
Abstract
Shiga toxin–producing Escherichia coli (STEC) are important emerging foodborne human pathogens. Ruminants are the main animal reservoir of STEC currently known, and meat can become contaminated at different stages of the production chain. The aim of this work was to subtype and establish the epidemiological relatedness of non-O157 STEC strains isolated from ground beef and the environment in butcher shops before (evaluation stage, 2010–2011 period) and after (verification stage, 2013) implementing improvement actions. Sixty-eight non-O157 STEC strains were tested for eae, saa, ehxA, iha, efa1, toxB, subAB, cdt-V, astA, aggR, and aaiC genes, and stx 1 and stx 2 variants were determined. Pulsed-field gel electrophoresis (PFGE) was carried out with XbaI and XmaJI. From the 68 strains, 92.6%, 75.0%, 58.8%, 53.5%, 10.3%, 7.3%, and 4.4% were positive for iha, ehxA, subAB, saa, cdt-V, astA, and eae, respectively. All strains were aggR/aaiC-negative. PFGE showed that 19 strains grouped in 9 clusters and 41 showed unique XbaI patterns. During the evaluation stage (2010–2011), we identified clonal strains in different samples, circulating clones in different butcher shops, and more than one different strain in the same butcher shop. The bovine origin of meat and its manufacturing process could not ensure the total absence of all non-O157 STEC serotypes in this foodstuff. Most strains isolated during the evaluation (2010–2011) and verification (2013) stages did not exhibit a genotypic profile associated with human disease. It is necessary to conduct periodic reviews of the new epidemiological information and verify that the analyses of non-O157 STEC in food are appropriate to identify strains affecting the population.
Introduction
S
Non-O157 STEC strains are responsible for over 60% of the STEC infections in the United States of America; O26, O45, O103, O111, O121, and O145 are the serogroups most frequently associated with severe disease worldwide (Atkinson et al., 2012; EFSA, 2013; Gould et al., 2013). In Argentina, non-O157 STEC strains are responsible for 25.1% of STEC infections, and O145, O121, O26, O174, O111, and O8 are the main serogroups isolated from ill patients (Rivas et al., 2010).
STEC strains are characterized by the presence of stx genes that codify for Shiga toxin (Stx), the main STEC virulence factor (Calderwood and Mekalanos, 1987; Etcheverria and Padola, 2013; Kruger and Lucchesi, 2015). Intimin, an adhesin encoded by the eae gene in the locus of enterocyte effacement pathogenicity island of the chromosome, is involved in gut colonization. Although it is not clear which combination of markers defines a pathogenic STEC strain, stx 2/eae is associated with a higher risk of more serious illness (EFSA, 2013).
In addition to stx and eae, STEC strains could harbor a complex set of genetic determinants, including other toxin and adherence genes such as saa (STEC autoagglutinating adhesin), lpf (long polar fimbriae), iha (IrgA homolog adhesin), efa (enterohemorrhagic E. coli factor for adherence), toxB (plasmid virulence gene of E. coli O157), cdt-V (cytolethal distending toxin), subAB (subtilase cytotoxin), astA (enteroaggregative E. coli heat-stable toxin 1), and ehxA (enterohemolysin) (USDA, 2012; Etcheverria and Padola, 2013; Franz et al., 2015).
Ruminants are the main animal reservoir of STEC currently known. Although contaminated foodstuffs derived from cattle such as ground beef are responsible for human illness (Hussein, 2007; Martin and Beutin, 2011; EFSA and ECDC, 2015), outbreaks attributed to leafy vegetables, dairy products, fruits, and other meats are more severe than those caused by beef, probably due to a change in strain virulence and host susceptibility by patient age and sex (Heiman et al., 2015).
Pathogen bacteria can be transferred to beef carcasses during slaughtering, dressing, chilling, or cutting at slaughterhouses, and from the equipment to meat at retail stores (Perez-Rodriguez et al., 2010; Vogeleer et al., 2014). While meat mincing machines and handlers contaminated with STEC have been associated with meat cross-contamination in retail stores (Papadopoulou et al., 2012), pathogen cross-contamination between food and the retail store environment has been described as a factor responsible for increased foodborne disease risk (Sirsat et al., 2014). In the Autonomous City of Buenos Aires, Argentina, zero-tolerance criteria to STEC in meat products from supermarkets and fast food shops were applied between 2004 and 2008.
