
Abstract
Ruminants are important reservoirs of E. coli O157:H7 and are considered as the major source of most foodborne outbreaks (e.g., 2017 outbreak in Germany, 2014 and 2016 outbreaks in United States, all linked to beef products). A promising strategy to reduce E. coli O157 is using antimicrobials to reduce the pathogen levels and/or virulence within the animal gastrointestinal tract and thus foodborne disease. The aim of the study was to determine the efficacy of a commercial mixture of natural antimicrobials against E. coli O157. The minimum inhibitory concentration and minimum bactericidal concentration of the antimicrobial were quantitatively determined and found to be 0.5% and 0.75% (v/v) of the natural antimicrobial, respectively. Microbial growth kinetics was also used to determine the effect of the antimicrobial on the pathogen. The natural antimicrobial affected the cell membrane of E. coli O157, as demonstrated by the increase in relative electric conductivity and increase in protein and nucleic acid release. The antimicrobial was also able to significantly reduce the concentration on E. coli O157 in a model rumen system. Biofilm assays showed that subinhibitory concentrations of the antimicrobial significantly reduced the E. coli 0157 biofilm forming capacity without influencing pathogen growth. In addition, the natural antimicrobial was able to reduce motility and exopolysaccharide production. Subinhibitory concentrations of the antimicrobial had no effect on AI-2 production. These findings suggest that the natural antimicrobial exerts an antimicrobial effect against E. coli O157 in vitro and in a model rumen system and could be potentially used to control this pathogen in the animal gut. The results also indicate that subinhibitory concentrations of the antimicrobial effectively reduce biofilm formation, motility, and exopolysaccharide production.
Introduction
E
There are no reliable commercially available strategies to control the carriage of E. coli O157 in ruminants (Rivas et al., 2010).
The objectives of this pilot study were to determine the antimicrobial effect of a commercial mixture of natural antimicrobials against different strains of E. coli O157:H7 and also to investigate its antimicrobial mechanism and also to determine its effect on motility, biofilm formation, exopolysaccharide production, and QS at subinhibitory concentrations.
Materials and Methods
Bacterial strains and antimicrobial
E. coli O157:H7 EDL 933 (ATCC 43895), E. coli O157 VSD 400, and E. coli O157 PHL 09 were maintained in Tryptone Soya Agar plus 0.6% yeast extract (TSAYE, Oxoid, United Kingdom) slopes at 4°C. When necessary the pathogens were cultured in Mueller-Hinton broth (MHB) at 37°C for 24 h. The antimicrobial product Auranta 3009 was obtained from Envirotech Innovative Products Ltd and consisted of lactic acid, E330 citric acid, and citrus extract. The manufacturer of this product is Bio Science Nutrition Ireland Ltd, Ireland, DAFM No.IELK241361.
Growth curves
The three individual E. coli O157 culture (6.0 log CFU/mL in MHB) was transferred into 96 well microtiter plates with final concentrations of the natural antimicrobial of 0%, 0.0625%, 0.125%, 0.25%, 0.5%, and 0.75% (v/v). The inoculum level was determined after plating out onto Mueller-Hinton Agar (MHA) plates and incubation at 37°C for 24 h. Cultures were incubated at 37°C, and optical density was monitored at 600 nm at intervals of 1 h, over 24 h, using an automatic plate reader (FLUOstar Omega; BMG LABTECH). The concentrations of the antimicrobial that did not inhibit bacterial growth were chosen as the subinhibitory concentrations and used for the subsequent phenotypic virulence assays.
Minimum inhibitory concentration and minimum bactericidal concentrations
To determine minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) we have used a methodology as previously described by Zhu et al. (2016). Briefly, the antimicrobial was diluted from 8% to 0.015% (v/v) in MHB. An overnight bacterial culture was diluted to approximately 6 Log CFU/mL in MHB. A tube containing each dilution of the natural antimicrobial in MHB was inoculated with 5.49 ± 0.16 log CFU/mL of bacterial culture. Noninoculated tubes were used as positive controls. Subsequently, the tubes were incubated at 37°C for 24 h. The highest dilution of the natural antimicrobial in the tube without visible bacterial growth was considered as the MIC. To determine the MBC, 100 μL of the cultures that showed no growth were spread plated onto MHA plates and incubated at 37°C for 24 h.
Cell membrane permeability
E. coli O157:H7 cell membrane permeability was determined by assessing the electric conductivity of cultures treated with different concentrations of the natural antimicrobial for 2, 6, and 24 h, as described by Stratakos et al. (2018). A conductivity meter (Jenway 4200) was used to quantify changes in electric conductivity. Bacteria were washed with 5% glucose until their electric conductivity was near to that of 5% glucose (isotonic bacteria). The antimicrobial was added to 5% glucose (0, MIC, MBC), and the electric conductivities were measured (L 1). The same concentrations of the antimicrobial were also prepared with the isotonic bacteria, and the conductivities were measured after 2, 6, and 24 h incubation at 37°C (L2). The conductivity of bacterial cells in 5% glucose treated in boiling water for 5 min was used (L0) as control. The permeability of cell membrane was calculated using the equation: Relative electric conductivity (%) = 100 (L2–L1)/L0.
