
Abstract
Two hundred fresh produce samples (cucumber, carrots, herbs, leaf lettuce, and ready-to-eat mixed salad leaves) were obtained from retail in northern Germany in 2015. These were investigated for microbial contamination and the presence of foodborne pathogens, including Listeria monocytogenes, Salmonella serovars, presumptive Bacillus (B.) cereus, and Shiga toxin–producing Escherichia coli using culture-dependent (enrichment, plating on selective media) and -independent (real-time polymerase chain reaction [PCR]) techniques. Overall, our results showed that the fresh produce samples generally showed high mean aerobic mesophilic bacterial counts of between 7 and 8 log10 cfu/g. However, there was also a considerable variation in total aerobic bacterial counts between different product samples. Although real-time PCR signals for pathogenic E. coli were detected in 14.0% of total samples analyzed, only one (0.5%) Shiga toxin–producing E. coli isolate of serotype O26:H11 was recovered from mixed salad leaves and contained stx1, stx2, and eae genes. Two L. monocytogenes isolates (1% of total samples) were recovered from packaged mixed salad leaves and belonged to PCR serogroups IIb and IVb, respectively. One Salmonella isolate (0.5%) was recovered after selective enrichment also from mixed salad leaves and it was identified as Salmonella Szentes serotype 16:k:1,2. Overall the incidence of foodborne pathogens on the northern German retail market in 2015 was very low.
Introduction
S
Because of the enterohemorrhagic/enteroaggregative Escherichia coli (EHEC/EAEC) epidemic in Germany in 2011, raw vegetable foods have come more into focus as a vehicle for the transmission of pathogens that can cause severe infections. A contributing factor is that these are often eaten raw (Berger et al., 2010) and are not subjected to preservation methods which have an impact on enhancing the food's safety. Fresh and fresh-cut vegetables are usually not processed in ways that will effectively eliminate human pathogens (Jung et al., 2014). Human pathogens that can be associated with produce include E. coli serovars O104:H4 and O157:H7 or other Shiga toxin–producing E. coli strains (STEC), Salmonella enterica ssp. enterica serovars, Campylobacter spp., as well as L. monocytogenes (Berger et al., 2010; Olaimat and Holley, 2012; Jung et al., 2014; Callejón et al., 2015). Probably the most serious outbreak of foodborne disease caused by a bacterial pathogen from a vegetable source in Europe was the EHEC/EAEC outbreak with E. coli serotype O104:H4 in Germany in 2011 (Buchholz et al., 2011).
This study aimed to investigate the incidence of foodborne pathogens on fresh produce obtained from retail in northern Germany in an effort to determine whether any specific recent problematic species or serovar could be identified. Furthermore, the study aimed to assess the microbiological quality of such products, as well as potential risk of foodborne infection associated with these products in this region.
Materials and Methods
Sampling
A total of 200 different vegetable products, including cucumber, carrots, herbs (mostly basil, parsley, chive, and dill), leaf lettuce (mostly romaine lettuce, lamb's lettuce, and arugula), and ready-to-eat mixed salad leaves (Table 1), were obtained from self-service counters of retail stores, local markets, and farmyard sales in northern Germany between January and October 2015. Samples originated from more than 40 different retail sources. Samples were transported to the laboratory and stored at 4°C. Microbiological analyses were carried out on the same day.
The minimum, maximum, and mean counts (incl. standard deviation) are shown.
minimum detection limit.
maximum detection limit.
lower than 5 log10.
above 5 log10.
lower than 3 log10.
above 7 log10.
LAB, lactic acid bacteria; SD, standard deviation.
