
Abstract
Salmonella Enteritidis remains a standout among the leading causes of foodborne diseases worldwide. Previous studies have demonstrated that a unique clonal group of Salmonella Enteritidis, named SE86, is involved in foodborne outbreaks in southern Brazil and is frequently identified among strains isolated from poultry. The aim of this study was to determine the influence of the isolation source (food products involved in salmonellosis outbreaks and poultry sources) on the phenotypic and molecular characteristics of Salmonella Enteritidis SE86. A biofilm formation assay, antimicrobial susceptibility test, polymerase chain reaction identification of virulence-associated genes, and phage type 4 (PT4) assessment were performed to characterize Salmonella Enteritidis SE86. The human strains presented less antimicrobial resistance than the poultry strains. Resistance to some substances was related to the isolation source of the strain. Strains of the same clonal group presented different biofilm production abilities. Biofilm formation was independent of the isolation source at all temperatures. Temperature influenced biofilm formation only by the poultry strains. Most of the investigated genes presented a high frequency and a regular distribution, regardless of the isolation source. The spvB, spiA, pagC, sipB, prgH, spaN, sitC, and lpfC genes were associated with the avian strains, whereas iroN was associated with the strains isolated from food products involved in salmonellosis outbreaks. Most strains belonged to PT4. No relationship was found between biofilm production and antimicrobial resistance or between the virulence profile and biofilm production or antimicrobial resistance.
Introduction
S
Previous studies have demonstrated close relationships between the Salmonella Enteritidis strains involved in foodborne outbreaks in southern Brazil during the past few years; this specific clonal group is named Salmonella Enteritidis SE86 (Geimba, 2005; Oliveira et al., 2009, 2012; Tondo et al., 2015). A recent study demonstrated the spread of this unique clonal group among strains isolated from poultry and outbreak food samples (Borges et al., 2017).
Resistance of Salmonella strains isolated from both humans and animals to the most commonly used antibiotics has increased over the past decade (EFSA and ECDC, 2015). Salmonella has also developed several other survival mechanisms in addition to antimicrobial resistance. For instance, biofilm formation favors bacterial survival in hostile environments, such as slaughterhouses and food-processing plants, and it is an important problem in public health due to the release of pathogenic microorganisms, such as Salmonella (Costerton et al., 1995; Hung and Henderson, 2009; Steenackers et al., 2012). Salmonella can invade host cells, and its pathogenesis and interaction with the host are complex phenomena that depend on several virulence factors encoded by a large number of genes distributed along its chromosome and in mobile genetic elements (Groisman and Ochman, 1996; Wallis and Galyov, 2000; Skyberg et al., 2006).
Characterization of the Salmonella Enteritidis SE86 is essential to better understand why the incidence of this specific clonal group has increased in southern Brazil since 1999. In this context, the aim of this study was to determine the influence of the isolation source on the phenotypic and molecular characteristics of Salmonella Enteritidis SE86 isolated from food products involved in salmonellosis outbreaks and poultry sources.
Materials and Methods
Bacterial strains
A total of 148 Salmonella Enteritidis strains isolated from food products involved in salmonellosis outbreaks (n = 80) and from poultry sources (n = 68), including chicken meat, internal organs, and cloacal and drag swabs, were selected for this study (Table 1). The strains were serotyped by the Oswaldo Cruz Institute Foundation (Fiocruz, Brazil). These strains were previously classified by polymerase chain reaction (PCR) ribotyping as belonging to the clonal group SE86 (Borges et al., 2017). The bacterial isolates were stored frozen at −80°C in brain heart infusion broth (Oxoid, Basingstoke, United Kingdom) supplemented with 15% glycerin (Synth, Diadema, Brazil).
Antimicrobial susceptibility test
Antimicrobial susceptibility was determined by the disk diffusion technique according to the Clinical and Laboratory Standards Institute method (CLSI, 2014a). The interpretation was based on the criteria described in the approved standards VET01-S2 (CLSI, 2014b) and M100-S26 (CLSI, 2016), as appropriate. An Escherichia coli (ATCC 25922) strain was selected to ensure the validity of the test. The following antibiotic disks (Oxoid) were used: amoxicillin (AMX), 10 μg; ceftiofur (TIO), 30 μg; ciprofloxacin (CIP), 30 μg; chloramphenicol (CHL), 30 μg; enrofloxacin (ENR), 5 μg; gentamicin (GEN), 10 μg; spectinomycin (SPT), 100 μg; sulfafurazole (SOX), 300 μg; sulfamethoxazole with trimethoprim (SXT), 1.25 μg/23.75 μg; and tetracycline (TCY), 30 μg. All strains classified as intermediate were considered nonsusceptible as determined by the WHO (2015). Strains that presented resistance to three or more classes of antimicrobials were considered multidrug resistant (Schwarz et al., 2010).
