
Abstract
The present study analyzed the prevalence and molecular characterization of Campylobacter at different processing steps in poultry slaughterhouses to determine where contamination mainly occurs. A total of 1,040 samples were collected at four different stages (preprocessing cloacal swabs, postevisceration, postwashing, and postchilling) in two processing plants. Campylobacter was detected in 5.8% (15 of 260) of the cloacal swabs and in 13.3% (104 of 780) of the processing samples. In both plants, the sampling points with the greatest contamination rates were after evisceration (20.5% and 15.4% for plants A and B, respectively) and significantly decreased after chilling (p < 0.05, from 20.5% to 10.9%) in plant A and after washing (from 15.4% to 2.9%) in plants B. In the result, however, the reduction in Campylobacter contamination was achieved through the sequential processing procedures in both plants. Campylobacter loads (>103 colony-forming units [CFUs]/mL) also decreased from 41.7% at evisceration to 20.0% in final carcasses. The genetic relationships of isolates were analyzed by the automated repetitive sequence-based polymerase chain reaction (rep-PCR) system, and the rep-PCR banding pattern was found to be unrelated to the processing plants, species, sampling point, or sampling day. As the gap in the intervention efficacy remains between plant A and B despite several consistencies, a national program for monitoring critical processing stages in poultry processing plants is recommended for the successful exportation of Korean-processed white mini broiler meat.
Introduction
C
In the United States, the regulatory approach toward foodborne pathogen control is focused primarily on poultry processing plants. For pathogen reduction, the U.S. Department of Agriculture introduced a new performance standard for Campylobacter in July 2011 as a risk management tool (FSIS, 2011; USDA, 2011). This quantitative risk assessment test for Campylobacter has been recommended by the Food and Agriculture Organization, World Health Organization, and Codex Alimentarius as a tool supporting risk management to harmonize international trade in poultry products (Nauta et al., 2009). However, any performance standard for Campylobacter has not been established in South Korea, and so, Campylobacter contamination has not been controlled in foods of animal origins processed in poultry processing plants (Yoo et al., 2014).
This study was conducted as an initial baseline survey for the introduction of new criteria for Campylobacter control at poultry processing plants in South Korea, before the exportation of Korean-processed white mini broiler meat. The primary objectives were to obtain comparative information on the distributions of Campylobacter prevalence, quantitative loads at the different poultry processing stages. In addition, the genetic relationship of isolates was assessed using the DiversiLab™ automated repetitive sequence-based polymerase chain reaction (rep-PCR) system.
Materials and Methods
Processing plant operations
This study was conducted in two poultry processing plants (plant A and B) between April and November 2012. As a prerequisite for the exportation of Korean-processed white mini broiler meat to the United States, the two plants were complying with Hazard Analysis and Critical Control Point (HACCP) requirements in accordance with the provisions of Article 9 of the Livestock Products Sanitary Control (LPSC) Act and the quarantine should be conducted in accordance with the provisions of Article 42 of the Prevention of Contagious Animal Disease (PCAD) Act. Both plants have some differences in the processes applied: Plants A and B had processing capacities of 500,000 and 50,000 birds per day, respectively. The carcasses were washed out with water pressure of 200 kPa in plant A and 304 kPa in plant B. The other difference was the chilling process: Plant A utilizes a water tank system added with NaClO followed by air chilling (also called “combi chilling”) and plant B relies on carcass cooling exclusively through water chilling added with NaClO (also called “spin chilling”).
Sample collection
Samplings were conducted at four main stages during poultry processing: I, cloacal swabs from live birds at the slaughter line; II, postevisceration; III, postwashing; and IV, postchilling (Fig. 1). At each plant, sample collections were conducted daily over the course of 5 months (132 visits per each plant). A total of 260 cloacal swabs were sampled from the individual chickens before they were processed. For the remaining three stages of bird processing, 780 carcasses were removed from the line at random for each specimen and immediately placed in a sterile plastic bag. Afterward, each chicken carcass was mixed with 400 mL of buffered peptone water (BPW; Oxoid, Hampshire, United Kingdom) in a sterile plastic bag and rinsed by shaking for 1 min; 50 mL of the rinsate constituted the sample. The rinse and swab samples were transported on ice packs to the laboratory for microbiological analyses.
