 2002) and E. faecalis (gdh, gyd, pstS, gki, aroE, xpt, and yqiL) scheme (Ruiz-Garbajosa et al., 2006), respectively. The sequences were generated by the Macrogen Service Center (Macrogen, Seoul, South Korea). The DNA sequences were searched against MLST databases (
Pulsed-field gel electrophoresis
On the basis of the PulseNet standard protocol, modified pulsed-field gel electrophoresis (PFGE) was applied using the CHEF Mapper system (Bio-Rad Laboratories, CA) (Choi and Woo, 2013). SmaI (Takara Bio, Shiga, Japan) was used for the digestion of DNA. PFGE conditions were as follows; gradient 6.0 V/cm at 14°C for 20 h with pulse times ramped from an initial start time of 1 s to a final start time of 20 sin 0.5 × Tris/borate/EDTA buffer. The genetic relatedness was analyzed by InfoQuest FP software, version 4.5 (Bio-Rad Laboratories, CA).
Tn1546 transposon typing
To analyze the composition of transposon Tn1546 harboring the vanA gene cluster, the target genes were amplified by PCR (Table 1). Pairs of highly amplified fragments were sequenced by the Macrogen Service Center (Macrogen, Seoul, Korea). The sequences were compared to the reference strain EFM BM 4147. The structure of the resistance genes was analyzed based on the results of PCR amplifications of IS1216V and IS1542 (Lee et al., 2003).
Statistical analyses
The proportions of Enterococcus isolates, E. faecium and E. faecalis, were computed from 540 fresh produces and their environments and 290 fecal samples, respectively. The proportions of vancomycin-resistant Enterococcus were derived from 160 fresh produces and their environments and 229 fecal isolates, respectively. Chi-square tests of homogeneity were conducted to test the difference between two proportions in sample sources (fresh produces vs. fecal samples) and Enterococcus types (E. faecium vs. E. faecalis). All statistical analyses were performed using SAS software version 9.3 (SAS Institute, Cary, NC) with a significance level of α = 0.05.
Results
Isolation of Enterococcus
A total of 389 Enterococcus isolates were derived from 830 samples. From the 540 fresh produce and their environment samples, 83 (15.4%) E. faecium and 77 (14.3%) E. faecalis were isolated, and 102 (35.2%) E. faecium and 127 (43.8%) E. faecalis were isolated from the 290 fecal samples. Chi-square tests suggested that the proportions of E. faecium and E. faecalis isolates in fecal samples (35.2% and 43.8%) were significantly higher than the proportions in fresh produce samples (15.4% and 14.3%), respectively (p < 0.0001), as shown in Table 2.
VRE, vancomycin-resistant enterococci.
Isolation of VRE
From a total of 389 Enterococcus isolates, 11 strains were resistant to both vancomycin (>128 mg/L) and teicoplanin (>32 mg/L) in the MIC test, including 3 isolates (1.9%) from fresh produce and 8 isolates (3.5%) from human fecal samples. These 11 isolates harbored the vanA gene. Therefore, vancomycin-resistant E. faecalis and E. faecium isolated from produce and fecal samples were identified as VanA phenotype-vanA genotype. For fresh produces, 2 E. faecium (2.4%) and 1 E. faecalis (1.3%) were VREs from 83 E. faecium and 77 E. faecalis, respectively. For fecal samples, 7 E. faecium (6.9%) and 1 E. faecalis (0.8%) were VREs from 102 E. faecium and 127 E. faecalis, respectively. In fecal samples, the proportion of vancomycin-resistant E. faecium isolates (6.9%) was significantly higher than the proportion of vancomycin-resistant E. faecalis (0.8%) (p = 0.013). However, the proportions of vancomycin-resistant E. faecium and E. faecalis isolates (2.4% and 1.3%) were not significantly different in fresh produce samples (p = 0.605).
Antimicrobial resistance profile
Of the 11 VRE isolates, 100% were resistant to ciprofloxacin, 91% were resistant to penicillin, and 73% were resistant to erythromycin, rifampin, and gentamicin. Multidrug resistance profiles, including resistance to more than 7 tested antibiotics, were identified for produce and fecal isolates (Table 3). There was no correlation between the antimicrobial-resistant patterns and the sample sources.
