
Abstract
Arcobacter butzleri, Arcobacter cryaerophilus, and Arcobacter skirrowii are Gram-negative pathogenic microorganisms that cause watery diarrhea and septicemia in humans. The aims of this study were to detect the presence of Arcobacter spp. in chicken meat from butcher shops in São Paulo (Brazil) and to verify their virulence genes and genotypic profiles. A total of 300 chicken cuts were analyzed. The results show the presence of Arcobacter spp. in 18.3% of samples, which were identified as A. butzleri (63.6%) and A. cryaerophilus (36.3%). All strains were positive for the virulence genes ciaB and mviN, followed by cj1349 (98%), pldA (94.4%), cadF (72.7%), tlyA (92.7%), hecA (49%), irgA (47.2%), and hecB (34.5%). These strains were subjected to single-enzyme amplified fragment length polymorphism. Nineteen genotypic profiles were obtained for A. butzleri, and 17 for A. cryaerophilus. These results confirm the presence of virulent strains of A. butzleri and A. cryaerophilus in the chicken meat in Brazil. The presence of potentially virulent strains of Arcobacter highlights a possible public health risk, particularly with respect to ingestion of undercooked foods and cross-contamination from uncooked foods during food preparation and contaminated utensils.
Introduction
A
Poultry and pigs serve as reservoirs of this agent, increasing the risk of infection associated with raw or undercooked meat from these sources (Ho et al., 2008). On the farms, infection of animals with Arcobacter spp. is generally unapparent and difficult to diagnose (De Smet et al., 2011). The clinical signs of human illness include bloody diarrhea, fever, and abdominal pain. Some patients may experience vomiting (Ferreira et al., 2014). An experimental study using strains of Arcobacter spp. in human colonic epithelial cells revealed the ability of these bacteria to produce gut barrier dysfunction leading to watery and potentially leaky diarrhea (Collado and Figueras, 2011).
Currently, the mechanisms of pathogenicity of Arcobacter and the virulence properties of strains are poorly understood. The potential limitations of methods for culture and isolation of the studied agent has led to the increased use of molecular approaches for detection and species identification, analysis of virulence genes, and epidemiological studies (Gonzalez et al., 2012). The pathogenesis of the agent has been studied by comparison with Campylobacter spp., including characteristics such as invasiveness, accession by fibronectin, and presence of hemolysin proteins (Ho et al., 2007). In addition, there is a gene regulator described on the outer membrane iron uptake system (irgA) in Arcobacter, which was previously reported in Vibrio cholerae and uropathogenic Escherichia coli (Douidah et al., 2012). Khoshbakht et al. (2014) described the detection of A. butzleri, A. cryaerophilus, and A. skirrowii in foods of animal origin and these isolates were found to contain six known virulence factors (including cadF, ciaB, cj1349, mviN, pldA, and tlyA).
The aim of this study was to isolate strains of Arcobacter spp. from cuts of chicken meat sold in São Paulo (Brazil) and to identify the pathogenic species, virulence factors, and circulating genotypes.
Materials and Methods
Poultry meat samples and bacterial isolation
Three hundred cuts of chicken meat (refrigerated raw skin on breast, wing, thigh, and drumstick) from 77 markets and butchers were analyzed. The samples were collected from July 2013 to July 2016, in five geographic regions in São Paulo, Brazil (North, South, East, West, and Center). The stores and samples were selected by convenience-based criteria. The samples were transported to the laboratory under refrigeration and were processed the same day. A total of 25 g of meat was put into sterile bags containing 225 mL of buffered peptone water. The samples were homogenized in Stomacher Bags and agitated for an average time of 1 min. An aliquot of 1 mL of peptone water was added to 9 mL of selective enrichment broth, as described by Johnson and Murano (1999), and the test tubes were homogenized for 15 s. The tubes were incubated in an aerobic environment for 48 h at 30°C. After the incubation period, a volume of 10 μL of broth was deposited on a sterile membrane (0.45 μm) and placed on the surface of JM selective agar (Johnson and Murano, 1999). The filters were removed after 1 h and the agar plates were streaked and incubated in an aerobic environment for 48 to 72 h at 30°C.