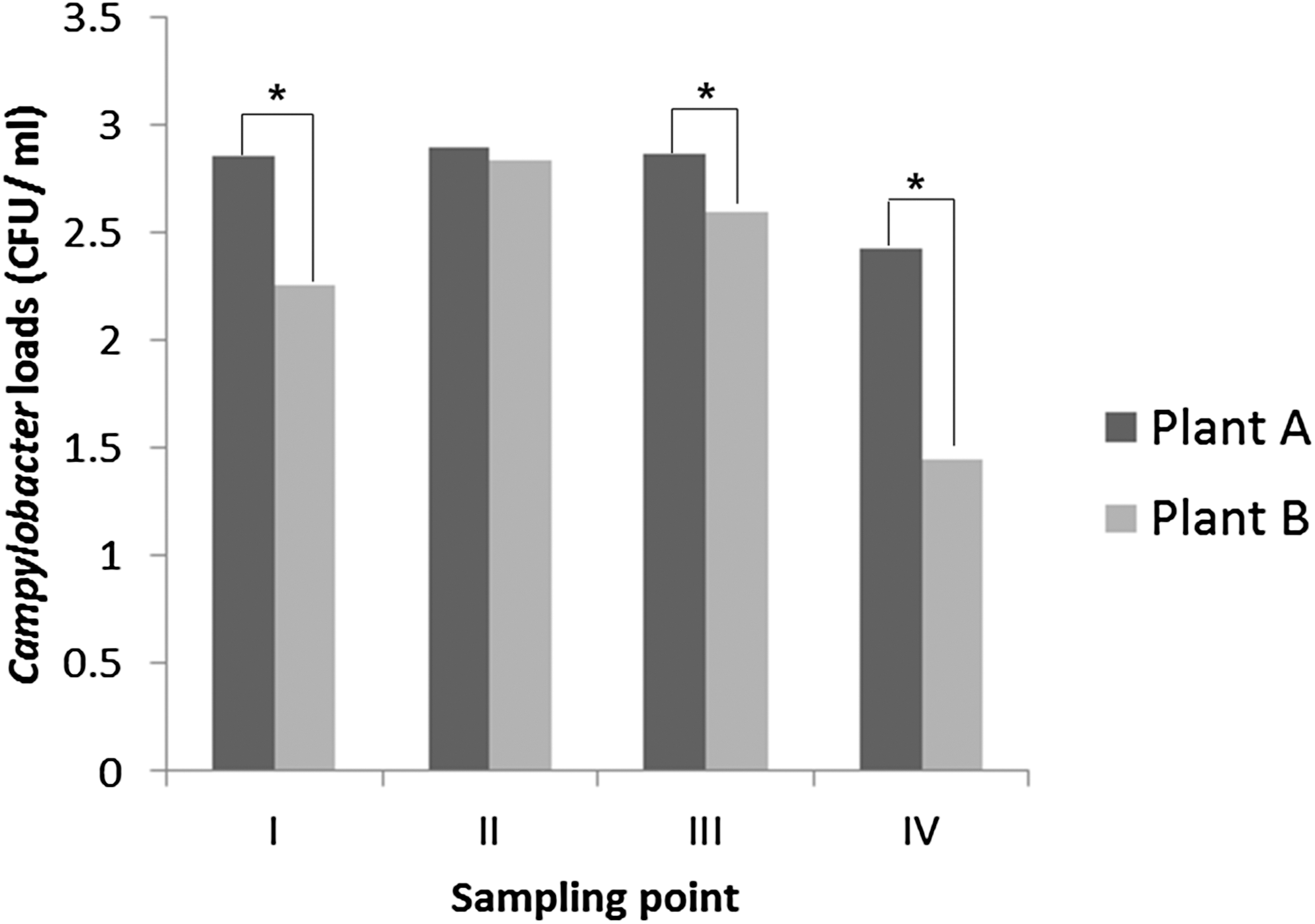
Distribution of Campylobacter loads at each poultry processing stage. Campylobacter loads were transformed as mean log10 CFUs/mL (±SD) in cloacal swab or carcass rinses taken from each sampling point. Roman numerals indicate the four sampling points in the slaughter process: I, fecal swabs on live birds; II, postevisceration; III, postwashing; IV, postchilling. Asterisks (*) indicate significant differences between plants A and plant B (p < 0.05). CFUs, colony-forming units; SD, standard deviation.