The aim of this work was to subtype and establish the epidemiological relatedness of non-O157 STEC strains isolated from ground beef and the environment in butcher shops before and after implementing improvement actions.
Materials and Methods
In October 2010, a pilot program called “Healthy Butcher Shops” was conducted in the city of Berisso, Buenos Aires, Argentina (Leotta et al., 2016). A comprehensive risk assessment was performed in 172 raw ground beef provided by 10 slaughterhouses (A–J) and 672 environmental samples collected from 86 butcher shops (B1–B86) during the evaluation (2010–2011) and verification (2013) stages of the study.
During 2011, to implement improvement actions in butcher shops, we performed collective training meetings for butchers, customized trainings for handlers, and individual counseling at the stores, providing recommendations about facilities, good manufacturing practices, sanitation standard operating procedures, raw food handling, and meat preservation (Leotta et al., 2016). Sixty-eight non-O157 STEC were isolated from ground beef and environmental samples (meat tables, knives, mincing machines, and manipulator hands; Leotta et al., 2016). All strains were stored at −70°C in the strain collection of IGEVET (Instituto de Genética Veterinaria “Ing. Fernando Noel Dulout,” UNLP-CONICET LA PLATA) for further characterization.
Strains were grown in 4 mL brain heart infusion (BHI) broth (Biokar, Zac de Ther, France) at 37°C ± 1°C for 18–24 h. This culture was streaked onto BHI agar plates and incubated overnight at 37°C ± 1°C. A single colony from each strain was selected and grown overnight in 4 mL BHI broth at 37°C ± 1°C.
All strains were tested for the presence of the eae, saa, ehxA, iha, efa1, toxB, subAB, cdt-V, and astA genes (Galli et al., 2010). The aggR and aaiC genes were detected by real-time PCR (ISS, 2013). The stx 1 variant was determined with primers stx1a-F1, stx1a-R1, stx1c-F1, stx1c-R1, stx1d-F1, and stx1d-R1 (Scheutz et al., 2012). Genotyping of the stx 2 variant was performed with primers VT2-c, VT2-d, VT2v-1, and VT2v-2 by restriction fragment length polymorphism analysis of the DNA fragments obtained by PCR (Tyler et al., 1991).
Molecular subtyping of non-O157 STEC strains was performed with pulsed-field gel electrophoresis (PFGE) using the 1 d (24–26 h) PulseNet standardized laboratory protocol (CDC, 2013). Restriction digestion of DNA in agarose plugs was carried out with XbaI and XmaJI (BlnI) as primary and secondary enzymes, respectively (Thermo Scientific). PFGE images of gels were obtained by MaestroGen slider® imager (Maestrogen, Inc., Nevada). TIFF image analysis was carried out with BioNumerics, version 6.6 (Applied Maths, Sint-Martens-Latem, Belgium) using the dice coefficient and the unweight pair group method with arithmetic mean (UPGMA) to generate dendrograms with 1.5% band matching tolerance. Two or more isolates were grouped in a cluster when they showed identical XbaI-PFGE pattern (100% similarity).
Results
Non-O157 STEC strains harbored different variants of the Shiga toxin gene, namely stx 2c(vh-b) (N = 23), stx 2a (N = 22), stx 1a/stx 2a (N = 12), stx 2a/stx 2c(vh-b) (N = 4), stx 2c(vh-a) (N = 3), stx 1a (N = 3), and stx 1a/stx 2c(v-hb) (N = 1) (Table 1).
-, strain that showed unique XbaI-PFGE pattern; B, Butcher shop; E, Evaluation stage (2010–2011); ID, strain identification number; N, number of strains isolated; N/A, not applicable (strain excluded from PFGE analysis due to DNA degradation); PFGE, Pulsed-field gel electrophoresis; S, Sampling stage; STEC, Shiga toxin–producing Escherichia coli; V, Verification stage (2013).