Small colonies with features of Arcobacter spp. were selected, submitted for Gram stain, subjected to polymerase chain reaction (PCR), and stored at −80°C. The reference strains, A. butzleri ATCC 49616, A. cryaerophilus ATCC 43158, and A. skirrowii ATCC 51132, were used as PCR control.
Identification and determination of virulence genes by PCR
The bacterial DNA was purified as described by Boom et al. (1990). The PCR for species identification was performed with primers described by Douidah et al. (2010), using 5 μL bacterial DNA (10 ng/μL), 1.5 mM MgCl2, 10 pmol of each primer, 1.0 U Taq DNA polymerase, 1 × PCR buffer, and water in a final volume 50 μL. The reaction was carried out with an initial denaturation at 94°C for 4 min followed by 35 cycles of 1 min at 94°C, 1 min at 55°C, and 1 min at 72°C.
The PCR method was also used to determine the presence of the putative virulence genes as described by Douidah et al. (2012). These genes codify for the following: adhesion (cadF and cj1349), invasion (ciaB), phospholipase (pldA), hemolysin (tlyA), regulation of an outer membrane protein associated with iron uptake (irgA), hemagglutinin (hecA), as well as genes associated with hemolysin activation (hecB) and a virulence marker (mviN). The amplification mixture consisted of 10 mM Tris-HCl buffer (pH 8.3), MgCl2, 200 mM deoxynucleoside triphosphates, DNA primers, 1 U Taq DNA polymerase, and ultrapure water at a final volume 50 μL. The amplified products were separated by electrophoresis in 1.5% agarose gel stained with BlueGreen® (LGC Biotecnologia, São Paulo, Brazil), and identified through a 100 bp DNA ladder (New England BioLabs, Inc., Ipswich, MA).
Single-enzyme amplified fragment length polymorphism
The Arcobacter strains were genotyped using single-enzyme amplified fragment length polymorphism (SE-AFLP). Restriction endonuclease digestion and ligation were performed as described by McLauchin et al. (2000). To 10 μL DNA, 24 U Hind III (New England BioLabs, Inc.) and ultrapure water were added, resulting in a final volume of 20 μL. After overnight incubation at 37°C, 5 μL digested DNA was added to 0.2 μg of ADH1 and ADH2 oligonucleotides, and 1 U T4 DNA ligase and ultrapure water, making 20 μL of final volume. The mixture was incubated for 3 h at room temperature and heated to 80°C for 10 min. PCR was performed using 5 μL ligated DNA, 2.5 mM MgCl2, 30 pmol primer (HI-G), 1 U Taq polymerase, and 1 × PCR buffer (final volume—50 μL). This mixture was denatured at 94°C for 4 min, 35 cycles of 1 min at 94°C, 1 min at 60°C, and 2.5 min at 72°C. Electrophoresis was performed using a 2% agarose gel at 24 V for 26 h. The amplified products were stained with GelRed™ (New England BioLabs, Inc.) and compared with a 100-bp DNA ladder (New England BioLabs, Inc.).
For analysis of the generated fragments, Bionumerics 7.6 (Applied Maths, Sint-Martens-Latem, Belgium) was used. The similarity of the isolates was estimated through the Dice coefficient. With the generated similarity matrix, it was possible to determine the genotypic groups using unweighted pair group method using arithmetic average (UPGMA). We applied a cutoff of 90% genetic similarity to determine the genotypic profiles. The discriminatory index was calculated according to Hunter and Gaston (1988).
Statistical analysis
The Fisher's exact test was used for comparisons between presence of Arcobacter in cuts of chicken meat and virulence scores. The software used was BioStat® 5.3.5 (AnalystSoft) (considered significant p-value ≤0.05).
Results
A total of 55/300 samples (18.3%) from 33/77 markets were positive for Arcobacter spp. One strain was evaluated from each meat sample—35/55 (63.6%) were identified as A. butzleri and 20/55 (36.3%) as A. cryaerophilus. The distribution of isolates and respective identified species are presented in Table 1. There were no statistical differences between the cuts of chicken meat analyzed.
