
Abstract
Seventeen Staphylococcus aureus strains were tested for production of staphylococcal enterotoxin D (SED) and staphylococcal enterotoxin R (SER) in milk and meat juice. SED was secreted in milk by 12 S. aureus strains at 6–54 ng/mL at 24 h and 9–98 ng/mL at 48 h. Another five strains secreted SED at 0.9–1.9 μg/mL at 24 h and at 1.2–2.4 μg/mL at 48 h. Strains producing high levels of SED in milk secreted 77–666 μg/mL of SED in meat juice at 24 h and 132–1225 μg/mL at 48 h. Strains producing lower amounts of SED in milk secreted 228–1109 ng/mL of SED at 24 h and 377–1782 ng/mL at 48 h in meat juice. Tested S. aureus strains produced SER in milk at 33–183 ng/mL at 24 h and 41–832 ng/mL at 48 h. Fourteen strains produced more SER in meat juice than in milk (17- to 232-fold and 15- to 269-fold more at 24 and 48 h, respectively). Three S. aureus strains secreted less than 74 ng/mL of SER in meat juice. Expression pattern of known enterotoxin regulators, that is, agrA, sarA, hld, rot, and sigB, was similar in selected strong and weak SED producers grown in both food matrices and could not explain differences in enterotoxin protein level. This suggests that enterotoxin regulation is more complex than previously thought. We demonstrated that in a number of tested S. aureus strains, production of SED and SER was significantly decreased in milk when compared with meat juice, supporting previous reports. However, certain strains secreted high amounts of SED and SER, irrespective of environment, likely contributing to higher food safety risk.
Introduction
E
The sed gene is encoded along with selj and ser genes on a pIB485-like plasmid (Zhang et al., 1998). sed promoter activity has been shown to be upregulated by Agr and SarA and downregulated by Rot and σB (Cheung et al., 2004; Tseng et al., 2004). Transcription of sed is moderately increased in the postexponential phase of growth, which was shown to result from an Agr-dependent decrease in Rot activity (Tseng et al., 2004; Derzelle et al., 2009). However, Yarwood and Schlievert (2003) suggested that SED is only partially upregulated by RNAIII and that it can be produced at high concentrations independently of Agr.
The gene encoding SER can be carried by a pIB485-like plasmid, together with sed and selj genes or by a family of pF5-like plasmids, which also contain ses and set genes, but not the sed gene (Omoe et al., 2003; Ono et al., 2008). The induction of ser expression was observed in the postexponential growth phase and was sustained throughout the stationary phase, suggesting that ser might be controlled by Agr (Derzelle et al., 2009).
Since most data on SED and SER expression come from studies conducted in microbial broths, our aim was to determine the production of SED and SER by S. aureus strains in milk and meat juice and evaluate expression of known enterotoxin regulators, that is, agrA, sarA, hld, rot, and sigB, during growth of S. aureus in both tested environments.
Materials and Methods
Bacterial strains and growth conditions
Seventeen sed- and ser-positive S. aureus strains were investigated (3 food, 13 human-derived isolates, and 1 reference strain) (Table 1). Enterotoxigenic reference S. aureus strains were kindly provided by Prof. Gerard Lina of the Centre National de Référence des Toxémies Staphylococciques, Faculté de Médecine, Lyon, France.
Ultra-high temperature 0.0% fat cow's milk (Mlekpol, Grajewo, Poland) and minced beef meat were purchased from a local store. Meat juice was obtained as described by Rantsiou et al. (2012) with an additional step introduced after material homogenization. It included supernatant sterilization at 121°C for 20 min and supernatant filtration through a 0.2-μm filter. S. aureus strains were grown in brain-heart infusion (BHI) broth (Biocorp, Warsaw, Poland) supplemented with 1% yeast extract (Biocorp) at 37°C, 230 rpm, overnight. Then, 100 mL of cow's milk or 100 mL of meat juice was inoculated with S. aureus BHI culture to obtain OD600 of 0.02 (∼107 colony-forming unit [CFU]/mL). Optical densities of BHI cultures were measured with Cary 100 UV-Vis Spectrophotometer (Varian, Inc.). Before inoculation, the BHI cultures were washed twice with phosphate-buffered saline to remove residual media. Cultures were incubated up to 48 h at 37°C with constant agitation at 230 rpm.