All strains were eae-negative (95.6%), except for three that were eae-positive (4.4%). The ehxA gene was carried by 51 isolates (75.0%). Forty-four ehxA-positive strains were saa-positive as well, whereas two saa-positive strains were ehxA-negative. The most prevalent putative adhesin was the one encoded by the iha gene, where 63 strains were positive (92.6%).
Gene sequences related to subAB, cdt-V, and astA toxins were present in 40 (58.8%), seven (10.3%), and five (7.3%) strains, respectively. One subAB-positive strain was also eae-positive and belonged to serotype O109:H25. The astA-positive strains belonged to serotypes O174:H21 (N = 3) and O178:H19 (N = 1), whereas one strain was nontypeable. None of the O174:H21 astA-positive strains could be subtyped by PFGE because of bacterial lysis. All strains were aggR/aaiC-negative. Only O26:H11 (eae-positive) harbored the efa1 and toxB genes, while the rest of the strains were efa1 and toxB-negative. The complete genotypic characterization is presented in Table 1.
Sixty-eight non-O157 STEC isolates were analyzed by XbaI-PFGE. Eight strains were excluded from the analysis because their DNA was degraded three times, even when thiourea was added to the running buffer (Table 1). The XbaI-PFGE UPGMA dendrogram is shown in Figure 1. Fifty XbaI-PFGE patterns were obtained, with a 54.8% similarity. Nineteen strains grouped in 9 clusters and 41 strains showed unique XbaI-PFGE patterns. Clusters I, II, III, IV, V, and VIII grouped strains from the 2010 to 2011 evaluation stage, whereas clusters VI, VII, and IX included strains of the 2013 verification stage.
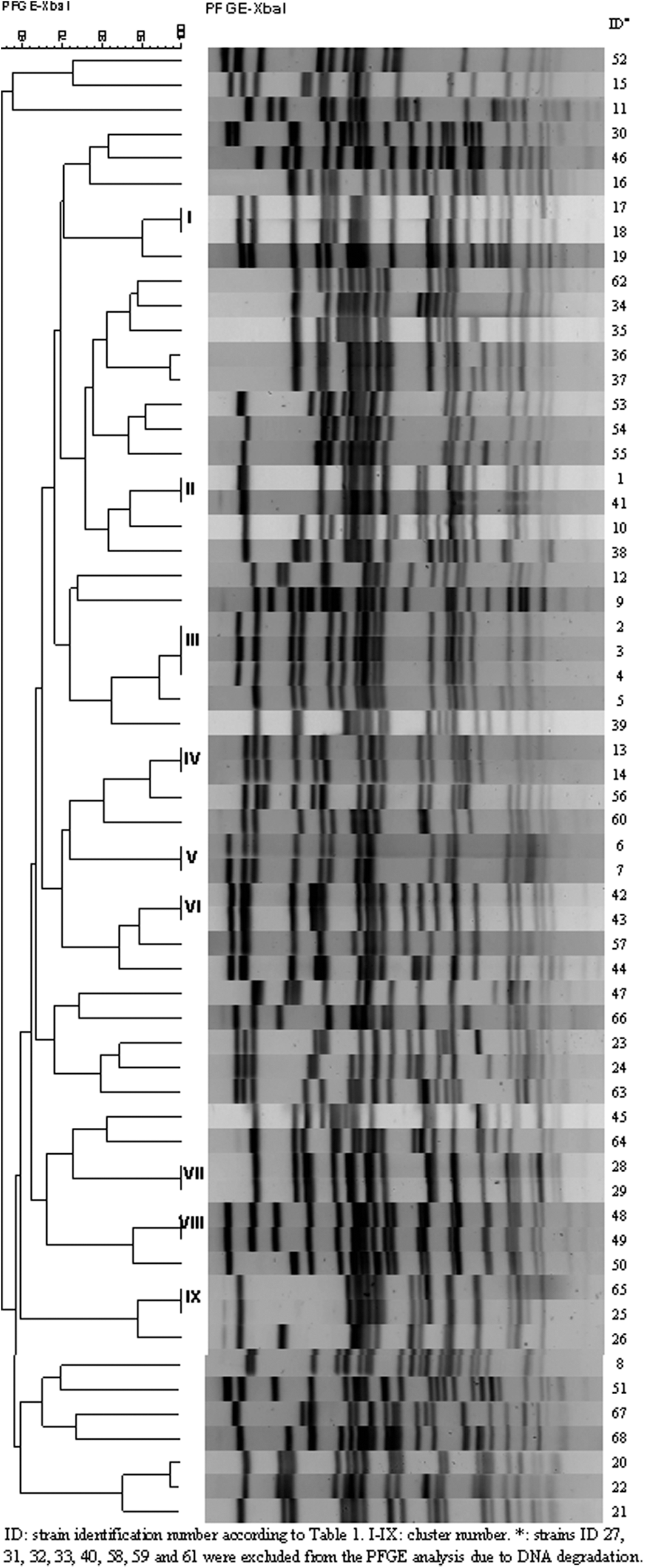
XBaI-PFGE UPGMA dendrogram, sample type, sampling stage, serotype and genotype of 60 non-O157 STEC strains isolated from butcher shop. PFGE, pulsed-field gel electrophoresis; STEC, Shiga toxin–producing Escherichia coli; UPGMA, unweight pair group method with arithmetic mean.