Biofilm formation assay
The inoculum preparation and biofilm formation assay using a microtiter plate method were performed as previously described (Borges et al., in press). Incubation was performed under the following different temperature conditions: 37°C, optimum temperature for Salmonella growth (Gast, 2008); 28°C, temperature for the expression of the extracellular matrix components (Steenackers et al., 2012); 12°C, required by the Brazilian sanitary service in cutting rooms of broiler processing plants (Brasil, 1998); and 3°C, the ideal average temperature for domestic refrigerators (da Silva et al., 2008). The cutoff optical density (OD) for the microtiter plate test was defined as three standard deviations above the mean OD of the negative control. The strains were classified into four categories: no biofilm producer (ODS ≤ ODC), weak biofilm producer (ODC < ODS ≤ 2 × ODC), moderate biofilm producer (2 × ODC < ODS ≤ 4 × ODC), or strong biofilm producer (4 × ODC < ODS) (Stepanović et al., 2004).
Detection of virulence-associated genes
DNA extraction was performed by heat treatment as described by Borges et al. (2017). PCRs were conducted to detect the presence of 27 virulence-associated genes in the Salmonella strains. The genes, their descriptions, and the respective references are summarized in Table 2. The cycling conditions and reaction mixtures (25 μL) are described in Table 3. All PCRs were performed with 2.5 μL of 10 × PCR buffer (Centro de Biotecnologia UFRGS, Porto Alegre, Brazil). The cycling program was performed in the Esco Swift MaxPro thermal cycler (Esco, Singapore). The amplified products were separated by electrophoresis in a 1.5% agarose gel and stained with ethidium bromide. The fragments were transilluminated with UV light. Mannheimia haemolytica ATCC 29694 and Salmonella Enteritidis ATCC 13076 were used as the negative and positive control, respectively, for all PCRs except for the cdtB gene, for which a strain of Salmonella Senftenberg (from our laboratory stock collection) was used as the positive control. In all PCRs, a mixture of all constituents of the PCR mixed without the addition of extracted DNA was used as a PCR control.
Forward (F) and reverse (R).
Reference for primer sequence.
PT4, phage type 4.
Reference for the reaction mixtures and cycling conditions.
PCR, polymerase chain reaction.
Determination of PT4 by PCR
Primers for PCR phage typing were specific for the amplification of a specific and conserved nucleotide sequence of PT4 by PCR and their sequences are summarized in Table 2. The cycling conditions and reaction mixtures (25 μL) are described in Table 3. PCR was performed as previously described. The positive control was a strain of Salmonella Enteritidis from our laboratory stock collection that was previously identified using a phage typing scheme standard as a PT4 strain.
Statistical analysis
The obtained data were subjected to statistical analysis using the PASW statistic software (IBM, Hong Kong). Descriptive statistics were used to determine the groupings of the samples according to biofilm formation, antimicrobial resistance, presence of virulence genes, and PT4. Student's t-test was performed to analyze biofilm production between origin sources and to determine the relationship between biofilm production, antimicrobial resistance, and the presence of virulence genes. The nonparametric chi-square (χ2) and Fisher's exact tests were used to compare antimicrobial resistance and the presence of virulence genes based on the isolate origins. The McNemar test was used to compare the frequencies of genes associated with virulence within groups separated by function. Significance was defined as p < 0.05.
Results
Antimicrobial susceptibility test
The antimicrobial susceptibility test results are pre-sented in Figure 1. Of the 148 analyzed strains, only 25 (16.9%) were susceptible to all antimicrobials tested, regardless of the isolation source. CIP and SOX presented a significantly higher number of nonsusceptible strains (p < 0.05). A total of 14 (9.5%) strains were classified as multidrug-resistant Salmonella Enteritidis. Most of these strains (10/14) were isolated from avian sources, although the difference between isolation sources was not significant (p > 0.05).