Samples for ribonucleic acid (RNA) extraction and ELISA were collected at specified time points during each experiment. Cells were quantified by plating serial dilutions of bacteria onto BHI agar at 0, 3, 5, 8, 24, 32, and 48 h in 56, 237, 238, 214, and FRI1151m S. aureus strains (to determine growth curves in milk and meat juice) and at 24 and 48 h in remaining strains. pH was measured in each strain at 24 and 48 h of growth by FE20-FiveEasy™ pH meter (Mettler-Toledo, Greifensee, Swiss). All experiments were carried out in triplicate with two biological replicates.
Detection of enterotoxin genes
Detection of sea-see was performed according to Sharma et al. (2000). The selu gene was detected using the method described by Letertre et al. (2003). For the detection of the tst gene, the primers and conditions were described according to Monday and Bohach (1999). Detection of sei, sem, sen, seo, seh, selj, sek, sel, and sep was done as previously described (Bania et al., 2006; Lis et al., 2009). S. aureus reference strains served as polymerase chain reaction (PCR) controls.
Sandwich ELISA
Recombinant enterotoxins D (rSED) and R (rSER) were obtained by cloning sed and ser genes into pET-22b vector, as described previously by Lis et al. (2012). Regions encoding mature SED and SER were PCR amplified from S. aureus 214 strain. Cloning forward and reverse primers (listed in Table 2) carried the restriction sites for NcoI and XhoI, respectively. Enterotoxins were expressed in Escherichia coli Rosetta cells (Merck, Darmstadt, Germany) using IPTG induction (Sigma-Aldrich, St. Louis, MI) and purified on His-Select Cobalt Affinity Gel (Sigma-Aldrich) with on-column refolding. Rabbit polyclonal anti-SED antibody was purchased from Acris Antibodies (Herford, Germany). Rabbit polyclonal anti-SER antibody was obtained as described by Lis et al. (2012). Antibodies conjugated with biotin N-hydroxysuccinimide ester (Sigma-Aldrich) were used as secondary antibodies.
RT-qPCR, reverse transcription-quantitative polymerase chain reaction.
Samples for SED and SER detection were collected at 24 and 48 h of growth and stored at −20°C until analyzed. ELISA was performed as described earlier (Schubert et al., 2016). The specificity of the ELISA was assessed using culture supernatants of S. aureus reference strains as controls for enterotoxins SEA, SEB, SEC, SED, SEE, SEG, SEH, SEI, SElJ, SEK, SEL, SEM, SEN, SEO, SEP, and SER. The detection and quantification limits of the assays were 3 and 4 ng/mL for SED and 1.5 and 3 ng/mL for SER, respectively.
Enterotoxin concentration was measured with rSED and rSER as standards, using a 4-parameter logistic curve fit. Concentrations of recombinant enterotoxins were determined with the Bicinchoninic Acid Kit (Sigma-Aldrich). Data analysis was carried out using GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA).
RNA extraction and RT-qPCR
Bacteria for RNA isolation were collected at 5, 8, and 24 h of growth. RNA extraction, purification, cDNA synthesis, and reverse transcription-quantitative polymerase chain reaction (RT-qPCR) were performed as described by Schubert et al. (2016). Primers used are listed in Table 2. rpoB, a housekeeping gene found to be stably expressed in milk, was used for cDNA normalization (Valihrach et al., 2014). Transcript levels relative to rpoB were calculated according to Pfaffl (2001). Data analysis was carried out using Bio-Rad CFX Manager software.
PCR efficiency for each primer pair was previously determined on genomic DNA from respective reference S. aureus strains by running serial 10-fold dilutions of the template (data not shown). Amplification efficiencies for all used primer sets were between 96% and 104%.
Statistics
Statistical significance of the results was assessed using the U Mann–Whitney test; p < 0.05 was considered statistically significant. Statistical analyses were performed using Statistica version 12 (StatSoft, Inc., Kraków, Poland).