Membrane integrity
Membrane integrity for the three E. coli strains was evaluated in terms of protein leakage and 260 nm absorbing materials into the cell suspension as previously described (Stratakos et al., 2018), after a 6 h treatment with different concentrations of natural antimicrobial (0, MIC and MBC). The protein content in the supernatants was assessed using the Pierce BCA Protein Kit (Thermo Scientific). Nucleic acid leakage in terms of optical density of 260 nm absorbing materials was performed using a UV-vis spectrophotometer (FLUOstar Omega; BMG LABTECH). Untreated samples were used as controls.
In vitro rumen assay
The antimicrobial efficiency of the natural antimicrobial (0, MIC, MBC, and 2 × MBC) against E. coli O157 EDL 933 was tested in the assay. Three dairy cows (Swedish-Red × Jersey/Holstein-Friesian) fitted with rumen cannula were used as rumen fluid donors. The rumen fluid was collected 3 h after the morning feeding. At the laboratory, the rumen contents (solid and liquid phases) were homogenized, filtered through four layers of cheesecloth, and maintained at 39°C under anaerobic conditions. The filtered rumen fluid was dispensed into bottles and diluted with four parts of a buffer solution. The buffer solution was prepared by combining two solutions: buffer A and buffer B (5:1). The following quantities were used to prepare one liter of buffer A: 10 g KH2PO4; 0.5 g MgSO4.7H2O; 0.5 g NaCl; 0.1 g CaCl2.2H2O; and 0.5 g Urea. One liter of buffer B was prepared using the following: 15 g Na2CO3 and 1 g Na2 S.9H2O. After mixing at 39°C, the combined buffer solution was adjusted to a final pH of 6.8. An overnight culture of E. coli O157 was prepared and serially diluted in 9 mL of PBS and inoculated into the rumen at approximately 6 Log CFU/mL. All bottles were flushed with CO2 for approximately 1 min before incubation at 39°C, with agitation. Samples were taken from each bottle at 0, 4, 18, and 24 h. After sampling at each time point, all bottles were flushed with CO2 for approximately 1 min before reincubation at 39°C. For spread plating, 1 mL aliquots taken from each bottle were serially diluted in 9 mL volumes of maximum recovery diluent (MRD), and 0.1 mL aliquots of each dilution were spread onto plates of TSA which were incubated at 37°C for 2 h. Subsequently, they were over poured with MacConkey Agar with Sorbitol, Cefixime, and Tellurite (CT-SMAC) and incubated for another 48 h to allow for the recovery of injured cells. The suspected E. coli O157 positive colonies were confirmed by latex agglutination (Prolex™ E. coli O157 Latex Test Reagent Kit; Pro-Lab Diagnostics). Treatments and control samples (without antimicrobial addition) were incubated in triplicate. The assay was performed in duplicate, using fresh rumen fluid on 2 separate days.
Biofilm formation and motility assay
A biofilm formation assay for E. coli EDL 933 was performed in 96-well polystyrene plates according to the method described by Lee et al. (2014). Briefly, overnight cultures were inoculated into Luria-Bertani (LB) broth (Oxoid) with known concentrations (0%, 0.0625%, and 0.125%) of the antimicrobial and cultured for 24 h without shaking at 37°C. To quantify biofilm formation, wells were washed thrice with PBS to remove all nonadherent cells. Cells were fixed with 200 μL of methanol for 20 min. Biofilms were stained with crystal violet (0.4% w.v) for 15 min and washed thrice with PBS. The formed biofilm was then dissolved with 200 μL 96% ethanol for 30 min, and absorbance was measured (570 nm) in a microtiter plate reader. Cultures with no added antimicrobial served as positive controls. Motility was determined as described by Kulshreshtha et al. (2016). Briefly 5 μL of E. coli O157 culture (∼7.0 log CFU) was stabbed into plates with semisolid medium (MHB supplemented with 0.5% agar) with and without 0.0625% and 0.125% of the natural antimicrobial. Plates with no added antimicrobial served as positive controls.
Exopolysaccharide content determination
An E. coli O157 EDL 933 culture grown in the presence and absence of sub-MIC of the antimicrobial was centrifuged, and the supernatant was filtered through a 0.22 μm filter. Three volumes of chilled 100% ethanol were added to the filtered supernatant and incubated overnight at 2°C to precipitate the EPS. The ethanol wash and centrifugation steps were repeated thrice (Huston et al., 2004; Al-Shabib et al., 2016). EPS was quantified using the colorimetric phenol-sulfuric acid method as previously described (DuBois et al., 1956).
Autoinducer-2 assay
To determine whether Autoinducer-2 (AI-2) production in the E. coli O157 EDL 933 is affected by subinhibitory concentrations of the novel antimicrobial, the AI-2 assay was performed as previously described (Duan and March, 2010; Castillo et al., 2015). Briefly, approximately 7 Log CFU/mL of E. coli O157 were inoculated and cultured in LB broth with 0.5% (w/v) glucose containing 0%, 0.0625%, and 0.125% of the antimicrobial at 37°C for 6 h. Vibrio harveyi BB170 was grown overnight at 30°C with agitation in Autoinducer Bioassay medium (AB) and diluted 1:3000. Cultures to be tested for CAI-1 activity were centrifuged (12,000 g for 5 min). The supernatant was filtered, and 20 μL of cell-free supernatant was added to 80 μL of diluted V. harveyi BB 170 reporter culture in a sterile 96-well plate and incubated at 30°C with shaking at 500 rpm. The luminescence values of the samples were measured in an automatic plate reader (FLUOstar Omega; BMG LABTECH) every 0.5 h. The analysis was repeated thrice. In addition, the E. coli cultures were enumerated by plating on brain-heart infusion agar. The aforementioned concentrations of the antimicrobial were also tested against growth of V. harveyi BB170 to ensure that decrease in bioluminescence was not caused by growth inhibition.