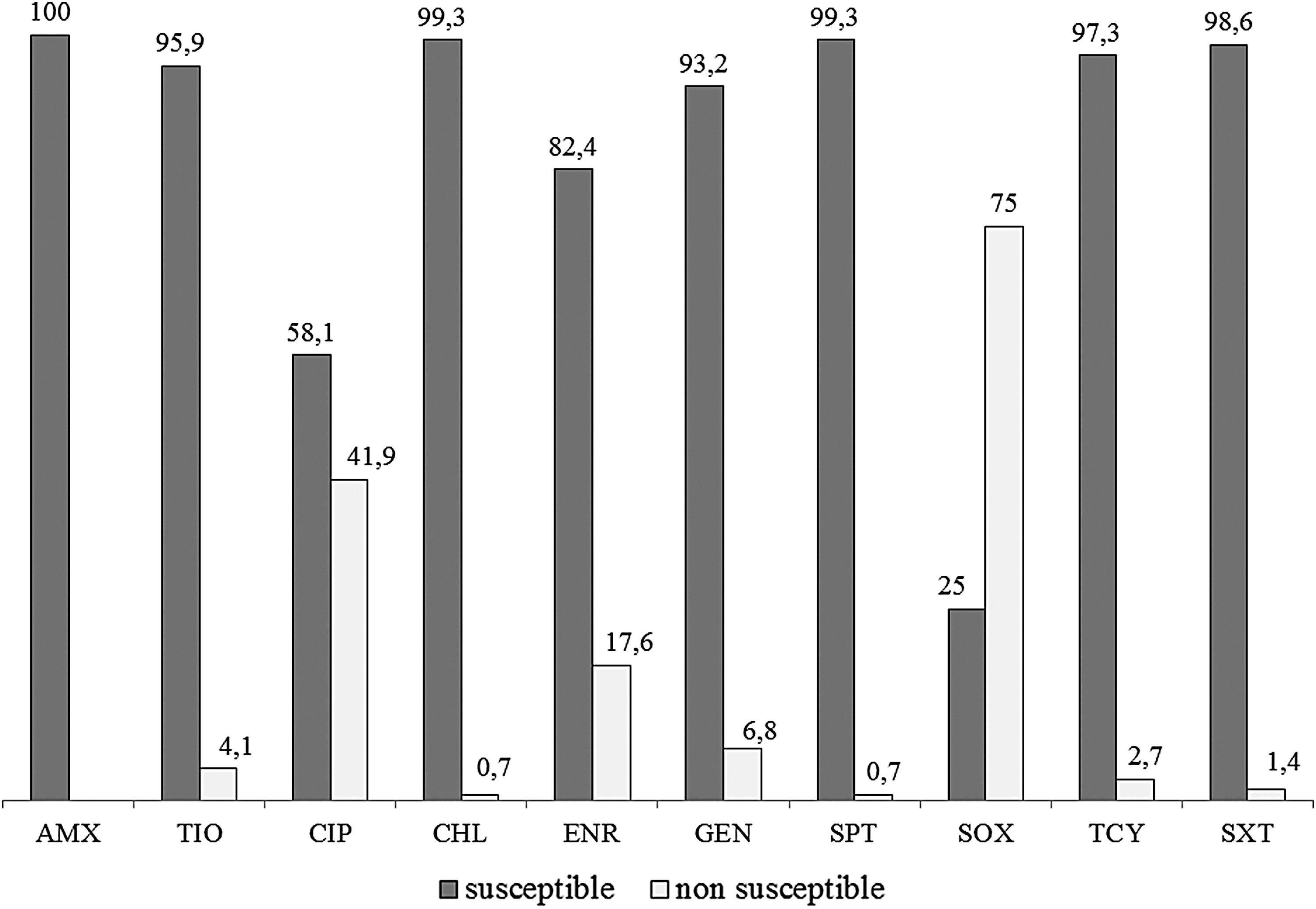
Antimicrobial susceptibility (%) of Salmonella Enteritidis SE86 to 10 antimicrobial agents. AMX, amoxicillin; TIO, ceftiofur; CIP, ciprofloxacin; CHL, chloramphenicol; ENR, enrofloxacin; GEN, gentamicin; SPT, spectinomycin; SOX, sulfafurazole; SXT, sulfamethoxazole with trimethoprim; TCY, tetracycline.
The antimicrobial resistance to the tested antimicrobials according to the isolation source is presented in Table 4.
Bold data indicate a positive association (p < 0.05) between antimicrobial and source of isolation.
AMX, amoxicillin; TIO, ceftiofur; CIP, ciprofloxacin; CHL, chloramphenicol; ENR, enrofloxacin; GEN, gentamicin; SPT, spectinomycin; SOX, sulfafurazole; SXT, sulfamethoxazole with trimethoprim; TCY, tetracycline.