The virulence genes ciaB and mviN were detected in 100% (55/55) of the strains, 98.1% (54/55) also had cj1349, 94.4% (52/55) had pldA, 72.7% (40/55) had cadF, 92.7% (51/55) had tlyA, 49% (27/55) had hecA, 47.2% (26/55) had irgA, and 34.5% (19/55) had hecB. The distribution of virulence genes detected in A. butzleri and A. cryaerophilus strains is described in Table 2. There were no statistical differences between the distribution of virulence genes in A. butzleri and A cryaerophilus. The 55 Arcobacter spp. strains were grouped into 21 virulence profiles (G1 to G21) (Table 3).
G*-profile of virulence.
For SE-AFLP analysis, it was decided to evaluate the A. butzleri and A. cryaerophilus species separately. The two dendrograms demonstrate high heterogeneity: The samples collected in the same year were arranged in completely different SE-AFLP profiles. The discriminatory index for A. butzleri was 0.96, which exhibited 19 different SE-AFLP profiles (Fig. 1), and that of A. cryaerophilus was 0.98, with 17 SE-AFLP profiles (Fig. 2).
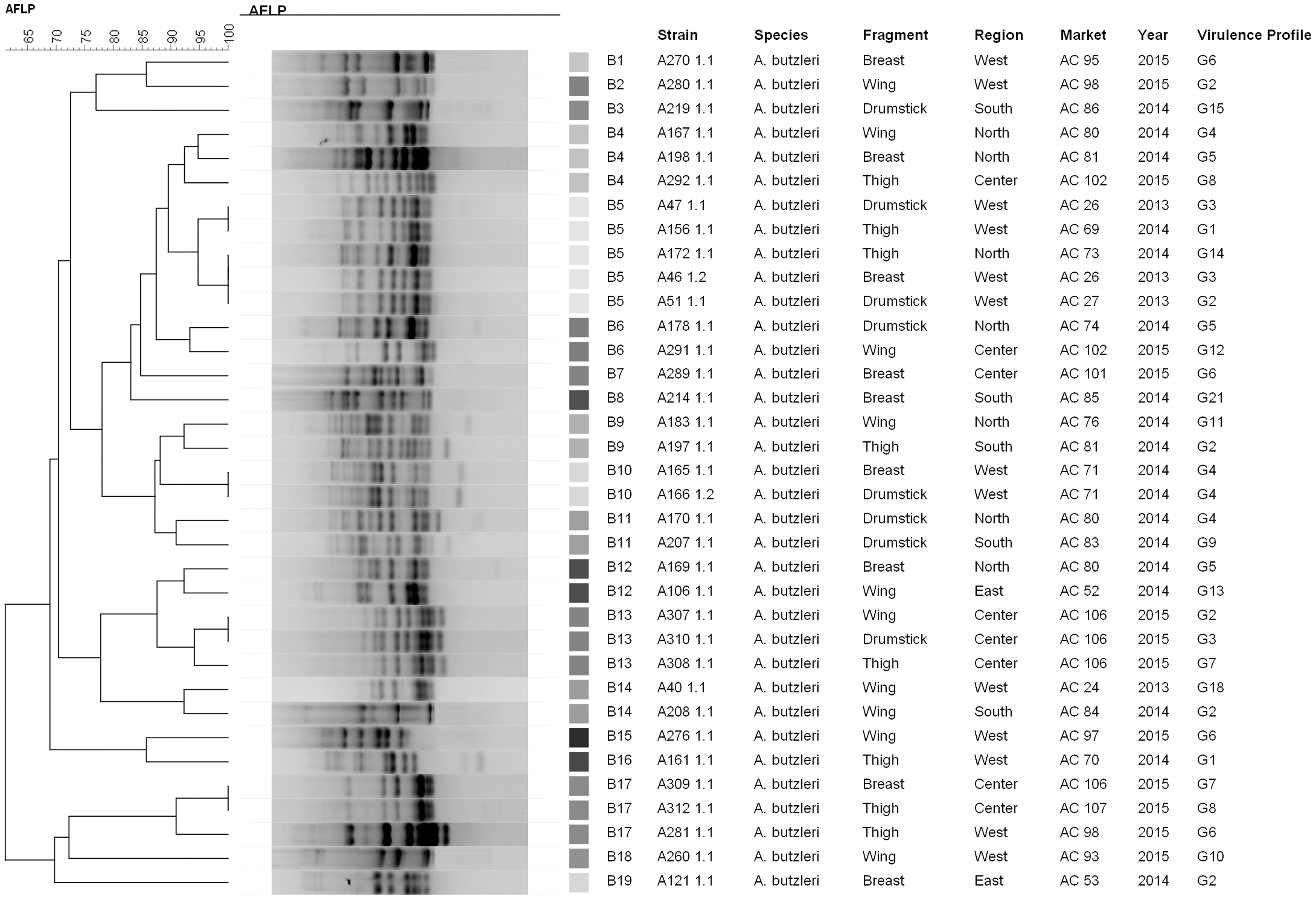
Genotypic profiles of Arcobacter butzleri detected in samples of chicken meat from butcher shops of the city of São Paulo. AFLP, amplified fragment length polymorphism.

Genotypic profiles of Arcobacter cryaerophilus detected in samples of chicken meat from butcher shops of the city of São Paulo.
Discussion
Food safety and disease prevention are among the goals of all governments worldwide. However, there are many limitations to traditional inspection, sampling, and analysis of animal products (ICMSF, 2006). Inspection sampling is often risk based and seeks a narrow range of pathogens; therefore, may not represent consumer exposure, ignoring potentially emerging pathogens, such as Arcobacter spp. This study analyzed 300 cuts of chicken meat and 55 (18.3%) samples were positive for Arcobacter spp. In a similar study, 37/80 (46.25%) chicken carcasses from a Brazilian slaughterhouse were contaminated with Arcobacter spp.; of these 85% of isolates were A. butzleri (Oliveira et al., 2001). The values obtained in our retail study were substantially lower than those observed in the slaughterhouse study. This difference may be related to the locale and year of collection, or, more likely, to methodologies, but still suggests that chicken carcass contamination can occur in slaughterhouses and extend to chicken meat at retail.