Microbiological analyses
Microbiological analyses were carried out on the basis of the German official collection of methods (National Official Collection of Methods of Analysis and Sampling) on the basis of § 64 of German Food and Feed Act (LFGB, 2013), described below. For the detection and enumeration of microorganisms, 25 g samples were taken aseptically to include a central and an outside portion of the vegetable, placed in a sterile stomacher bag (VWR, Darmstadt, Germany) with 225 mL of buffered peptone water (BPW; VWR) (1:10 dilution) and homogenized in a stomacher (Stomacher 400; Seward, United Kingdom) for 120 s, 200 rpm at room temperature. In case of cucumbers and carrots, layers of ∼2 mm thickness were aseptically removed from the exterior area of the surface, and 25 g of these layers were transferred in a sterile stomacher bag.
For enumeration of bacteria, tenfold serial dilutions were performed using Buffered Sodium Chloride/Peptone Solution (VWR), and aliquots (0.1 mL) of the appropriate dilutions were plated out. Mesophilic aerobic counts were enumerated on plate count agar (PCA; VWR) incubated at 30°C for 72 h. Enterobacteriaceae were grown on VRBD agar (Merck, Darmstadt, Germany) at 37°C for 24 h, and all pink to red colonies larger than 1 mm were counted. E coli counts were determined on Brilliance™ E.coli/coliform Agar (CCA; Oxoid, Wesel, Germany) after 24 h at 37°C, and lactic acid bacteria (LAB) numbers were assessed on de Man, Rogosa, and Sharpe (MRS) agar (VWR) at 30°C for 48 h. Presumptive B. cereus was enumerated on Bacillus-Cereus-Agar (PEMBA; Sifin, Berlin, Germany) at 30°C for 48 h. All plates were incubated under aerobic conditions.
Pathogen detection
As only low pathogen counts were expected to occur in these products, a nonselective enrichment step and a second selective enrichment step were used for L. monocytogenes and Salmonella detection. For STEC, only the first nonselective enrichment step was carried out. For the first nonselective enrichment 25 g of sample were transferred into 225 mL BPW, homogenized, and incubated at 37°C for 18 h.
After nonselective enrichment, a volume of 0.1 ml BPW was directly plated out on CHROMagar™ STEC (VWR) and incubated for 24 h at 41.5°C for STEC detection (suitable to detect major STEC serovars, i.e., O26, O103, O45, O111, O121, O145, and O157). Presumptive STEC colonies (mauve colored) were genotyped by conventional multiplex-polymerase chain reaction (PCR) for their Shiga toxin and intimin genes according to Paton and Paton (1998) (Supplementary Table S1; Supplementary Data are available online at
For L. monocytogenes detection, 1 mL of BPW enrichment was transferred to 9 mL of L-PALCAM-Listeria selective enrichment broth (Merck) and was incubated at 37°C for 48 h. After that PALCAM broth was streaked out onto PALCAM-Agar (VWR) and Listeria-Selective-Agar (ALOA; VWR) (10 μL), and the plates were incubated at 37°C for 48 h. Suspected Listeria isolates were confirmed and serogrouped using multiplex PCR according to Doumith et al. (2004).
For Salmonella detection, a 0.1 mL of the BPW enrichment was added to 10 mL Rappaport-Vassiliadis Salmonella Enrichment Broth (RVS; VWR) and then incubated at 41.5°C for 24 h. To confirm the results, 1 mL BPW enrichment was also added to 10 mL Tetrathionate-Novobiocin Broth according to Muller-Kauffmann (MKTTn; VWR), then incubated at 37°C for 24 h. A loopful (10 μL) of each of the RVS and MKTTn enrichments were separately streaked out onto Xylose-Lysine-Deoxycholate-Agar (XLD; Oxoid) and Salmonella-Shigella Agar (SSA; Carl Roth, Karlsruhe, Germany), respectively, and the plates were then incubated at 37°C for 24 h. Suspected Salmonella isolates were confirmed and serotyped at the national reference center (NRC) for Salmonella at the Robert Koch Institute (RKI) (Wernigerode, Germany).