Biofilm formation assay
Of the 148 analyzed strains, 136 (91.9%) produced biofilms at a minimum of one of the tested temperatures, although to various extents (Fig. 2). A total of 107 (72.3%) strains produced biofilms at 37°C, 98 (66.2%) at 28°C, 87 (58.8%) at 12°C, and 52 (35.1%) at 3°C independent of the isolation source.
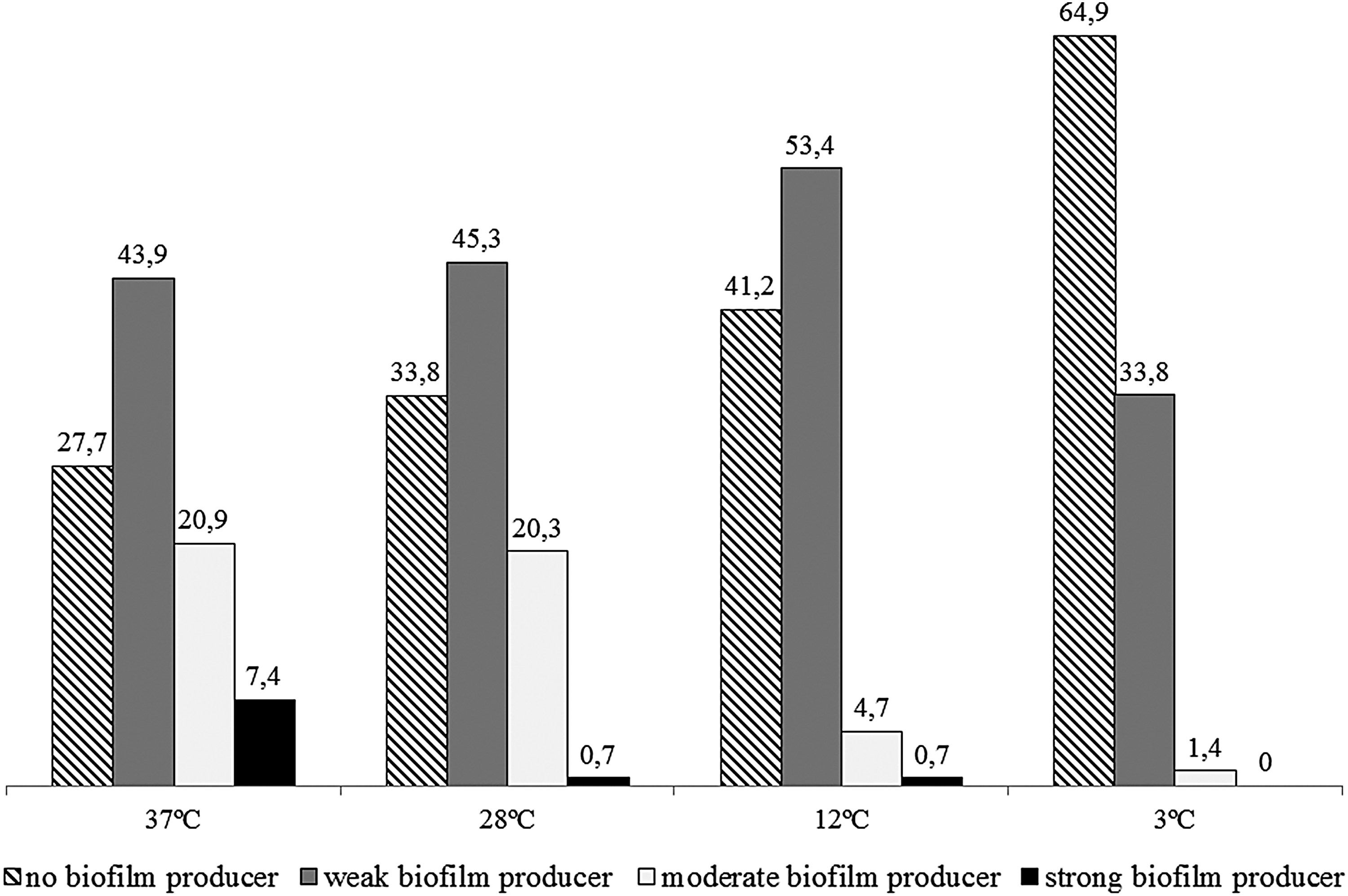
Classification of Salmonella Enteritidis SE86 according to biofilm production at 37°C, 28°C, 12°C, and 3°C (%).