Statistical analysis
All experiments were performed in triplicate. To measure the statistical significance we used two-tailed Student's t-test. A group difference was assumed to be statistically significant when p < 0.05. All results were expressed as mean ± SD.
Results
Antimicrobial activity and mechanism
The antimicrobial exerted a potent effect against all E. coli O157 strains tested, with MIC and MBC values of 0.5 and 0.75% (v/v) of the natural antimicrobial. The antimicrobial activity can be attributed to the lactic acid, citric acid, and citrus extract present in the antimicrobial solution. The results revealed (Fig. 1) that the electric conductivity of all treated samples increased significantly (p < 0.05) with exposure time (2–24 h) and with increasing concentrations of the antimicrobial (p < 0.05). These results indicate that the gradual increase in cell membrane permeability leads to cellular leakage. Membrane integrity was also studied by monitoring protein (Fig. 2A) and nucleic acid release (Fig. 2B) in terms of 260 nm absorbing materials. When the antimicrobial was applied at MIC and MBC levels, protein release was found to progressively increase with time (p < 0.05). In addition, when the antimicrobial was applied at the MBC level, protein release was significantly higher compared to the MIC treated E. coli O157. A very similar trend was observed for nucleic acid release (OD260nm). Application of the natural antimicrobial at MIC and MBC resulted in 5- to 12-fold increase in 260 nm material release (p < 0.05) after 24 h. There was also a gradual increase in the 260 nm material release during the 24 h incubation, with the release of MBC treated cells being always higher compared to the MIC treatment (p < 0.05). Protein and nucleic acid release for control samples remained stable in all cases. The above results suggest that membrane structural damage is taking place at both MIC and MBC for all three E. coli O157 strains tested. Since the three strains had the same MIC and MBC values and exhibited similar behavior in terms of cell membrane integrity and permeability, only E. coli EDL 933 was used in subsequent experiments as it is widely used in other studies thus allowing better comparisons.
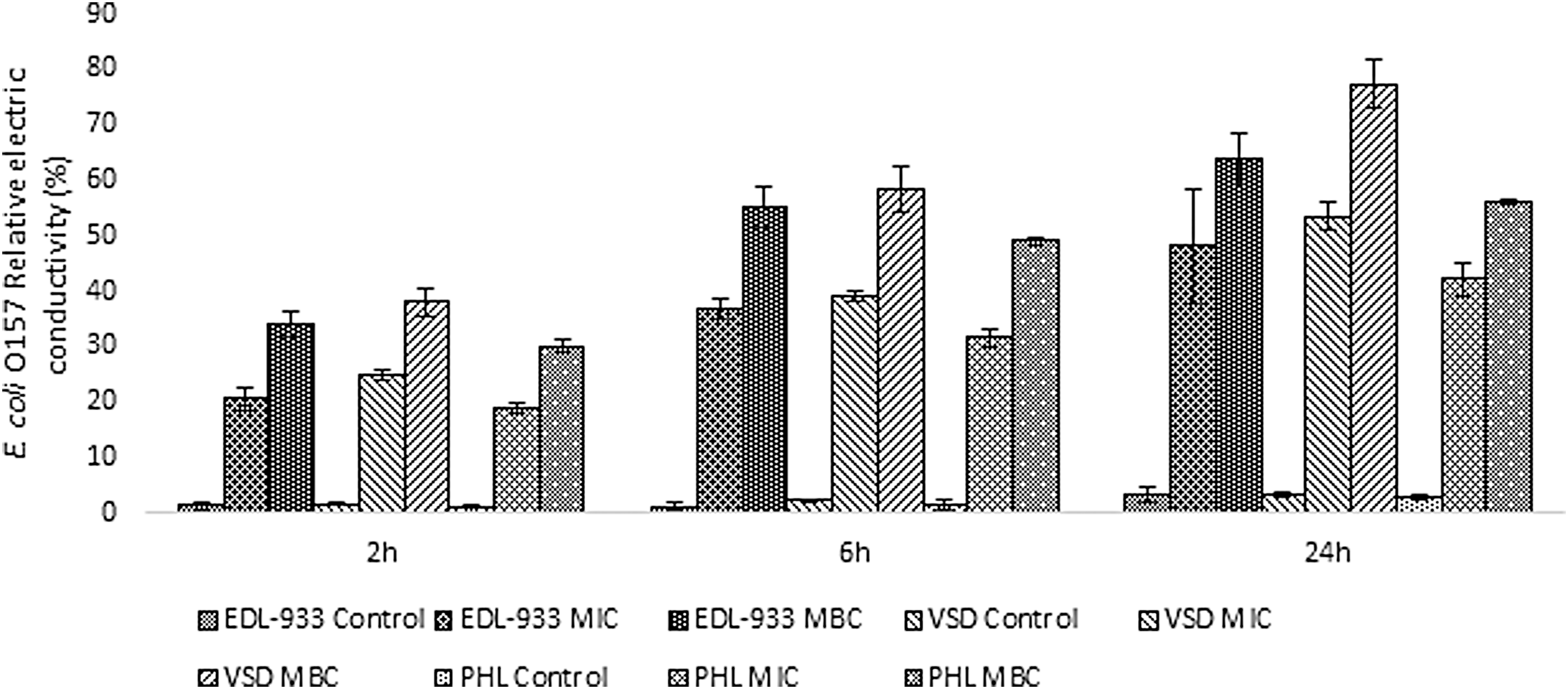
Effect of different concentrations of the antimicrobial on the three E. coli O157:H7 strain cell permeability as determined by measuring changes in relative electric conductivity. Each point represents the mean ± standard deviation. MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration.