Khoshbakht et al. (2014) evaluated the presence of different species of Arcobacter in broiler carcasses, equipment, and facilities of Iranian slaughterhouses. The results showed A. butzleri in 73% of the 540 strains, followed by A. cryaerophilus (9%), and A. skirrowii (4.1%). These data agreed with the results obtained in our research, with the predominance of the A. butzleri species in 35/55 isolates, followed by A. cryaerophilus with 20/55. The two species were identified in all the cuts examined (wing, thigh, drumstick, and breast) (Table 1). A. butzleri has been isolated in animal reservoirs such as birds (chickens and turkeys) and mammals, for example swines, bovines, dogs, and cats that are alive, dead, healthy, and sick (Collado and Figueras, 2011).
A. butzleri species (65.4%) and A. cryaerophilus (34.5%) found in this study were analyzed to identify their virulence genes: cadF (fibronectin protein), ciaB (invasion), cj1349 (fibronectin protein), pldA (lysis of erythrocytes), mviN (peptidoglycan biosynthesis), tlyA (hemolysin), hecB (encoding hemolysin protein activation), hecA (hemagglutinin filaments which involves attack, aggregation, and cell death), and irgA (regulator of protein for iron acquisition) (Douidah et al., 2012; Karadas et al., 2013). The nine putative genes researched have received special attention, due to their homology to genes associated to pathogenicity in other microorganisms (Ferreira et al., 2016). The virulence factors studied are the homologues those of Campylobacter spp. and other bacteria such as E. coli and Vibrio spp. Only the species A. butzleri has a number of well-established virulence genes (Douidah et al., 2012). In our work, it was possible to identify the coexistence of all nine genes in 4/55 strains of Arcobacter obtained from chicken meat (wing, breast, and drumstick) from different locations (North and West regions of São Paulo).