The results obtained for the strains with different isolation sources according to the incubation temperature are shown in Table 5. Strains from both origins produced more biofilms at 37°C. A total of 77 of the 80 strains isolated from salmonellosis outbreaks (96.3%) produced biofilms, whereas 59 (86.8%) of the strains isolated from avian sources produced these structures at a minimum of one of the tested temperatures. However, biofilm production was independent of the isolation sources of the analyzed samples (p > 0.05) for all temperatures.
NP, no biofilm producer; WP, weak biofilm producer; MP, moderate biofilm producer; SP, strong biofilm producer.
Detection of virulence-associated genes
The frequencies for the 27 genes according to the isolation source are described in Table 6. Most of the investigated genes presented a high frequency and a regular distribution regardless of the origin.
Bold data indicate significant association (p < 0.05) between gene and source of isolation.
TTSS, type three secretion system.
Determination of PT4 by PCR
Although all Salmonella Enteritidis SE86 strains belong to a unique clonal group, only 142 (95.9%) of the strains were classified as PT4 by PCR. Two strains isolated from food products involved in salmonellosis outbreaks and four strains from poultry sources belonged to PTs other than PT4.
Discussion
High resistance rates to sulfonamides and tetracycline are common in farm animals and humans and are possibly related to the extensive use of these agents (WHO, 2011a, 2011b; EFSA and ECDC, 2015; Ziech et al., 2016; Voss-Rech et al., 2017). In Brazil, the therapeutic use of these substances for veterinary use is not controlled. However, these drugs are banned for use as a feed additive (Brasil, 2009).
A total of 38.5% of the strains were resistant to ENR and CIP simultaneously. Fluoroquinolones are the main antimicrobial agent class for the treatment of salmonellosis in humans (WHO, 2011a; EFSA and ECDC, 2015). Although ENR is used only in animals, microorganisms that are not susceptible to this drug can become resistant to other structurally related substances, such as CIP (Goncagül et al., 2004; Marshall and Levy, 2011; WHO, 2011a, 2011b). In Australia and Finland, where their use has never been allowed in animal production, the rate of CIP-resistant bacteria in humans is very low, particularly compared with the rates in countries where their use is allowed or has been allowed for several years, such as the United States (Collignon, 2005; Cheng et al., 2012).
Because the use of fluoroquinolones is not recommended for children due to their potential adverse effects (EFSA and ECDC, 2015), third-generation cephalosporins are the first choice for the treatment of salmonellosis in these patients (WHO, 2011a; EFSA and ECDC, 2015). Strains that are resistant to TIO may present cross-resistance to ceftriaxone, which is widely used in humans. In this study, only six (4.1%) strains were nonsusceptible to TIO, all of which were isolated from avian sources.
According to the WHO, antimicrobial resistance in humans and animals is the major challenge in public health in the 21st century (WHO, 2011a), and the relationship between resistant bacteria in humans and the use of antibiotics in food animals has been continuously studied. In our study, only 4 (5.9%) strains isolated from poultry were susceptible to all antimicrobial agents, whereas 21 (14.2%) of the strains isolated from outbreaks did not present resistance to any antibiotics. Nineteen antimicrobial resistance profiles were obtained, but only five were common among poultry and outbreak strains: (1) SOX; (2) SOX, CIP; (3) SOX, CIP, ENR; (4) SOX, SXT; and (5) SOX, CIP, ENR, GEN. Profiles (1), (2), and (3) were the most common for both origins. Some authors also found greater resistance in poultry strains than in strains isolated from humans (Oliveira et al., 2005; Cailhol et al., 2006; Vaz et al., 2010). According to Foley and Lynne (2008), strains isolated from different species of farm animals usually present higher resistance than human strains. However, other studies have found different results and demonstrated a higher antimicrobial resistance among human strains (Al-Dawodi et al., 2012; Elgroud et al., 2015).
Our data indicate that resistance to TIO, gentamicin, and SOX is associated (p < 0.05) with strains of avian origin. The exclusive use of TIO in veterinary medicine and the wide use of sulfonamides in poultry production may explain this association. Gentamicin is usually used in Brazilian hatcheries for the treatment of enterobacteria, which may lead to the spread of strains resistant to this antibiotic in the field (Almeida and Palermo-Neto, 2005).