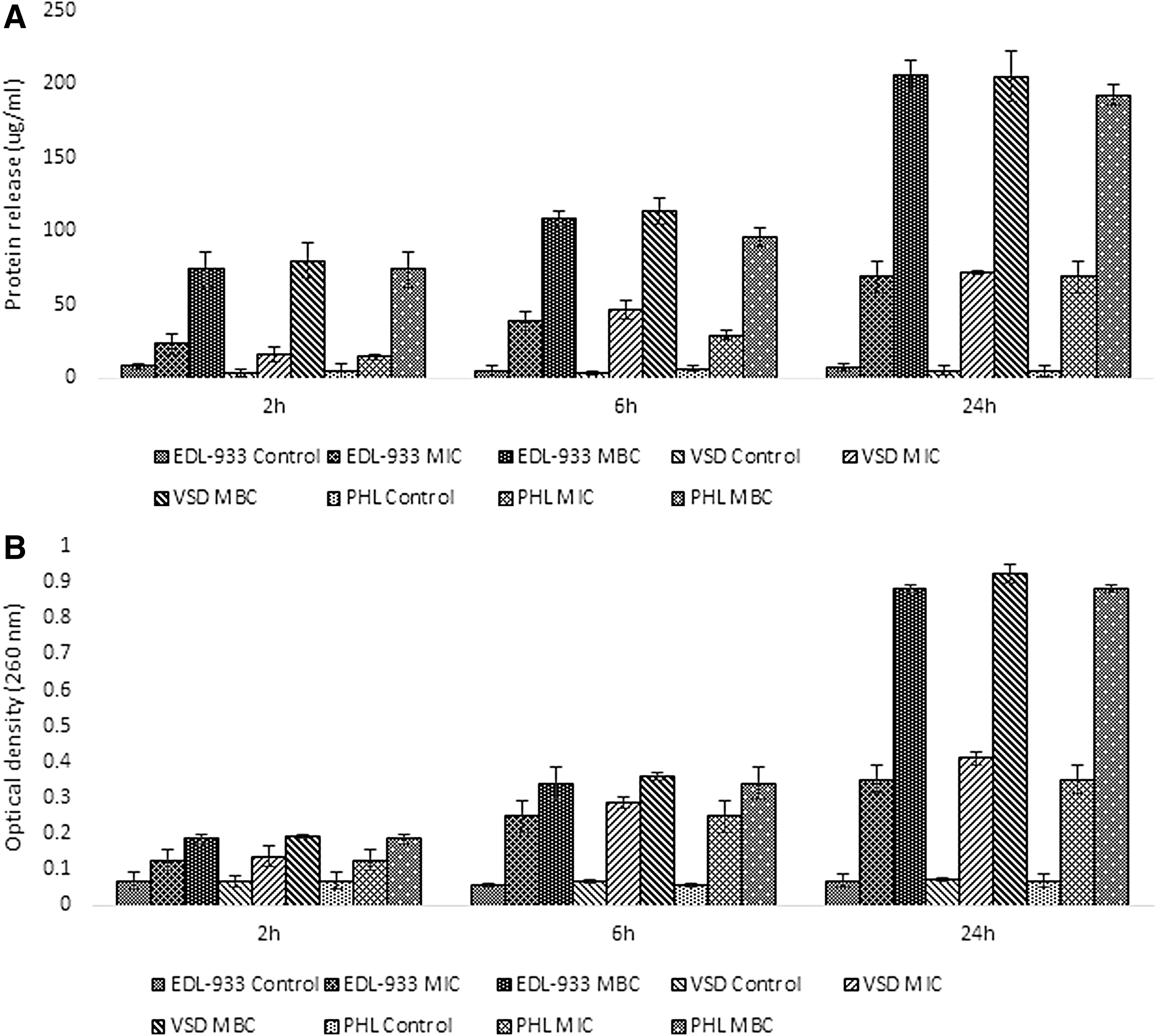
Effect of different concentrations of the antimicrobial on the three E. coli O157:H7 strains protein
Growth curves
Growth curves revealed complete inhibition of growth at MBC and MIC, as expected. The 1/2 MIC (0.25% v/v) caused a prolongation of the lag phase of all three E. coli O157 strains tested for 1 h, whereas 1/4 MIC (0.125% v/v) and 1/8 MIC (0.625% v/v) showed a very similar growth curve to the control showing that thesis concentrations had no effect on growth (Fig. 3). Therefore these two concentrations were selected to study the effect on motility, biofilm formation, exopolysaccharide production, and QS.