Real-time PCR detection of STEC
A real-time PCR targeting stx1, stx2, and eae genes was performed from all products in triplicate according to Pavlovic et al. (2010). For this, 1 mL of the incubated BPW was centrifuged for 5 min at 13,000 g, and the supernatant was removed. The cell pellets were washed twice with 1 mL distilled water, and the samples were centrifuged for 3 min at 13,000 g following each washing step. After the last washing step, the cells were suspended in 200 μL bi-distilled water, and DNA was extracted by ZR Fungal/Bacterial DNA MiniPrep™ (Zymo Research, Freiburg, Germany) according to the manufacturer's instructions. Five μL of isolated DNA was used for the real-time PCR in 25 μL reactions in a CFX96 Real-Time PCR System, using a 96-well plate format (Bio-Rad, München, Germany). The reaction mixtures contained iQ™-Multiplex Powermix (Bio-Rad), 0.3 μM of each primer, and 0.2 μM of each probe specific for stx1, stx2, and eae (Supplementary Table S1). The two-step PCR program consisted of an initial denaturation step for 7 min at 95°C, followed by 40 cycles of denaturation for 5 s at 95°C, and annealing ( = extension) for 30 s at 58°C. To prevent false positives, genomic DNA of E. coli O157:H7 EDL933 was used as positive control, and no-template-control was used as negative control.
Characterization of presumptive B. cereus sensu lato isolates
Presumptive B. cereus isolates were not obtained from 89 of the 200 vegetables sampled in this study. The remaining 111 samples contained presumptive B. cereus with typical characteristics on PEMBA medium (smooth nonrhizoid colony of blue color surrounded by a distinguishing egg yolk precipitate). However, isolates could not be obtained from all plates because of overgrowth with other microbiota, and therefore, isolates from only 75 independent samples were purified and characterized. The cultures were grown overnight in Caso Bouillon (VWR), and genomic DNA extraction was done using the ZR Fungal/Bacterial DNA MiniPrep™ Kit (Zymo Research). The 16S rRNA gene was amplified using 27F and 1492R primer (Wilmotte et al. 1993). The PCR amplification was performed in 25 μL using 0.1 mM concentration of each primer, 50 ng of genomic DNA, and DreamTaq Green PCR Master Mix (2X) (VWR) according to the manufacturer's protocols. The reaction products were sequenced in both directions. Sequences were analyzed using Geneious (v9.0.5; Biomatters Limited, New Zealand), and phylogenetic analysis was done using BioNumerics (v7.1, Applied Maths, Sint-Martens-Latem, Belgium) using pairwise alignment and unweighted pair group method with arithmetic mean clustering.
B. cereus sensu lato toxin genes were detected by multiplex PCR according to Ehling-Schulz et al. (2006). The strains B. cereus ATCC10987 (cytK), B. cereus DSM4312 (ces), B. cereus DSM2301 (nhe), and B. cereus MRI B200 (hbl) were used as positive control.
Data analyses
Data were exported from Excel 2010 (Microsoft, Redmond) to SigmaPlot 11 (Systat Software, Inc., San Jose) for analysis, including descriptive statistics, means, standard deviations, and box plots.
Results
The mean total aerobic mesophilic plate counts of the five different produce groups (cucumber, carrots, herbs, leaf lettuce, and ready-to-eat mixed salad leaves) were all ca. 6 to 7.5 log10 cfu/g, with only the mean counts for the cucumbers and herbs being lower than this value, but still above 6 log10 cfu/g (Fig. 1A). The results varied within the product between 3 and 5 log units, so that the lowest total aerobic counts were ∼4 log10 cfu/g (e.g., for a cucumber and a herb sample), while the highest counts were higher than 9 log10 cfu/g for several samples of leaf lettuce, herbs, and mixed salads (Fig. 1A and Table 1). The exceptions to this were cucumber and carrot, for which no samples showed counts higher than 8.3 or 8.4 log10 cfu/g, respectively (Table 1). Among the product groups, the majority of carrots, leafy lettuce, and mixed salad leave samples (50%, 42.5%, and 32.5%, respectively) showed highest counts between 7 and 8 log10 cfu/g (Table 1) and were therefore the product groups with the highest average microbial loads. Moreover, high percentages of these product samples also contained microbial loads between 6–7 log10 cfu/g (Table 1).