Campylobacter isolation
All experimental procedures for isolating and identifying Campylobacter were conducted following the USDA MLG 41 protocol recommended by the U.S. Food Safety and Inspection Service (USDA, 2013). For pre-enrichment, cloacal swabs were suspended in 10 mL of BPW and incubated at 37°C for 24 h. For chicken carcass samples, 30 mL of the rinsate was combined with the same volume of double-strength, blood-free Bolton's enrichment broth (Oxoid) and then incubated at 42°C ± 1°C for 48 ± 2 h in a microaerobic atmosphere (5% O2, 10% CO2, and 85% N2) generated using a BBL GasPak anaerobic system (Becton Dickinson). After pre-enrichment, a loopful of each enriched sample was streaked onto BBL Campy-Cefex agar (Becton Dickinson) and incubated at 42°C for 48 h. The suspected colonies (up to five per plate) were studied by Gram staining and tested for catalase and oxidase production, motility, and indoxyl acetate hydrolysis.
Campylobacter enumeration
All Campylobacter cultures were performed according to the culture method (MLG 41) recommended by the U.S. Food Safety and Inspection Service (USDA, 2013). Enumeration of Campylobacter spp. was performed by spread plating 1 mL of fecal and carcass rinse samples from the first dilution onto four Campy-Cefex plates (0.25 mL/plate) and additionally 0.1 mL from each dilution onto each of two Campy-Cefex plates (duplicates). Plates were incubated for 48 h at 42°C under microaerobic conditions, and presumptive positive colonies were enumerated. The mean colony number on four (or two) plates for each dilution was calculated and colony counts were converted to log10 colony-forming units (CFUs)/mL. Suspected colonies were confirmed as Campylobacter and species typed using multiplex PCR, as described below.
Multiplex PCR assay
Multiplex PCR detection was used to identify Campylobacter jejuni, Campylobacter coli, and Campylobacter lari species. Briefly, DNA extraction was performed using the UltraClean Microbial DNA Isolation Kit (MoBio Laboratories, Solana Beach, CA), according to the manufacturer's instructions. Specific detection of the Campylobacter genus was based on PCR amplification of the 16S rRNA gene using oligonucleotide primers, as described previously (Wang et al., 2002). All DNA extracts were examined for the presence of C. jejuni, C. coli, and C. lari species based on PCR amplification of the C. jejuni hipO, C. coli glyA, and C. lari glyA genes, respectively, as well as the C. jejuni 23S rRNA gene. DNA amplification was performed in a Biometra T-Personal thermal cycler (Biometra GmbH, Germany) using an initial denaturation step of 95°C for 6 min followed by 30 cycles of amplification (denaturation at 95°C for 0.5 min, annealing at 59°C for 0.5 min, and extension at 72°C for 0.5 min), ending with a final extension at 72°C for 7 min. The amplified products were electrophoresed in a 1.5% agarose gel, stained with SafeView™ (Applied Biological Material, Inc., Richmond, Canada), and photographed under UV light using a BioRad Molecular Imager® GelDoc™ XR (BioRad Laboratories). The positive control strains were represented by C. jejuni ATCC 33560, C. coli ATCC 33559, and C. lari ATCC 35221.
rep-PCR DNA fingerprinting
All the isolated strains were further characterized for molecular subtyping. Genomic DNA was amplified by PCR using the DiversiLab™ Campylobacter Kit (BioMérieux) and characterized using an automated DiversiLab rep-PCR system to evaluate the genetic similarity of Campylobacter isolates. The thermal cycling parameters and PCR conditions of the rep-PCR assay were in accordance with the manufacturer's instructions. The amplified products were analyzed using a 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA), and a dendrogram was constructed. Computer-generated virtual gel images were prepared using the DiversiLab system and subtyping banding patterns were determined using Pearson's correlation statistical methods. Isolates were classified based on the similarities between the rep-PCR banding patterns.