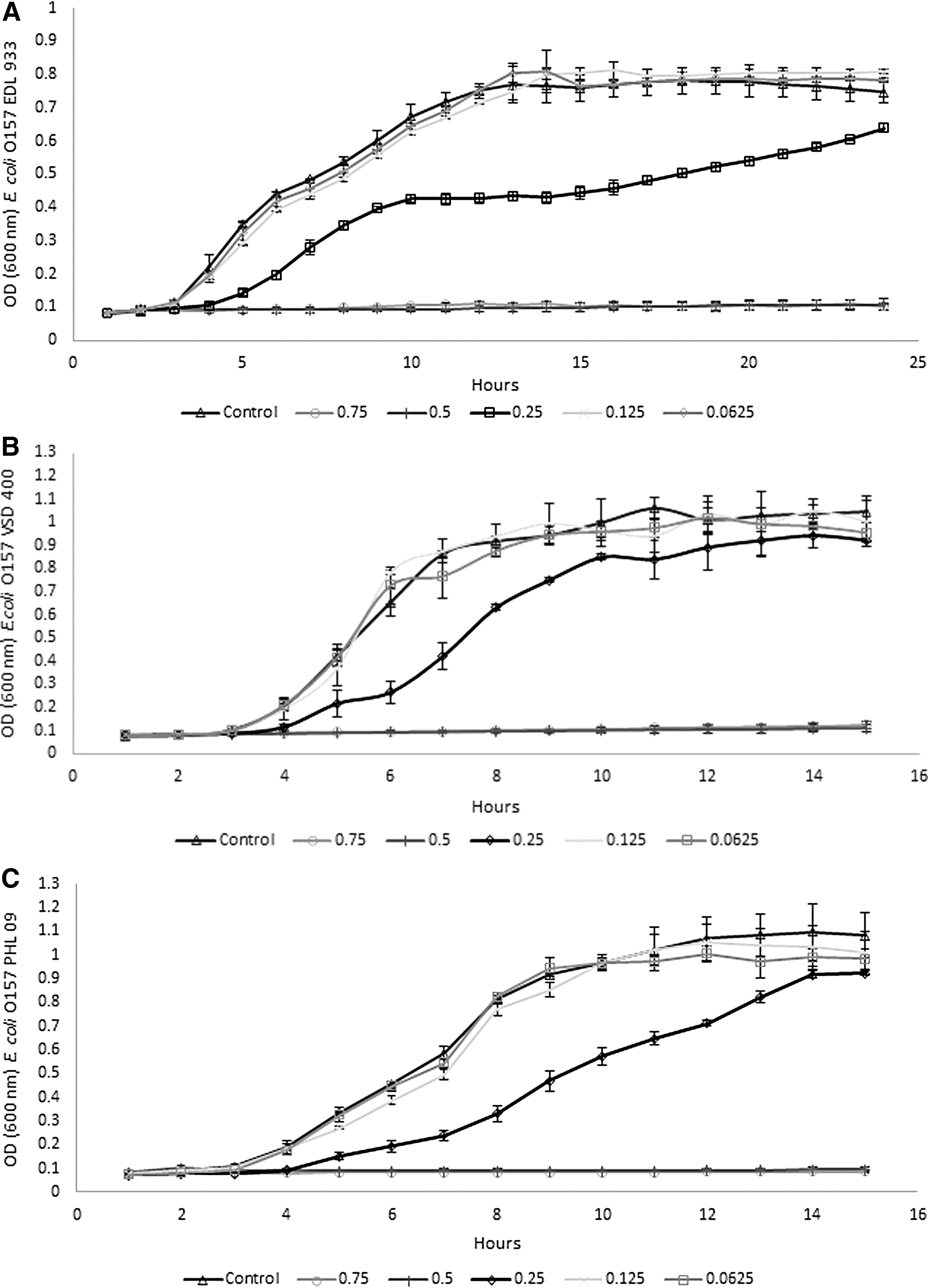
Growth of E. coli O157:H7 EDL 933
In vitro rumen system
Application of the antimicrobial at the MIC level did not influence the growth of the pathogen, which was very similar to the control (p > 0.05). Application at the MBC level resulted in a gradual reduction in the E. coli O157 counts, with the levels of the pathogen being significantly lower (5.15 ± 0.38 log CFU/mL) compared to the control and MIC treatment after 18 h of incubation (Fig. 4). After 24 h, the pathogen levels were reduced by 1.63 log CFU/mL for the MBC treatment. The 2 × MBC reduced the levels of E. coli O157 by ∼1 log CFU/mL within 4 h, compared to the control and MIC treatments (p < 0.05). The pathogen levels were reduced by 3.15 log CFU/mL after 24 h, which was significantly lower compared to the pathogen levels at 24 h for the MBC treatment (p < 0.05).