P, fresh produce; F, fecal sample.
TEC, teicoplanin; VA, vancomycin; AM, ampicillin; P, penicillin; C, chloramphenicol; E, erythromycin; GM, gentamicin; LZD, linezolid; TE, tetracycline; RA, rifampin; S, streptomycin; SYN, quinupristin/dalfopristin.
MICs, minimum inhibitory concentrations.
Multilocus sequence typing
Four STs, ST 78 (4 isolates), ST 192 (2 isolates), ST 230 (1 isolate), and ST 262 (2 isolates), were observed in the E. faecium isolates. These STs grouped into three clonal complexes (CCs), that is, CC18 (ST 262), CC78 (ST 78, ST 230), and CC192 (ST 192), which were derived from CC17. Therefore, all vancomycin-resistant E. faecium isolates obtained from fresh produce and fecal samples were included in the CC17 group, which is a well-known hospital-adapted strain. Vancomycin-resistant E. faecalis confirmed as ST 64 and ST 152 was classified as CC8 and CC4, respectively (Table 3).
Pulsed-field gel electrophoresis
As shown in Figure 1, 11 VRE were grouped into three groups based on PFGE profile, Group A, Group B, and Group C. Fresh produce isolates were classified into Group A, and the isolates from fecal samples were grouped into Group B or C. The isolates from Group B such as 12RDA-EFM-3 and 12RDA-EFM-4, 12RDA-EFM-17 and 12RDA-EFM-18 showing more than 90% genetic similarity were originated from a nearby sampling place and at a similar sampling time from the diarrheal patients.
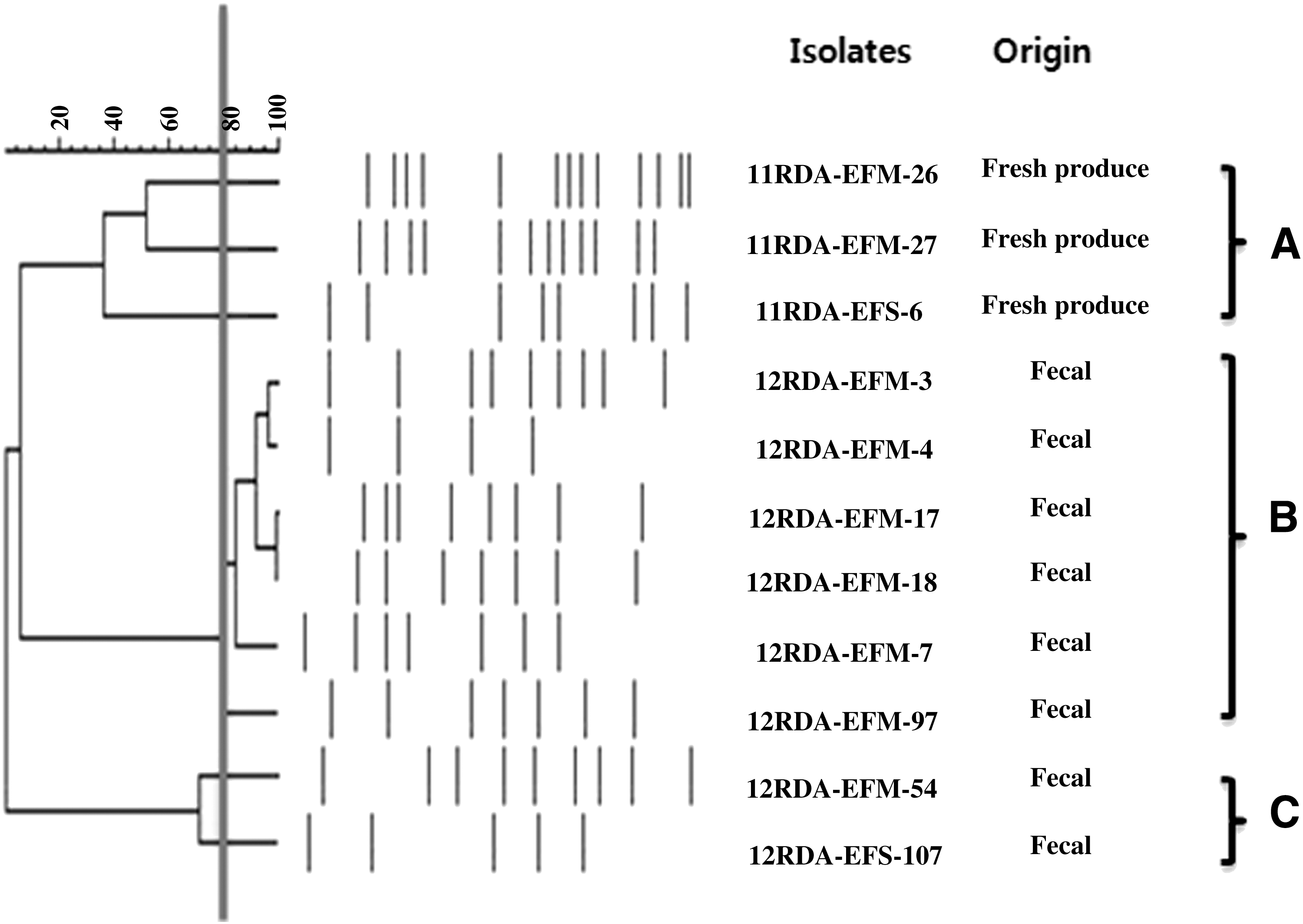
PFGE profile of VRE isolates. Fresh produce isolates were classified into Group
Tn 1546 transposon typing