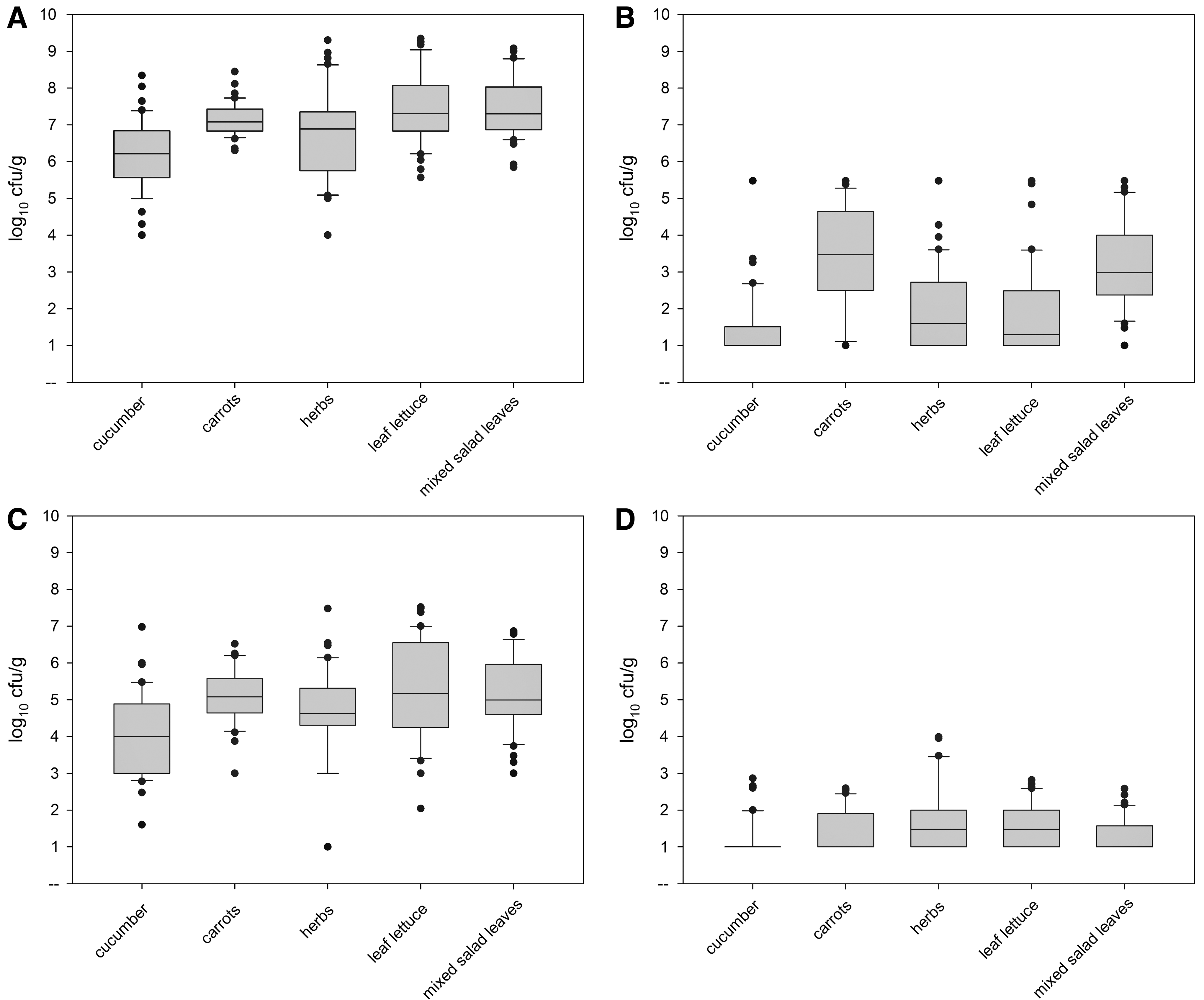
Aerobic, mesophilic
LAB counts determined on MRS agar showed variable counts both between products and between samples of individual products (Fig. 1B). The mean of LAB counts were the highest for carrots and mixed salad leaves at 3.4 and 3.1 log10 cfu/g followed by herbs, leaf lettuce, and cucumber with average counts ranging between 1.9 and 1.4 log10 cfu/g (Table 1). Similar to the total aerobic counts, the LAB counts of individual samples varied in a range of 5 log units or more (Fig. 1B). For example, for carrots, the highest LAB counts for a specific product sample was >5.4 log10 cfu/g, while the lowest product sample count was <1 log10 cfu/g (Fig. 1B and Table 1).