Results
Bacterial growth
The growth curves were determined in milk and meat juice for five randomly selected S. aureus strains. S. aureus FRI1151m strain exhibited a significantly lower growth rate than the other strains in milk at all time points (p < 0.05). During growth in meat juice, higher bacterial counts were determined in S. aureus 56 strain at 24 and 32 h (p < 0.05) and in S. aureus 237 strain at 3 and 48 h (p < 0.05). Lower cell numbers were recorded for S. aureus 238 strain at 3, 8, and 24 h (p < 0.05) and for S. aureus FRI1151m strain at 32 and 48 h of culture (p < 0.05) (Fig. 1). All the 17 S. aureus strains reached 8.3–9.1 log CFU/mL at 24 h and 8.4–9.3 log CFU/mL at 48 h of culture in milk. During growth in meat juice, they reached 7.7–9.4 log CFU/mL at 24 h and 8.2–9.5 log CFU/mL at 48 h. Bacterial counts determined at 24 h were significantly higher in cultures conducted in milk than in meat juice (p < 0.05). No significant differences were noted in strains cultured both in milk and meat juice at 48 h.
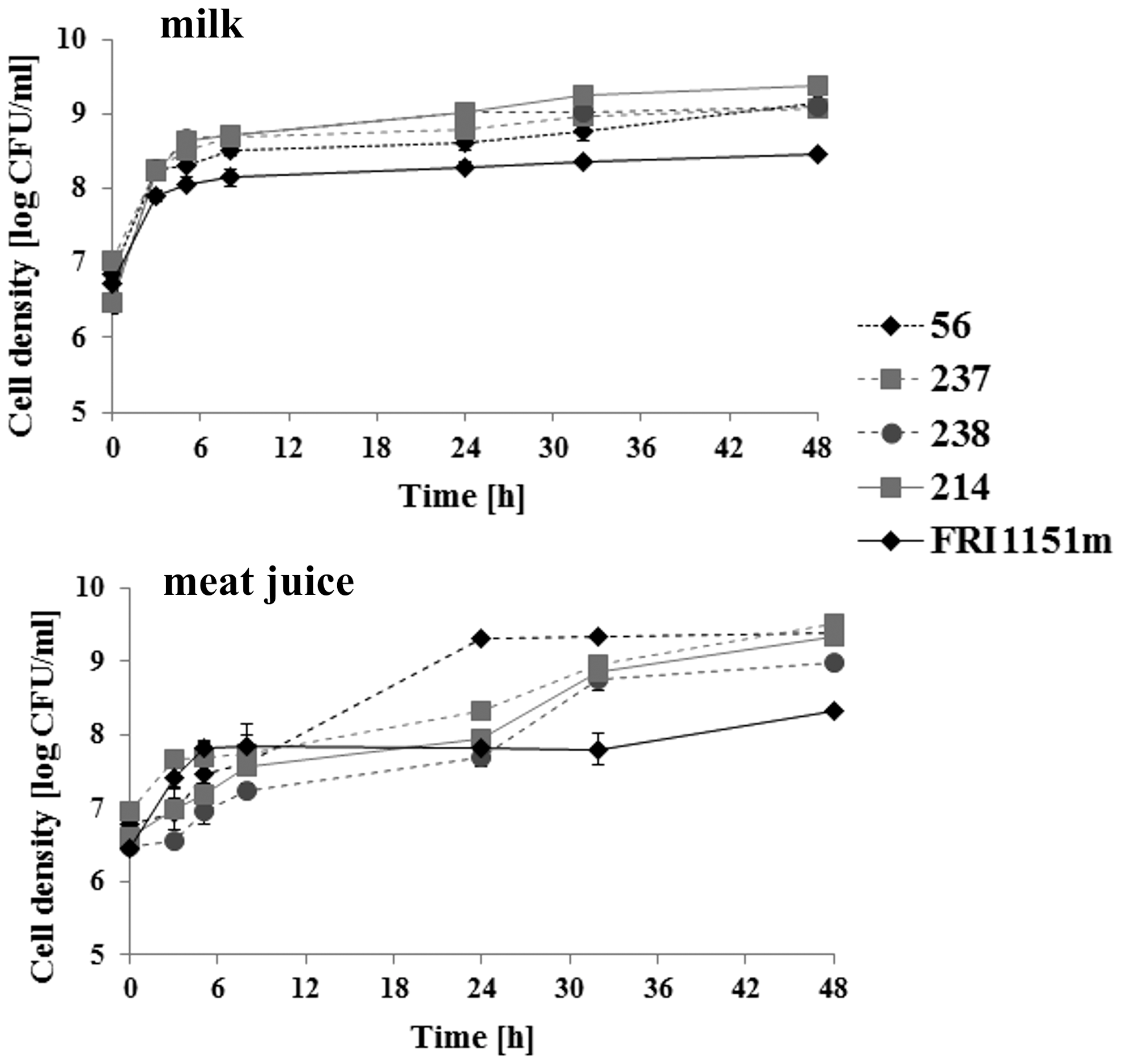
Growth of Staphylococcus aureus strains producing high levels of staphylococcal enterotoxin D (SED) in milk, that is, FRI1151m and 214 (marked with ______), and randomly selected weak SED producers, that is, 56, 237, and 238 (marked with - - - -), determined in milk and meat juice.