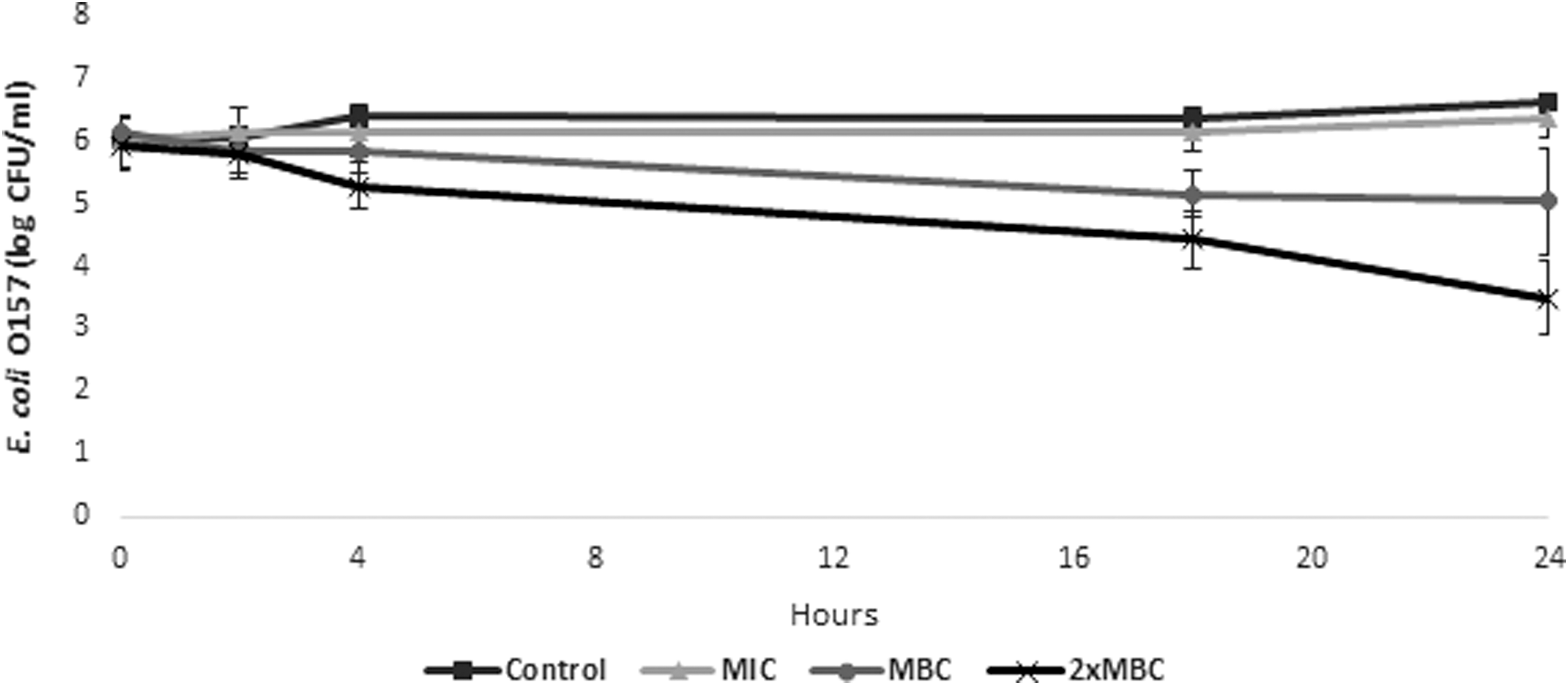
Levels of E. coli O157:H7 EDL 933 following treatment with different concentrations of the antimicrobial in an in vitro rumen system. Each point represents the mean ± standard deviation.
Effect on biofilm and motility
Addition of the natural antimicrobial at subinhibitory levels significantly decreased (p < 0.05) biofilm formation by E. coli O157, in a dose dependent manner by 43.20% and 52.13% compared to the control (Fig. 5). The effect of the novel antimicrobial on the pathogen motility is shown in Figure 6. The natural antimicrobial