The mean counts of Enterobacteriaceae determined on VRBD medium were generally higher than LAB counts (Fig. 1B, C and Table 1). These counts were ca. 4 to 5 log10 cfu/g, but again the counts of different product samples showed considerable variations. For example, the counts of Enterobacteriaceae of the herb samples occurred in a span of almost 7 log units, ranging from 1 log10 cfu/g to almost 7 log10 cfu/g (Table 1).
Mean presumptive B. cereus counts on PEMBA agar were overall comparatively low, generally between 1 log10 and 2 log10 cfu/g (Fig. 1D and Table 1). In this case, the inter-product sample counts showed considerably less variation, of only about 2 to 3 log units, except for the case of herbs, for which the highest presumptive B. cereus count determined was 3.9 log10 cfu/g (Table 1). In Germany, there is a guideline for numbers of B. cereus in fresh produce only for mixed salads, which was defined by the German Society of Hygiene and Microbiology (DGHM) and which is not legally binding (
Using culture-dependent methods and direct plating, pathogens such as Salmonella, STEC, and L. monocytogenes can frequently not be detected, because of uneven distribution or due to their presence in low concentrations. In contrast to Salmonella and L. monocytogenes, the required infection dose to cause disease is considerably lower for STEC. For this reason, the samples were first enriched before pathogen detection by either culture-dependent (Salmonella, L. monocytogenes, and STEC) or culture-independent methods (STEC).
This way, PCR signals for pathogenic E. coli could be detected in 29 of the 200 (14.5%) produce samples (Supplementary Table S2). For eight of these (8/200 samples = 4%), only the eae gene was detected by real-time PCR, showing that these eight samples were probably indicative of enteropathogenic E. coli, while either stx genes alone or a combination of eae and stx1 and/or stx2 genes were detected in 21 other samples (21/200 samples = 10.5%), and they were indicative of the presence of STEC. Using real-time PCR as detection system, STEC signals were predominantly detected in herbs (6/40 samples = 15%), mixed salad leaves (5/40 samples = 13%), leaf lettuce (5/40 samples = 13%), and carrots (4/40 samples = 10%), while only one STEC positive signal was detected among the 40 cucumber samples (2.5%). Despite the rather frequent detection of STEC signals in products with real-time PCR, it was only possible to recover one STEC isolate directly from a selective agar plate in one sample (1/200 samples = 0.5%). The STEC isolate was recovered from a sample of mixed salad leaves (Supplementary Table S3), which was produced and packaged in Germany and was sold by a supermarket. The mesophilic aerobic plate count for this product was 8.1 log10 cfu/g, and the product showed an Enterobacteriaceae count of 4.8 log10 cfu/g. This isolate was sent to the NRL (BfR), where it was typed and determined to be an E. coli as belonging to the O26:H11 serotype. This isolate contained the eae, stx1, and stx2 genes and a typical light violet (mauve) colony color on CHROMagar™ STEC (VWR).