In S. aureus strains grown in milk, the initial pH value of 6.86 was unaltered until 48 h of growth. The pH of meat juice decreased from 6.22 at the beginning of culture to 5.51–6.20 at 5 h of growth and subsequently increased reaching 6.38–8.11 at 24 h and 6.75–9.03 at 48 h. The pH values at 24 and 48 h of bacterial growth in milk and meat juice are listed in Table 3.
Production of SED by S. aureus strains
Twelve of 17 S. aureus sed- and ser-positive strains, whose gene content is presented in Table 1, produced relatively small amounts of SED in milk with mean levels of 29 ng/mL at 24 h and 49 ng/mL at 48 h of culture. Five strains (FRI1151m, 214, 427, 453, and 553) secreted significantly higher amounts of SED in milk (p < 0.005) with mean levels of 1.2 μg/mL at 24 h and 2.4 μg/mL at 48 h. They were designated as strong SED producers. Significantly more SED was produced when S. aureus strains were grown in meat juice (p < 0.05). Strong producers of SED in milk secreted SED in meat juice with mean levels of 247 μg/mL at 24 h and 422 μg/mL at 48 h of growth. These amounts were significantly higher than for the remaining strains (p < 0.005), which produced SED in meat juice with mean levels of 683 ng/mL at 24 h and 980 ng/mL at 48 h.
Strong SED producers secreted between 65 and 459 times and between 77 and 538 times more SED in meat juice than in milk at 24 and 48 h, respectively, whereas the remaining S. aureus strains produced between 6 and 171 times and between 8 and 141times more SED in meat juice than in milk at 24 and 48 h, respectively (Table 3).
The bacterial numbers reached by tested strains in meat juice at 24 and 48 h as well as in milk at 24 h were not related to higher or lower SED expression. In turn, weak SED producers cultured in milk grew to higher densities at 48 h than strong SED producers (p < 0.05).
Production of SER by S. aureus strains
Tested S. aureus strains produced moderate amounts of SER in milk with mean levels of 111 ng/mL at 24 h and 206 ng/mL at 48 h of growth. Among them, 2 strains expressed less than 100 ng/mL of SER at 48 h, 11 strains produced between 100 and 200 ng/mL, and 4 strains secreted more than 200 ng/mL in milk. Fourteen S. aureus strains produced significantly higher levels of SER in meat juice after 24 and 48 h (p < 0.005). SER production by these strains in meat juice ranged from 2.7 to 14.6 μg/mL after 24 h and 3.4 to 27.3 μg/mL after 48 h. They produced between 17- and 232 times and between 15 and 269 times more SER in meat juice than in milk at 24 and 48 h, respectively. Three S. aureus strains secreted less than 74 ng/mL of SER in meat juice (Table 3).
The cell numbers reached by tested strains in meat juice and milk at 24 and 48 h were not related to higher or lower SER expression.
Expression of sed and ser genes and selected genes involved in their regulation
The level of transcripts was assessed in all five S. aureus strains producing the highest levels of SED in milk, that is, S. aureus FRI1151m, 214, 427, 453, and 553, and six randomly selected weak SED producers, that is, S. aureus 56, 237, 238, 266, 281, and 508.
sed transcripts were detected in all tested S. aureus strains. A rise of relative level of sed RNA was observed between 8 and 24 h of culture in milk in S. aureus FRI1151m, 281, and 553 (p < 0.05). The highest sed levels during bacterial growth in meat juice were noted at 5 h in S. aureus FRI1151m and 281 and at 8 h in S. aureus 214 and 281 (p < 0.05) (Fig. 2).