at both subinhibitory concentrations resulted in a clear reduction in the motility of the pathogen (34.11 ± 3.95 mm) compared to 68.77 ± 5.50 mm for the control (p < 0.05).
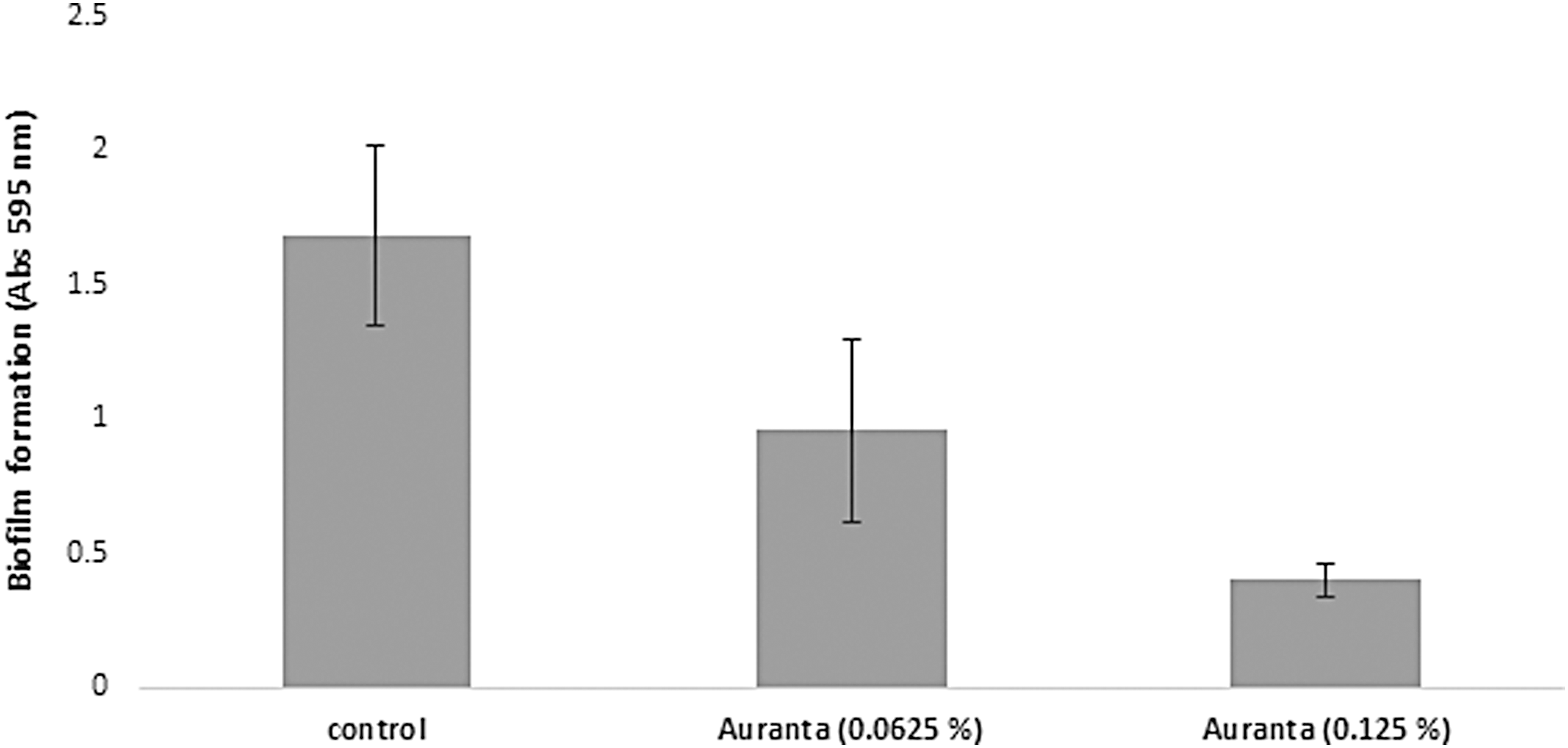
Effect of different subinhibitory concentrations of the antimicrobial on biofilm formation of E. coli O157:H7 EDL 933. Each point represents the mean ± standard deviation.
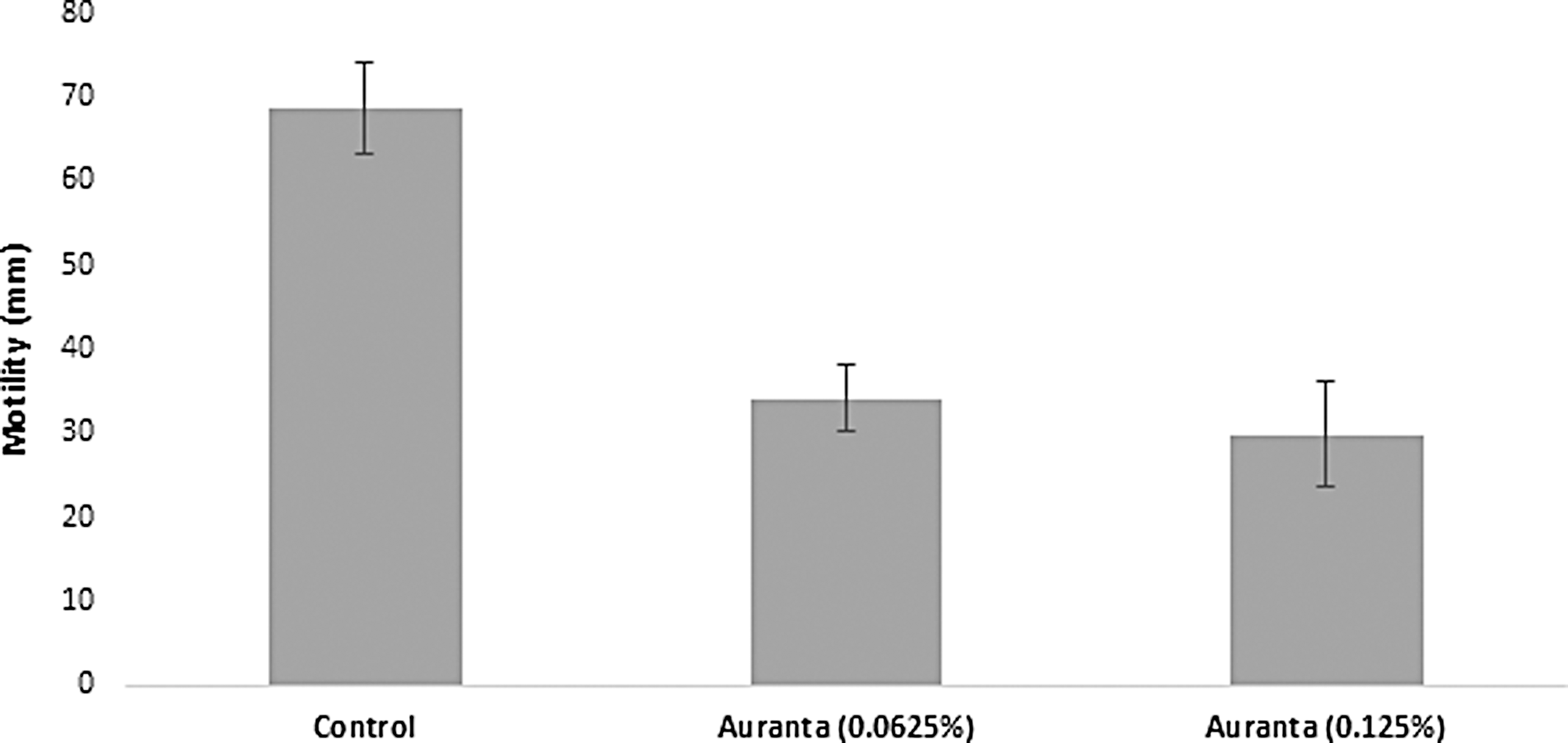
Effect of different subinhibitory concentrations of the antimicrobial on E. coli O157:H7 EDL 933 motility. Each point represents the mean ± standard deviation.