The Salmonella Enteritidis serotype has been described as the strongest biofilm producer (Schonewille et al., 2012; Puffal, 2013). The frequent involvement of Salmonella Enteritidis in salmonellosis outbreaks may be a consequence of the strong capacity of some strains to produce biofilms (Ivana et al., 2015). Salmonella Enteritidis SE86 was previously shown to have higher adhesion capabilities on several surfaces, including different types of stainless steel (AISI 304 or 316), polyethylene, and metal inert gas and tungsten inert gas melts (Casarin et al., 2014, 2016; Tondo et al., 2015). SE86 also demonstrated acid adaptation after sublethal pH exposure and became more resistant to both acid and thermal exposure (Malheiros et al., 2009; Tondo et al., 2015).
Previous studies demonstrated a strong influence of temperature on biofilm production, particularly for some serotypes, including Salmonella Enteritidis (Borges et al., in press). However, in this study, only the avian strains presented a significant difference (p < 0.05) between the four analyzed temperatures. For the outbreak strains, no significant difference (p > 0.05) in biofilm production was found between the different temperatures.
Biofilm production was independent of the sources of isolation at any temperature. However, other authors found significant differences in biofilm production by several microorganisms, including Salmonella, according to the origin of isolation (Trappetti et al., 2013; Almohamad et al., 2014; Doijad et al., 2015; Piras et al., 2015; Lamas et al., 2016).
Our biofilm formation assay demonstrated that strains within the same clonal group isolated from different sources showed variation in biofilm production behavior. These data reinforce the importance of studying these complex structures to better understand the influence of the genetic apparatus involved in the mechanisms underlying biofilm formation.
The frequencies of the screened virulence-associated genes were similar to previous reports (Elemfareji and Thong, 2013; Mezal et al., 2013, 2014; Abdallah et al., 2014; Krawiec et al., 2015). spvB, spiA, pagC, sipB, prgH, spaN, sitC, and lpfC genes were positively associated (p < 0.05) with the avian strains, whereas iroN showed a positive association (p < 0.05) with the strains isolated from salmonellosis outbreaks. However, previous studies did not find a link between the isolation source of a strain and its virulence gene repertoire (Oliveira et al., 2003; Salisbury et al., 2011; Mezal et al., 2013).
Regardless of the isolation source, the comparisons of genes within functional groups showed significant differences (p < 0.05) in the structural proteins of the type three secretion system and iron metabolism genes. orgA gene was the most frequent, possibly because this gene is essential for cellular invasion by Salmonella (Klein et al., 2000). Similarly, sitC presented a higher frequency than iroN. The absence of these genes is unlikely to hinder the survival and multiplication of Salmonella strains due to the presence of several alternative mechanisms (Janakiraman and Slauch, 2000).
Since the introduction of the PT4 in Brazil in 1993, several studies have shown that this PT remains the most identified type in southern Brazil (Ribeiro et al., 2007; Vaz et al., 2010). Our results are similar to the results of previous studies and official data (Pang et al., 2007; Ribeiro et al., 2007; Brasil, 2008, 2012; Kalender et al., 2009; Vaz et al., 2010). According to Mare et al. (2001), strains with high genetic relatedness can be classified as different PTs because they are not always genetically related. However, there are reports of PT conversion after the acquisition or the loss of plasmids (Brown et al., 1999). It has been described that PT4 can be converted to PT7, PT8, or PT24 after plasmid acquisition or mutations (Rankin and Platt, 1995; Brown et al., 1999). This could explain that not all SE86 strains were classified as PT4 in our study.
Finally, we did not confirm the previously described association between biofilm production and resistance to antimicrobials (Wang et al., 2013; Ghasemmahdi et al., 2015; Piras et al., 2015). However, we observed that 92.9% (13/14) of the multidrug-resistant strains were able to produce biofilms. No relationship was found between antimicrobial resistance, biofilm production, and the presence of virulence-associated genes.
Conclusions
Our results indicated that poultry strains presented higher resistance and a greater number of multidrug-resistant strains than strains isolated from food involved in salmonellosis outbreaks. Biofilm production was independent of the isolation source at any temperature. Most of the investigated genes presented a high frequency and a regular distribution regardless of the origin. The majority of the strains were classified as PT4. No relationship was found among biofilm production, antimicrobial resistance, and virulence profile.
Footnotes
Acknowledgment
This work was supported by the Brazilian National Council of Technological and Scientific Development—CNPq [Grant No. 476092/2013-2].
Disclosure Statement
No competing financial interests exist.