Strain clusters were as follows: I, one strain from ground beef and one from meat mincing machine of B85; II, one strain from manipulator hands of B5 and one from mincing machine of B2; III, one strain from meat table of B23, one from knife of B29, and one from meat table of B57 (the meat supplier of these three butcher shops was slaughterhouse A); IV, one strain from meat table of B27 and one from mincing machine of B63 (the meat supplier for both butcher shops was slaughterhouse A); V, one strain from ground beef of B20 and one from ground beef of B83; VI, one strain from meat table and one from knife of B61; VII, one strain from meat table and one from mincing machine of B61; VIII, one strain from ground beef of B81 and one from meat table of B82; and IX, two strains from ground beef of B49.
Strain ID44 isolated from ground beef of B61 (Fig. 1) showed one band of difference with strains of cluster VI isolated from meat table and knife of the same butcher shop. Due to the epidemiological relationship between these strains, they could be considered as a clone with a common origin (Tenover et al., 1995). Strains ID 36 and ID 37 isolated from meat table and knife of B76, respectively, showed one band of difference between them (Fig. 1), and could be considered clones (Tenover et al., 1995). Strains from cluster II and VII were distinguished by cutting with Bln1-PFGE.
Discussion
The principle of the bacterial subtyping approach is to compare the distribution of subtypes in potential sources (e.g., animals and food) with the subtype distribution in humans (EFSA, 2013). In this work, we investigated the prevalence of the new scheme of virulence genes proposed by the EFSA plus four putative adhesins and three toxin genes in 68 non-O157 STEC strains isolated from 86 Argentinian butcher shops in a previous study (Leotta et al., 2016). The strains were also molecularly subtyped to compare the distribution of potential clones.
According to the EFSA (EFSA, 2013), there is not a unique combination of markers that define pathogenic STEC strains. However, stx 2/eae and stx 2/aaiC/aggR strains were associated with a higher potential risk of causing severe illness than other combinations of virulence genes (EFSA, 2013). In Argentina, 94.3% of STEC isolates from acute diarrhea, bloody diarrhea, or HUS cases harbored stx and eae genes (Rivas et al., 2010).
In this study, all non-O157 STEC strains were aaiC/aggR-negative and 96.5% were eae-negative. Although most of the strains isolated corresponded to serotypes associated with illness in Argentina, they did not exhibit a genotypic profile associated with human disease, probably accounting for the absence of HUS cases in Berisso during the 2010–2013 study period (Galli et al., 2016; Leotta et al., 2016). Epidemiological surveillance should be intensified with the aim of identifying emerging strains with new pathogenic potential and determining whether they are associated with foodstuffs.
Among the eae-positive strains showing a genotypic profile associated with severe disease, we found one O26:H11 and one O109:H25 strain. O26:H11 (stx 1/eae) was the only strain showing a close association of the efa and toxB genes with intimin-positive strains. It also exhibited a highly potential risk of causing diarrhea or HUS/HC, as was described in previous studies (Cergole-Novella et al., 2007). Strain O109:H25 isolated from mincing machines was cdt-V and subAB-positive.