Data analysis
The number of positives between samples obtained during the same poultry processing stage at plants A and B was compared in pairs using Fisher's exact test in GraphPad InStat software (GraphPad Software, Inc., San Diego, CA). Results are reported as the mean and standard deviation. The data were analyzed using one-way analysis of variance in SPSS (version 15, SPSS, Chicago, IL). Differences between plants were determined using Tukey's honestly significant difference test at a significance level of p < 0.05.
Results and Discussion
Prevalence of Campylobacter at the four main stages of chicken processing
The prevalence of Campylobacter at each processing stage is presented in Table 1. Individual cloacal swabs yielded the same prevalence of 5.8% (9 of 156 in plant A; 6 of 104 in plant B) for Campylobacter. The prevalence of Campylobacter in both plants was highest at the evisceration stage (20.5% in plant A; 15.4% in plant B); these rates decreased as carcasses were further processed into the washing and chilling step. However, significant reduction showed at different processing stages: Before and after the chilling stage in plant A (p < 0.05; from 20.5% to 10.9%) and before and after the washing stage in plant B (p < 0.05, from 15.4% to 3.8%).
Values followed by different lowercase letters (a,b) in the same column are significantly different (p < 0.05). Values followed by different uppercase letters (A,B) in the same row are significantly different (p < 0.05).
The highest prevalence of Campylobacter after the evisceration stage is consistent with the findings of the previous studies (Figueroa et al., 2009; Hue et al., 2010; Sasaki et al., 2014), suggesting that the evisceration step should be controlled as the most critical point. Generally, systematic decontamination procedures have been known to be effective approaches to reduce pathogens such as Campylobacter from carcass surfaces (Hue et al., 2010). However, previous studies have reported inconsistent results with respect to the intervention efficacy of washing (Guerin et al., 2010). Our results also showed different results (significant decrease only in plant B but no change in plant A) in the prevalence of Campylobacter before and after the washing step. This difference may be related to the water pressure of the two plants; carcasses in plant A were processed with water supply (water pressure of 200 kPa) and carcasses in plant B were processed with tap water (water pressure of 304 kPa). It was reported that the efficacy of washing is variable and depends on several factors, including the initial load of pathogens, type of system, washing pressure, water temperature, and time (Dickson and Anderson, 1992). In the result, however, the reduction in Campylobacter contamination was achieved passing through the chilling process, which is reported to be the most critical stage for preventing bacterial contamination (Sanchez et al., 2002; Zhang et al., 2011). This finding is consistent with the results of Hinton et al. (2004), who reported lower prevalence of Campylobacter in postchill carcass rinses. Our results indicate that the use of chlorinated water during immersion chilling process can be effective in reducing microbial contamination of broiler carcasses in processing plants, as reported in previous studies (Northcutt et al., 2003; Berrang et al., 2007; Loretz et al., 2010).
Enumeration of Campylobacter at the four main stages of chicken processing
The distributions of Campylobacter loads in culture-positive samples are shown in Table 2. Campylobacter abundances in cloacal swabs from live birds ranged from 1.7 to 3.4 log10 CFUs/mL. At the postevisceration stage, the percent of positive samples containing >1000 CFUs/mL significantly increased (p < 0.05), compared to that in the range of <1000 CFUs/mL. The mean Campylobacter load in the carcass rinses sampled during the evisceration process was 3.0 log10 CFUs/mL, which was significantly higher than the mean load (1.4 log10 CFUs/mL) in postchill carcass rinses (p < 0.05, Fig. 1). Previous studies have demonstrated significant reductions in Campylobacter loads during the chilling stage of processing; Berrang et al. (2007) observed reductions of 2.23 log10 in Campylobacter loads between the re-hang and postchill carcass rinses. Northcutt et al. (2003) also observed overall reductions of 1.3 log10 for Campylobacter in carcass rinses before and after chilling. In this study, the change of Campylobacter loads in the range of >103 CFUs/mL also indicated that chilling systems are effective in reducing Campylobacter contamination of carcasses, showing a reduction from the evisceration stage (41.7%) to the final carcasses (20.0%). Especially, the mean Campylobacter loads in culture-positive rinses in plant B decreased by 1.40 log10 CFUs/mL during processing, compared to a corresponding reduction of 0.6 log10 CFUs/mL in plant A.