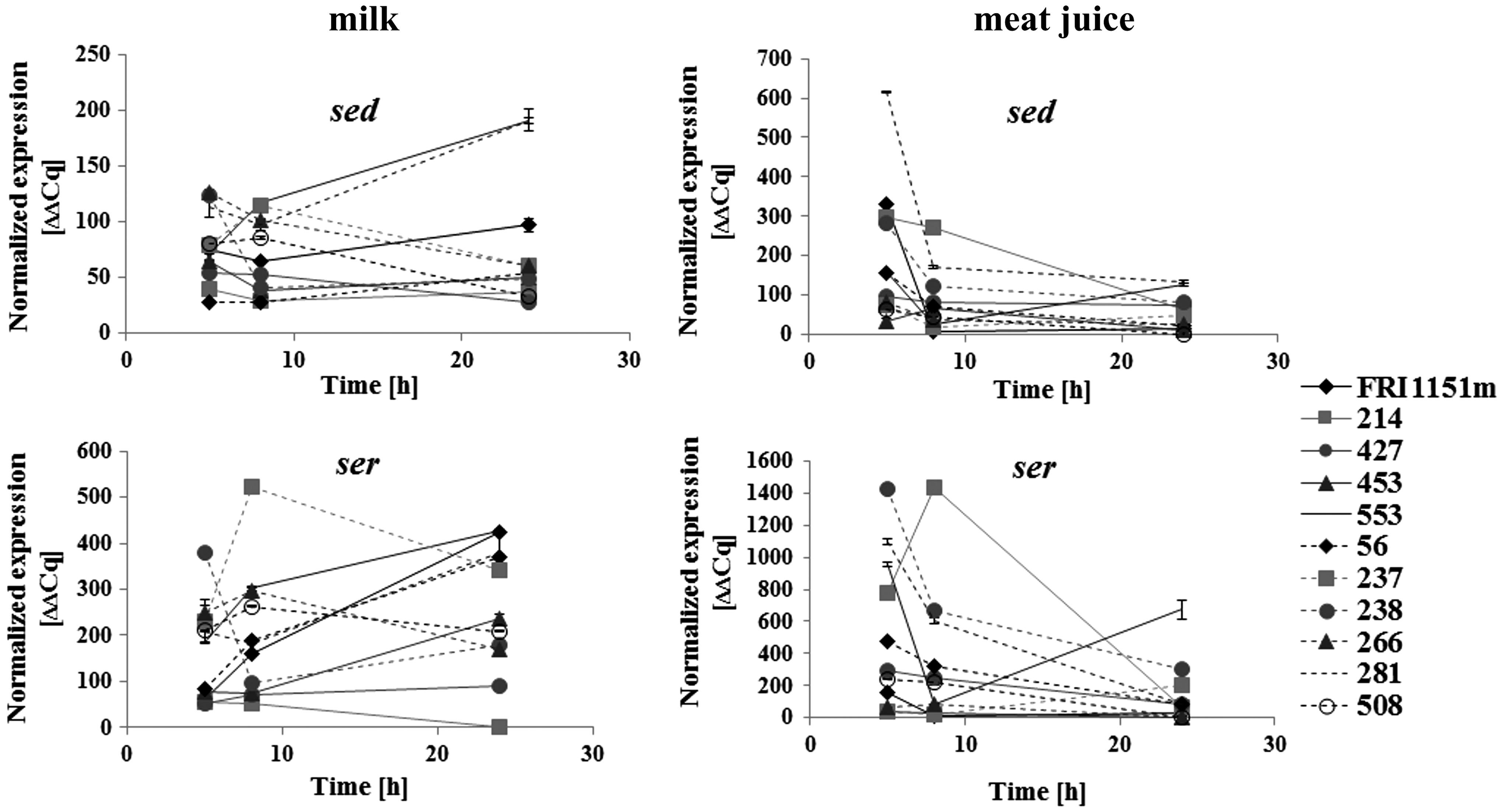
Relative sed and ser ribonucleic acid (RNA) levels in strong SED producers in milk, that is, Staphylococcus aureus FRI1151m, 214, 427, 453, and 553 strains (marked with ______), and randomly selected weak SED producers, that is, S. aureus 56, 237, 238, 266, 281, and 508 strains (marked with - - - -), determined in milk and meat juice.
The relative level of ser was upregulated between 5 and 8 h of growth in milk in S. aureus FRI1151m, 56, 237, and 553 and between 8 and 24 h in S. aureus FRI1151m, 56, 238, 281, and 553 (p < 0.005). There was a significant downregulation of ser RNA between 5 and 8 h of culture in meat juice in S. aureus FRI1151m, 56, 238, 281, and 553 (p < 0.05) and between 8 and 24 h in S. aureus 214, 238, and 281 (p < 0.005). Relative level of ser transcript was the highest at 24 h in S. aureus 553 grown in meat juice (p < 0.05) (Fig. 2).