All VRE isolates were divided into six groups on the basis of insertion sequences (ISs) and the deletion and/or insertion of genes in the vanA cluster. None of the isolates was identical to the Tn1546 of E. faecium strain BM 4147 (Fig. 2). Two vancomycin-resistant E. faecium produce-derived isolates (11RDA-EFM-26 and 11RDA-EFM-27) and two vancomycin-resistant E. faecium fecal isolates (12RDA-EFM-17 and 12RDA-EFM-18) had identical transposons. There was no deletion in nine clusters, and two insertion regions were confirmed, that is, the orf2-vanR intergenic region and vanX and vanY intergenic region, with respect to the prototype. An IS1542insertion was detected in the orf2-vanR region and an IS1216V insertion was detected in the vanXY region. Three fecal isolates (12RDA-EFM-3, 12RDA-EFM-7, and 12RDA-EFM-4) had a deletion in the orf1 region as well as IS1542 and IS1216V insertions. One fecal isolate (12RDA-EFM-54) had a deletion in the orf1 region and the vanX-vanY intergenic region. Two fecal isolates (12RDA-EFM-97 and 12RDA-EFS-107) showed deletions in orf1 and orf2 and an IS1216V insertion in vanXY. One vegetable-derived isolate (12RDA-EFS-6) had a unique IS1216V insertion. A difference between fecal isolates and produce-derived isolates was not confirmed with respect to the Tn1546 cluster.
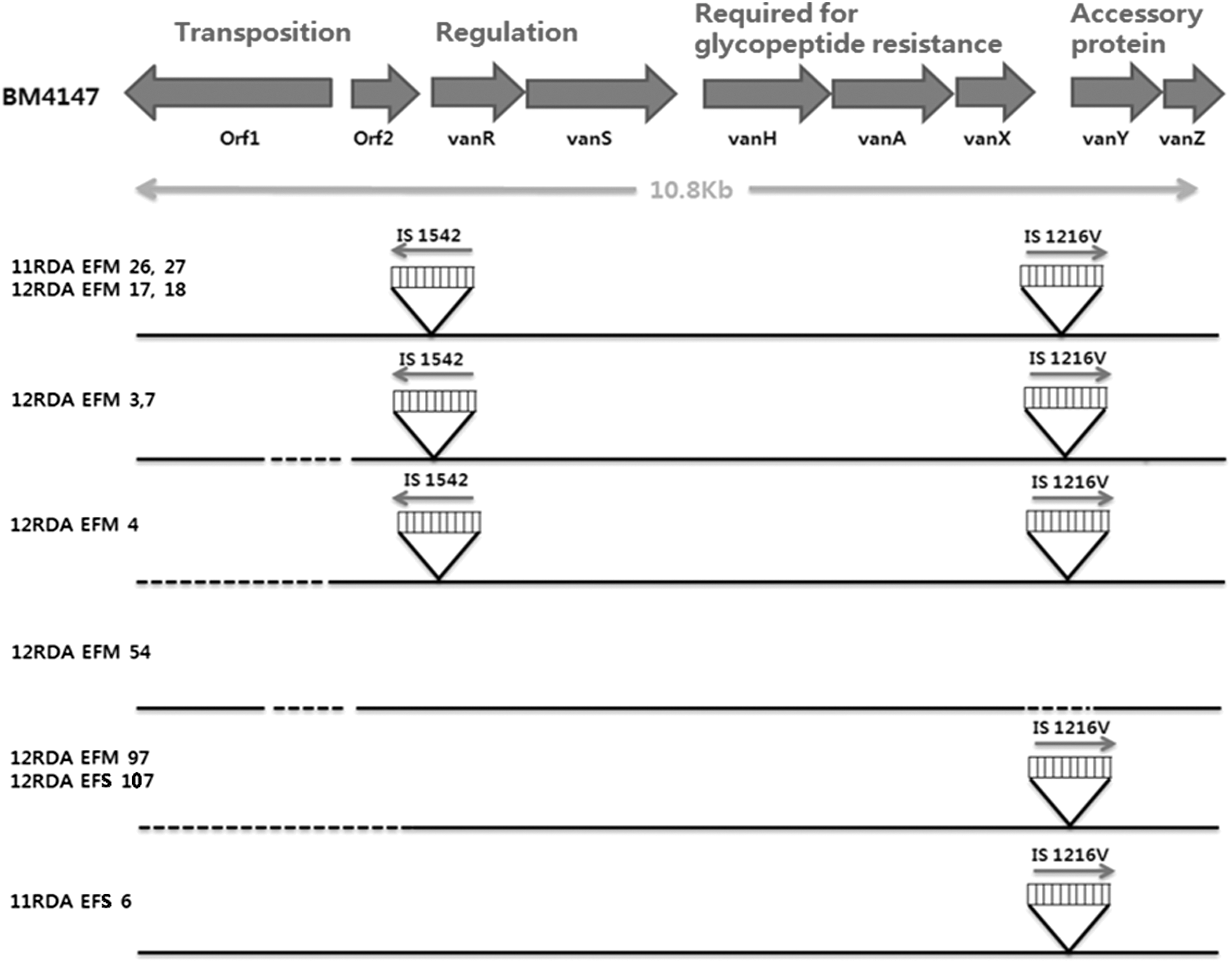
Structures of the Tn1546 elements in nine Enterococcus faecium and two Enterococcus faecalis isolates. The location of the genes and deletion of a normal Tn1546 element in the strain BM4147 (prototype) are shown at the top. Red arrows represent IS elements. Dotted lines represent deletions. Three isolates (11RDA EFM 26, 27, 6) were fresh produce isolates. IS, insertion sequence.