Two L. monocytogenes isolates (2/200 samples = 1.0%) were recovered from packaged mixed salad leaves (Supplementary Table S3) after enrichment and were PCR typed to belong to serogroup IIb (containing serovars 1/2b, 3b, and 7) and serogroup IVb (containing serotypes 4b, 4ab, 4d, and 4e), respectively (Doumith et al., 2004).
Only one Salmonella isolate (1/200 samples = 0.5%) was recovered from all samples following Salmonella specific enrichment. This isolate was obtained from mixed salad leaves (Supplementary Table S3) and showed typical Salmonella spp. characteristics of hydrogen sulfide production and colonies with black at the center on XLD agar. It was sent for serotyping to the NRC at the RKI in Wernigerode, Germany, and was identified as a Salmonella enterica ssp. enterica Szentes serotype 16:k:1,2. This isolate showed a very similar pulsotype to the Salmonella Szentes strains isolated from clinical samples and a sprout sample in Switzerland (Nüesch-Inderbinen et al., 2015), with an equal amount of bands and a highly similar band pattern (data not shown).
The 16S rRNA sequencing showed that all isolates except one clustered together closely at r > 99.5% with the type strains of B. cereus sensu lato (Fig. 2), identifying these isolates as belonging to this group. All of the isolates possessed the genes for at least one toxin [64 of 75 isolates possessed nhe (85%); 56 of 75 isolates hbl (75%); 28 of 75 isolates cytK (37%); and none possessed ces].

Dendrogram obtained by UPGMA of similarity value r of 16S rRNA gene sequences of 75 presumptive B. cereus isolates. The result of multiplex PCR for toxin genes is indicated for each strain (black square = positive); these results were not considered in the dendrogram construction. The dendrogram shows 74 of 75 isolates to group into the B. cereus sensu lato group. The isolate names indicate the origin of the independent isolates, K stands for herbs, M for carrots, B for leaf lettuce, G for cucumber, and MS for mixed salad leaves. Type strain was not tested for toxins. PCR, polymerase chain reaction; UPGMA, unweighted pair group method with arithmetic mean.
Discussion
Overall, our results showed that the specific fresh produce samples investigated in this study generally showed quite high means of aerobic, mesophilic bacterial counts between 7 and 8 log10 cfu/g. However, there was also a considerable variation in mesophilic aerobic bacterial counts between different samples of a specific product. Such variations in type and counts of microorganisms have been suggested to depend on the type of vegetable, environmental considerations, seasonality, and conditions under which a particular vegetable is grown (Rico et al., 2007). Levels of enterobacteria in this study were ca. 5 log10 cfu/g for most products and also showed considerable variation. Although E. coli was not selectively isolated or counted, these high enterobacteria counts indicated that pathogenic E. coli might be present. Therefore, the possible presence of STEC was determined using enrichment and selective plating, as well as by real-time PCR.
In this study, stx1 or stx2 genes alone or in combination with eae genes, indicative of STEC strains, could be detected with real-time PCR in ∼10.5% of the samples investigated. In contrast, only one STEC isolate from a mixed leaf salad sample (0.5% of total samples) could be cultured. This isolate was serotyped as O26:H11 and infections with E. coli O26:H11 strains from various sources occur in Germany in less than 80 cases per year (RKI, 2016). A STEC O26:H11 (stx1, stx2, and eae positive) strain caused a small outbreak in several European countries in 2016 (EFSA, 2016), and this study shows that mixed leaf salads can also be one source for their transmission. In this study, an incidence of 10.5% of STEC-positive samples could be determined using culture-independent methods. However, we did not have any information whether DNA from dead cells or phages was detected. In contrast, due to the fact that our method did not include an external amplification control, there remains a possibility of having obtained false negative results and of underreporting the incidences of STEC. When looking at culture-dependent detection, however, only one STEC isolate could be recovered, which is in line with the reported low incidences of STEC in these type of products (Johannessen et al., 2002; Johnston et al., 2005, 2006; Abadias et al., 2008; Bohaychuck et al., 2009).