The highest agrA levels during bacterial growth in milk were observed at 5 h in S. aureus 56, 237, and 453 (p < 0.005). There was a significant upregulation of agrA RNA between 8 and 24 h of culture in milk in S. aureus 281 (p < 0.05). The relative level of agrA was downregulated from 5 to 8 h of growth in meat juice in S. aureus 56, 214, 237, 238, and 281 (p < 0.005), followed by a further drop up to 24 h in S. aureus 214 (p < 0.05). Contrarily, the level of agrA RNA was upregulated between 8 and 24 h of culture in meat juice in S. aureus 56, 281, and 427 (p < 0.005) (Table 4).
Indicates points where significant changes in gene expression were noted (p < 0.05).
The relative level of sarA was upregulated from 8 to 24 h of growth in milk in all tested strains, except S. aureus 427 and 508 (p < 0.005). The expression of sarA RNA in S. aureus strains grown in meat juice revealed a similar pattern as in cultures conducted in milk. Most strains, except S. aureus 214, 453, and 508, showed significant upregulation of sarA RNA between 8 and 24 h of growth (p < 0.005), whereas a drop in sarA level was noted in S. aureus 214 and 453 (p < 0.005) (Table 4).
The rot level was downregulated from 5 to 8 h of culture in milk in S. aureus 238 (p < 0.05) and from 8 to 24 h in S. aureus 266 (p < 0.05). There was a significant increase in rot level between 8 and 24 h of growth in milk in S. aureus 56, 214, 281, and 453 (p < 0.05). The highest levels of rot during bacterial growth in meat juice were observed at 5 h in S. aureus FRI1151m and 281 (p < 0.005). Relative level of rot was upregulated from 8 and 24 h of culture in meat juice in S. aureus 281, 427, and 553 (p < 0.005) (Table 4).
The level of hld was upregulated between 8 and 24 h of growth in milk in all tested strains, except S. aureus FRI1151m, 56, and 266 (p < 0.005), in which the hld level was unaltered. Relative level of hld RNA increased from 5 to 8 h of culture in meat juice in S. aureus 214 (p < 0.05) and decreased in S. aureus 56, 237, 281, 508, and 553 (p < 0.05). There was a significant upregulation in hld level between 8 and 24 h of growth in meat juice in S. aureus 281 (p < 0.05) and downregulation in S. aureus 214, 238, and 453 (p < 0.05). hld transcripts were not detected in S. aureus 266 grown in milk and S. aureus FRI1151m and 266 cultivated in meat juice (Table 4).
Relative level of sigB was upregulated from 5 to 8 h of culture in milk in S. aureus 238 and 266 (p < 0.05). There was a drop in sigB level between 8 and 24 h of growth in milk in S. aureus 214 (p < 0.05) and its increase in S. aureus 237 and 281 (p < 0.005). The highest levels of sigB during bacterial growth in meat juice were observed at 5 h in S. aureus 214 and 281 (p < 0.005). The level of sigB RNA was upregulated from 8 to 24 h of culture in meat juice in S. aureus 281, 453, and 553 (p < 0.005) and downregulated in S. aureus 56, 214, 237, 238, 266, and 508 (p < 0.005) (Table 4).
Discussion
Milk and milk products are considered to be of particular significance as an staphylococcal enterotoxin (SE) source. In the European Union, only some milk-derived products are examined for enterotoxin content when the number of coagulase-positive staphylococci exceeds 105 CFU/g in a product (
SED is a well-characterized toxin, and strains that produce this toxin have been involved in numerous SFP cases (Wieneke et al., 1993; Kérouanton et al., 2007). Since SED and SER are encoded on the same plasmid in S. aureus, contribution of both emetic toxins to SFP is not easy to define. According to Omoe et al. (2003), specific S. aureus Fukuoka clones harbor plasmids from which the sed gene is absent. These strains can secrete up to 200 ng/mL of SER in broth culture and were implicated in SFP case in Japan (Omoe et al., 2003). It could thus be suggested that SER can contribute to food poisoning independently of SED. However, further study by the same team demonstrated that the Fukuoka strains encode two more emetic toxins, namely SES and SET, which complicate interpretation of the role of SER in SFP (Ono et al., 2008).