In previous research on the genetic diversity of A. butzleri and A. cryaerophilus in beef, pork, and chicken, it was reported that 100% of the strains of A. butzleri presented the tlyA gene, and 100% of the strains of A. cryaerophilus presented the ciaB gene (Zacharow et al., 2015). The strains of A. cryaerophilus isolated from chicken meat did not present any other virulence factor, whereas those of A. butzleri chicken isolates also showed that 90% contained cadF, 71% had cj1349, 100% encoded ciaB, 53% contained irgA, 34% had hecA, 48% encoded hecB, 93% contained pldA, and 98% had mviN (Zacharow et al., 2015). Although there is a divergence between the virulence profile of A. cryaerophilus reported by Zacharow et al. (2015) and those obtained in the present study, the genetic diversity reported by the authors was also observed in this study.
In our study, the groups called G1 and G2 were those that had a higher number of isolates (n = 7), with a combination of eight and six genes, respectively; and the G4 group had the four isolates that contained all nine virulence factors. The other virulence profiles presented a varied combination of genes; however, the G2 virulence profile presents the combination of six important genes (cadF, ciaB, cj1349, mviN, pldA, and tlyA), previously reported by Douidah et al. (2012) from pathogenic strains of A. butzleri.
Douidah et al. (2014) suggests that the high genome heterogeneity of the Arcobacter genus results in pathogenicity differences between species. In our study, the profiles demonstrate greater heterogeneity in A. cryaerophilus when compared with A. butzleri. Some isolates of A. cryaerophilus came from the same butcher. The samples collected in the same year were arranged in completely different SE-AFLP profiles, as were those isolated from the butchers AC 74 and AC 75, which, despite having a collection site in common, were distributed among six different SE-AFLP profiles (C1, C3, C7, C11, C12, and C14), featuring completely different virulence profiles.
A. butzleri profiles also show heterogeneity, without a common grouping between isolates from the same collection. The SE-AFLP profile named B5 grouped isolates with high similarity, despite samples being collected in different years. They came from butchers and distinct regions, and their virulence profiles varied. However, the SE-AFLP profile B13 contained grouped isolates with high similarity, collected at the same establishment and in the same year, but with variability in their virulence profiles. In contrast, profile B10 is a homogeneous group composed of two isolates that have high similarity, with the collected samples in the same place and year, with the same virulence profile.
The SE-AFLP method has been shown to be sensitive in epidemiological investigations, as is shown in the survey of A. butzleri in chickens slaughtered in Nigeria, whose strains were grouped into 12 genotypic profiles, with 90% similarity (Amisu et al., 2003). A Danish study (On et al., 2004) using SE-AFLP of 73 strains of A. butzleri from different geographical locations had similar results compared with those reported in this study, showing great genetic diversity and multiple genotypes (51 among 73 strains).
Chicken meat is considered the main route of transmission of Arcobacter and poor management practices and hygiene failures in the handling of food may increase the risk of contamination by potentially disease-causing strains of Arcobacter (Fernandez et al., 2015). In conclusion, our study showed the presence of virulent strains of A. butzleri and A. cryaerophilus in chicken meat sold in butcher shops and municipal markets in São Paulo, Brazil. The strains had a great diversity of genotypes and the same virulence traits described for Arcobacter spp. isolated from human with gastroenteric disease. Preventive actions should be taken to reduce this emerging foodborne illness, including biosecurity-based strategies on poultry farms, systematic analysis of the critical control points in slaughterhouses, butchery hygiene practices, and consumers' orientation. Further studies on the pathogenicity of these strains, using animal models, will be useful to understanding the risks associated with the ingestion of Arcobacter-contaminated chicken meat.
Footnotes
Acknowledgments
This study was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (Grant 14/07837-6). OLIVEIRA, M.G.X. is recipient of postgraduate fellowship from FAPESP (Grant 14/06584-7).
Disclosure Statement
No competing financial interests exist.