Values followed by different lowercase letters in the same column are significantly different (p < 0.05).
CFUs, colony-forming units.
Campylobacter species
The overall distribution of Campylobacter species was as follows: C. coli (52.9%) were predominant, followed by C. jejuni (21.0%) and C. lari (17.6%). However, the proportions of Campylobacter species differed between plants A and B: C. coli was predominant (56.7%), followed by C. jejuni (22.2%) and C. lari (13.3%) in plant A, while C. coli was predominant (41.4%), followed by C. lari (31.0%) and C. jejuni (17.2%) in plant B (Table 3).
The distribution of total Campylobacter species was similar to that reported in Ireland, Italy, and Spain, where C. coli was the most commonly isolated species (EFSA, 2011). However, in most investigations conducted in various other countries, C. jejuni was identified as the most common species in chicken samples (Jørgensen et al., 2002; Lahuerta et al., 2011; Garin et al., 2012). The difference in species predominance may depend on local environmental conditions, sampling techniques and methods, seasonal conditions, and laboratory methodologies (Yang et al., 2014). The samples contaminated with two or three different species accounted for approximately 8.4% of Campylobacter-positive samples, where co-contamination by C. jejuni and C. coli was more frequent than that by other species.
Molecular subtyping of isolates
Based on their rep-PCR banding patterns, Campylobacter strains exhibited high variation and no significant relationships to the species or the processing steps (data not shown). Therefore, the rep-PCR profiles of C. coli strains most frequently isolated in this study were representatively presented among all Campylobacter species identified (Fig. 2). The dendrograms of 47 C. coli showed the low-similarity index value, nor did it appear to distinguish samples originating from different sampling points or processing plants.

Dendrogram and virtual gel image of DiversiLab™ automated rep-PCR fingerprinting analysis. The analysis included 47 Campylobacter coli isolates obtained from various poultry processing stages, of which 40 were from plant A and 7 were from plant B. Roman numerals indicate the four sampling points in the slaughter process: I, fecal swabs on live birds; II, postevisceration; III, postwashing; IV, postchilling. rep-PCR, repetitive sequence-based polymerase chain reaction.
Our results indicate that the ability to type Campylobacter species (C. jejuni, C. coli, and C. lari) using rep-PCR appeared to be limited although the genetic similarity in rep-PCR patterns has been highly observed depending on serotype in previous studies with Salmonella isolates (Wise et al., 2009; Hyeon et al., 2011, 2013). The low genetic relativeness among isolates in this study may relate to variations in the sampling date, sampled flocks, or difference of sample source (feces or carcasses). Although the automated rep-PCR technique has been applied for the genotyping and strain identification of several pathogens, including Neisseria meningitidis, Candida spp., and Archaea (Cleland et al., 2008), it has not been extensively tested with Campylobacter isolates (Behringer et al., 2011). Therefore, a wide comparison of subtyping methods for Campylobacter species is considered necessary to validate the efficacy of rep-PCR typing for Campylobacter.
Conclusion
The present study was performed in cooperation with two broiler processing plants to evaluate changes in Campylobacter loads during carcass processing. Although the prevalence observed may not be representative of the entire broilers processed in Korea, the results are meaningful as baseline data to investigate the management practices of the surveyed processing plants and to introduce new criteria for Campylobacter control in white mini broiler processing plants. For the trade promotion of Korean-processed white mini broiler meat, additional researches through consecutive and countrywide survey are needed to develop and quantify the effectiveness of pathogen control methods. Further studies may make advances in the estimation of Campylobacter prevalence and quantitative loads in commercial poultry processing plants in South Korea and expand our understanding of the benefits and limitations of various typing methods for epidemiological studies of Campylobacter spp.
Footnotes
Acknowledgments
This work was supported by the Animal and Plant Quarantine Agency (No. B-FS03-2011-12-01) and the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (No. 2015R1A2A2A05001288).
Disclosure Statement
No competing financial interests exist.