Milk has previously been observed as an unfavorable environment for expression of staphylococcal enterotoxins (Valihrach et al., 2013, 2014; Hunt et al., 2014; Schubert et al., 2016). Most studies published to date reported on staphylococcal strains carrying the sec gene. Based on these results, it was suggested that SEC production in milk should not account for high food safety risk (Hunt et al., 2014). We recently demonstrated significant downregulation of SEC and SEH expression in milk in a number of S. aureus strains harboring the seh gene (Schubert et al., 2016).
In this study, we demonstrate that other staphylococcal toxins, namely SED and SER, are expressed by S. aureus strains at lower levels in milk when compared with meat juice. Maximal SED concentration in milk produced by these strains did not exceed 100 ng/mL, while SER did not exceed 400 ng/mL. However, some strains included in our collection were able to secrete over 2 μg/mL of SED in milk. Meanwhile, strains producing low amounts of SED in milk were able to secrete up to 1.3 μg/mL of SED in meat juice, while strong SED producers secreted more than 650 μg/mL of SED into this medium.
Strong enterotoxin producers were also identified within SEH-producing strains, and high-level SEH production was related to a decrease of milk pH during culture, which was not observed in weak SEH producers (Schubert et al., 2016). High-level SED production, observed in a number of our strains, was not related to modification of milk pH by S. aureus up to 48 h of culture. Relationship of milk pH and enterotoxin production seems not to apply to all toxins, indicating different regulation of their expression. The pH of cultures carried out in meat juice consequently decreased at 5 h of growth, then increased up to 48 h, reaching different values depending on the strain.
The decline of pH in our cultures can result from an accumulation of fermentative end products (Regassa et al., 1992). Weak acids can pass the bacterial membrane and dissociate in the cytoplasm, which results in decrease of internal pH. To withstand unfavorable conditions, S. aureus can modify the pH of its environment. It was speculated that it can be achieved by metabolizing acids and producing diacetyl, pyrazine, and ammonia (Rode et al., 2010). Thus, the pH variation observed in our meat juice cultures can result from differences in levels and types of end products secreted by a given strain.
Expression of virulence genes in S. aureus, including enterotoxins, is controlled by transcriptional regulators such as the sarA family and sigB, as well as two-component systems, for example, agr and saeRS (Novick, 2001). We examined expression of five genes previously shown to be involved in sed gene regulation; however, we could not establish a clear relationship between the enterotoxin protein level and expression of investigated regulators in strong and weak SED producers. Moreover, in a number of our strains, enterotoxin RNA profiles seemed not to reflect respective protein levels. It was recently shown that sed RNA level may not be convergent with SED level, suggesting that yet unexplored ways of enterotoxin regulation may exist (Sihto et al., 2016).
Using mutational analysis of global regulators, Sihto et al. (2016) also demonstrated that SED expression may not be tightly controlled by Agr and that the effect of global regulators on SED may vary between strains. Schelin et al. (2011) demonstrated that enterotoxin production can be altered by the food matrix. Nonetheless, very few authors reported on molecular changes in S. aureus grown in milk. Lammers et al. (2000) investigating the response of S. aureus to the milk environment indicated four groups of genes specifically expressed in milk. They were involved in cell wall synthesis, nucleotide synthesis, transcriptional regulation, and carbohydrate metabolism. No changes in genes potentially involved in enterotoxin regulation were mentioned.
Conclusions
We demonstrated that in a number of tested S. aureus strains, SER was produced at higher levels than SED in milk and meat juice. With most strains tested in this study, production of SED and SER was significantly decreased in milk when compared with meat juice. However, certain S. aureus strains were still able to secrete considerable amounts of SED and SER in milk. No clear relationship of enterotoxin protein level and expression of known regulators could be established. Altogether, our results indicate that enterotoxin production in meat is likely to pose higher risk for food safety than enterotoxin production in milk, suggesting that an area of routine food control should be expanded. Nevertheless, food safety risk related to enterotoxin production in milk should still be considered important.
Footnotes
Acknowledgments
This project was financially supported by the National Science Centre, Poland, on the basis of decision DEC-2012/05/B/NZ9/03343. Costs of publication were supported by Wrocław Center of Biotechnology, program Leading National Research Center (KNOW) for the years 2014-2018.
Disclosure Statement
No competing financial interests exist.