In our study, nonselective enrichment followed by plating onto selective media, allowed us to isolate an O26:H11 serotype STEC. The large outbreak of EHEC/EAEC from sprouts in 2011 in Germany was an atypical EHEC outbreak, due to an enteroaggregative E. coli O104:H4 strain, which was atypical in that it appeared to have acquired the stx2 gene for Shiga toxin production and caused serious infections in young adults (Buchholz et al., 2011).
Only one Salmonella enterica ssp. enterica serotype Szentes isolate could be recovered from 200 investigated products (0.5% incidence). Salmonella is known to occur on fresh produce and is one of the major pathogens that can be isolated from these (Abadias et al., 2008; Berger et al., 2010; Olaimat and Holley, 2012; Tan et al., 2016). Relatively recently, Salmonella enterica serotype Szentes caused a 9-month outbreak in Switzerland in 2013 and 2014. One of the isolates involved was obtained from a sprout sample and it was indistinguishable from the human clinical isolates obtained during the outbreak. Thus, produce was considered to have been a possible source of infection (Nüesch-Inderbinen et al., 2015).
L. monocytogenes isolates could be recovered after enrichment from two mixed salad leave samples, respectively (2/200 = 1.0% incidence). PCR serogrouping showed that the two isolates belonged to serogroups IIb and IVb, respectively. European Food Safety Authority (EFSA [2015]) recently reported the European data for outbreaks with foodborne pathogens in 2014. A total of 12 Listeria outbreaks were reported by seven states, which were slightly higher than in previous years. Only in one of these outbreaks, a mixed salad was involved and consumption of this in a hospital/medical facility caused one death (EFSA, 2015). Stephan et al. (2015) reported on a larger outbreak with L. monocytogenes in 2013–2014, for which the source of Listeria was ready-to-eat salad. In our study, only a low incidence of L. monocytogenes was determined for fresh produce (1% incidence considering all products investigated), which was similar to reports of other studies (Johannessen et al., 2002; Johnston et al., 2005, 2006; Abadias et al., 2008; Olaimat and Holley, 2012; Korir et al., 2016).
B. cereus counts were generally quite low, that is, between 1 log10 and 2 log10 cfu/g. Nevertheless, all isolates contained at least one diarrheal toxin gene. The distribution of toxin genes among B. cereus sensu lato isolates in this study was similar as reported elsewhere, except for the fact that not all isolates encoded nhe, as was previously reported (Böhm et al., 2015). Interestingly, the gene for the emetic ces toxin could not be identified among any of the produce-associated B. cereus sensu lato isolates.
Conclusions
Generally, the results of our study showed that the incidence of foodborne pathogens in fresh produce on the market in northern Germany in 2015 was very low. This indicated that even though containing high mean numbers of mesophilic aerobic bacteria of ca. 7–8 log10 cfu/g, fresh produce on the German market appeared to be relatively safe for consumption as based on the low incidence of pathogens. The presence of toxin genes do, however, indicate that these products may pose a health risk if high numbers of toxigenic bacteria are present and sufficient levels of toxin are produced. The presence of pathogens, although at low numbers, also gives rise to caution and reinforces that we should be vigilant and regularly monitor this important food supply, to avoid potential similar disasters as the EHEC/EAEC outbreak with O104:H4 EHEC in 2011.
Footnotes
Acknowledgments
The authors gratefully acknowledge the technical assistance of Petra Horn, Gesa Gehrke, and Lena Niclausen. The authors also thank the National Reference Center for Salmonella at the Robert Koch Institute (RKI, Wernigerode) for serotyping the Salmonella isolate and the National Reference Laboratory for Escherichia coli of the Federal Institute for Risk Assessment (BfR, Berlin).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.