Discussion
Avoparcin has been used to promote growth in broilers and pigs since 1975. VRE have emerged owing to the similar cross-resistance properties of avoparcin, vancomycin, and teicoplanin (Nilsson, 2012; Cetinkaya et al., 2013). There is an established relationship between antibiotic use and antibiotic resistance. Therefore, most countries have prohibited the use of avoparcin to reduce the risk of VRE development. After banning antibiotics, vancomycin resistance has been declined. For example, the VRE isolation rate from poultry was decreased from 14.6% to 8.0% after banning avoparcin for one and a half years in Italy (Pantosti et al., 1999). According to Danish surveillance report, the isolation rate of glycopeptide-resistant E. faecium from broiler cloacal swabs was also decreased from 72.7% in 1995 to 5.8% in 2000 (Aarestrup et al., 2001).
Although avoparcin use is prohibited in Korea, vanA-type VRE have been detected from the meat and feces of pigs and from raw milk samples (Jung et al., 2007), as well as chicken meat (Shon et al., 2013). The continuous detection of VRE might be explained by coselection. In Denmark, tylosin use in pigs may coselect vancomycin resistance among enterococci since the genes encoding resistance to the two agents are located on the same plasmid. Coselection via copper resistance and a plasmid addiction system have also been suggested (Nilsson, 2012). Jung et al. (2007) proposed that the continuous detection of vanA-type enterococci since the avoparcin ban might be attributable to the frequent use of tetracycline and macrolides as growth promoters and for animal treatment in Korea.