Several authors proposed the possible role of the CDT-V and SubAB toxins in the infection pathogenesis caused by STEC (Bielaszewska et al., 2004; Talbot et al., 2005). Cergole-Novella et al. (2007) and Galli et al. (2010) observed these toxin genes in intimin-negative strains. However, in the present study, subAB was found in one O109:H25 eae-positive strain. Despite this serotype was previously associated with human and animal origin (Beutin et al., 2004; Krause et al., 2005), none of those strains carried the subAB gene. Consequently, the potential clinical relevance is also unknown.
Food equipment has been recognized as an important vehicle of contamination throughout the meat supply chain (Gounadaki et al., 2008; Perez-Rodriguez et al., 2010). During the evaluation stage (2010–2011), clonal non-O157 STEC strain in ground beef and mincing machine of B85 (cluster I) was detected by XbaI-PFGE. The poor sanitation of the mincing machine could have been the origin of ground beef contamination.
Biofilm presence and transference of pathogenic bacteria between food and utensils and equipment and handlers have been previously described in different stages of the meat production chain (Phang and Bruhn, 2011; Papadopoulou et al., 2012; Vogeleer et al., 2014). In addition, circulating clones were found in different butcher shops, such as those grouped in clusters III, IV, V, and VIII. Some of these clusters were composed of strains from butcher shops with the same meat supplier, and the source of contamination could have been at the slaughterhouse. Futures studies of meat suppliers could confirm whether they are a source of STEC contamination.
During the 2010–2011 evaluation stage, more than one different non-O157 STEC strain in B20, B29, and B81 was identified. The presence of several nonclonal strains in the same butcher shop, even in the same sample, could be due to different sources of contamination such as slaughterhouse, transport, handlers, water, and vectors (Persad and LeJeune, 2014; Vogeleer et al., 2014).
After the implementation of improvement actions, the percentage of STEC strains corresponded to the main non-O157 serotypes associated with illness cases in Argentina. Such percentage was reduced from 43.9% to 13.5% (Leotta et al., 2016), and none of the clonal strains observed at this stage were the same as those isolated during the evaluation stage.
The lack of a defined combination of virulence factors required for STEC-associated clinical infection is not conclusive (EFSA, 2013). From the first published report of STEC serotypes in 1980, 1152 different serotypes have been described (Bettelheim and Goldwater, 2014). Most of them are eae/aaiC/aggR-negative and belong to serogroups other than O157, O26, O103, O145, O111, and O121. Thus, the prevailing uncertainty lies in their ability to cause disease or not (EFSA, 2013). We consider that due to the bovine origin of meat and its manufacturing process, it is impossible to assure the total absence of all non-O157 STEC serotypes in this foodstuff. In addition, there are no available methodologies for the isolation of all STEC serotypes in food (Brusa et al., 2016).
Conclusion
Zero-tolerance criteria to non-O157 STEC in ground beef and environmental samples were applied in the isolation of all STEC strains from butcher shops of Berisso, Argentina. However, most strains isolated during the evaluation (2010–2011) and verification (2013) stages of the study did not exhibit a genotypic profile associated with human disease. These results indicate that the efforts to apply zero-tolerance criteria to non-O157 STEC in meat without a risk analysis that includes food and consumers would be excessive. Cross and multiple contamination in butcher shops as well as of circulating clones among butcheries in the city of Berisso were identified using PFGE. In this sense, slaughterhouses were recognized as a possible common source of STEC contamination in butcher shops.
Thus, studies of meat suppliers could be useful to know more about their importance as a source of contamination in the meat production chain. Strengthening epidemiological surveillance, based on molecular analysis, could give an insight into the clinical implications of virulence genes and allow the classification of STEC strains more efficiently according to risk. It would be necessary to conduct periodic reviews of the new epidemiological information and verify that the analyses of non-O157 STEC performed in foods are appropriate to identify strains affecting the population. The continuous work of all members of the food chain will allow a better approach to know the prevalence of non-O157 STEC strains in foodstuffs affecting both inhabitants and the epidemiological relationship between patients and meat.
Footnotes
Acknowledgments
The authors acknowledge the support of Cristian Adriani for sample collection at butcher shops. They also thank A. Di Maggio for editing the article.
Disclosure Statement
No competing financial interests exist.