In an MLST analysis, ST 78 and ST 192 were dominant among vancomycin-resistant E. faecium isolates in a Korean hospital (Ko et al., 2005). Consistent with previous reports, four STs belonging to CC17, ST 78, ST 192, ST 262, and ST 230, were detected in vancomycin-resistant E. faecium in this study. ST 17, ST 78, and ST 80 have previously been isolated in Europe and the United States, and most of the vancomycin-resistant E. faecium clinical isolates were classified as CC17, which is associated with a globally epidemic E. faecium clone (Rathnayake et al., 2011). Cha et al. (2012) used MLST in VR-EFM human- and poultry-derived isolates and found that all of the human isolates belonged to CC17, while most of the poultry isolates belonged to CC26. In EFS, CC2 and CC9 are high-risk CCs owing to their vancomycin and gentamycin resistance, β-lactamase production, and pathogenicity islands (Rathnayake et al., 2011). In this study, vancomycin-resistant E. faecalis isolates were ST 64 (CC8) and ST 152 (CC4). CC8 originated in humans in Japan, Cuba, and other locations, and CC4 is associated with human infections in other Asia-Pacific countries (Quinones et al., 2009; Sun et al., 2012; Kudo et al., 2014). Therefore, the relationship of VRE from fresh produce with human VRE isolates should be carefully considered and investigated.
To date, the ISs IS1216V, IS1251, IS1542, IS1476, and IS19 have been detected in VRE antimicrobial resistance genes. IS1216V is found worldwide, IS1542 is found in Europe, and IS1251 is commonly reported in the United States, although there is regional variation. Similar to European isolates, most VRE isolated in Korea have IS1542 and IS1216V, regardless of region and origin (Kim et al., 2010). Recently, IS256 was confirmed from the tetracycline-resistant enterococcal strains from Nigeria (Ayeni et al., 2016).
The genetic similarity of fresh produce and fecal VREs in the PFGE was low, but the antimicrobial-resistant characteristics such as Tn1546 were identical from both VRE groups as shown in Figure 2. Therefore, the dissemination of antimicrobial-resistant determinants through plasmids or transposons can be important factors (Cheng et al. 2015). Shon et al. (2013) studied the duration of colonization for the prolonged carriage of VRE after patient discharge from the hospital, and observed a median duration of VRE colonization of 5.57 weeks. Therefore, patients with specific risk factors should be managed carefully to prevent the transmission of VRE in clinics.
In this study, VRE were detected at low rates in fresh produce and fecal samples, but differences in antimicrobial resistance, virulence factors, and Tn1546 transposons were not observed between sample sources. The results in this study indicated that vegetables may become contaminated with VRE through environmental sources, such as human and animal fecal matter, and, in turn, these contaminated vegetables could act as a vehicle to infect humans and animals. The transmission cycle must be interrupted by various means, such as proper personal hygiene, good agricultural practices compliance, thorough food hygiene practices, and infection control. Therefore, ongoing investigations of antibiotic resistance in hospitals, livestock and agricultural products, as well as environmental samples linked to the use of antibiotics are necessary, and the proper use of antibiotics should be promoted to control VRE proliferation.
Conclusions
In this study, the antimicrobial resistance and genetic properties of VRE isolates derived from fresh produce and human fecal samples were investigated and compared. VRE were detected at a low rate from fresh produce (3 isolates, 1.9%) and fecal samples (8 isolates, 3.5%) out of 389 total Enterococcus isolates. Differences in genetic properties and antimicrobial characteristics between the isolates obtained from fresh produce and fecal samples were not observed. Although VRE were detected at a low level from fresh produce and fecal samples, the potential of hazard might be high in the future if not properly controlled. A continuous nationwide and regional comparative surveillance study on VRE is necessary at the government level and through international cooperation.
Footnotes
Acknowledgments
This research was supported by grants from the Rural Development Administration (PJ00758504, PJ010500). We thank Professor Hyejung Chang (Kyung Hee University) for statistical advice. We also thank the Korea University Food Safety Hall and the Institute of Food and Biomedicine Safety for allowing the use of their equipment and facilities.
Disclosure Statement
No competing financial interests exist.