Effect on EPS production
Spectrophotometric determination of the extracted EPS revealed that the concentration of EPS decreased when the antimicrobial was applied (Fig. 7). A statistically significant reduction in the EPS production was recorded at both subinhibitory concentrations tested. The natural antimicrobial treated samples showed a 48.17% ± 5.67% and 52.07% ± 11.50% reduction in EPS production. However, no difference was found between the two treatments (p < 0.05).
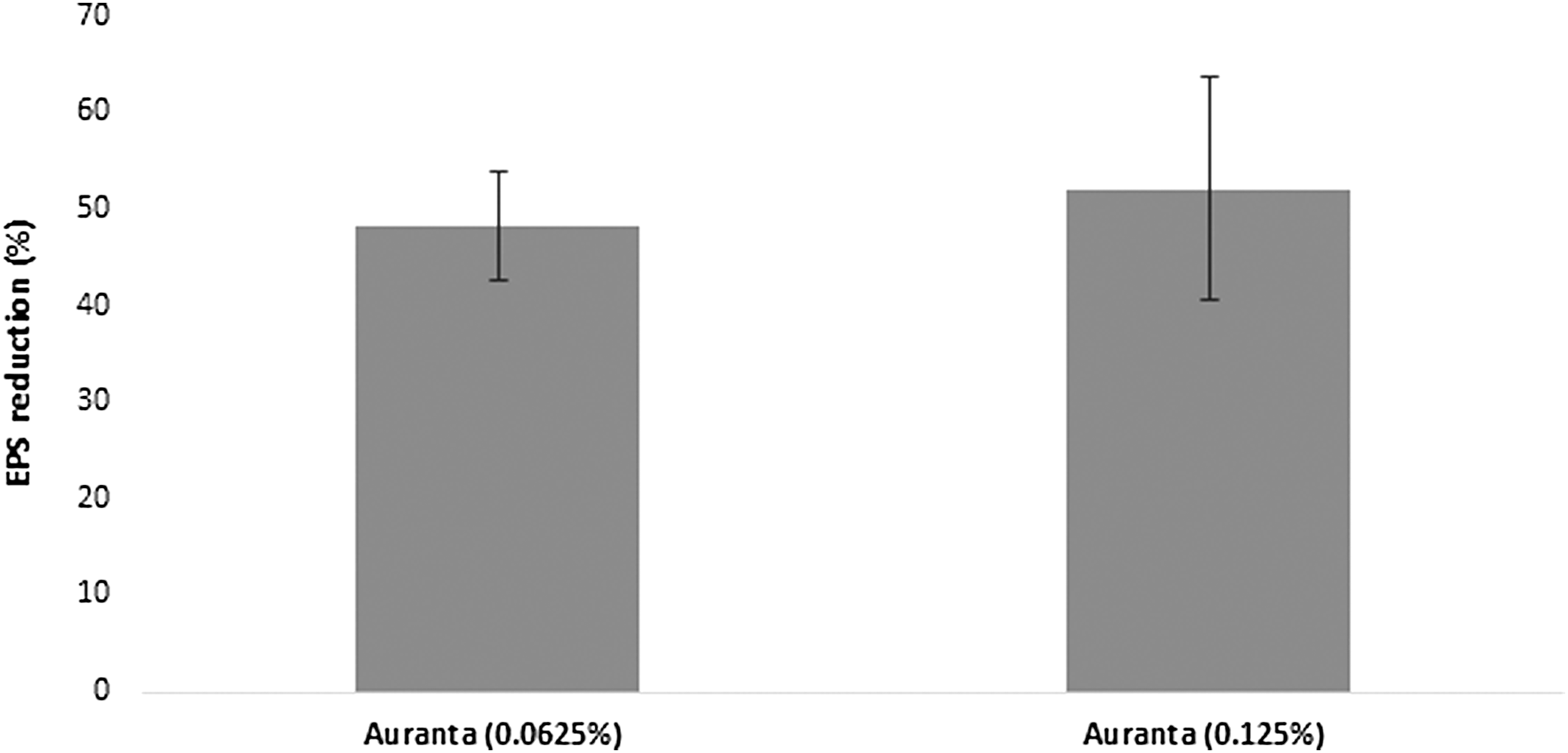
Quantitative determination of EPS inhibition E. coli O157:H7 EDL 933 after exposure to different subinhibitory concentrations of the antimicrobial. Each point represents the mean ± standard deviation. Results are expressed in percentages of the control treatment. EPS, exopolysaccharide.
Effect on QS
The effect of the natural antimicrobial on AI-2 production by E. coli O157:H7 was assayed using the V. harveyi BB170 reporter strain. The reporter strain is bioluminescent in response to the presence of AI-2. Results showed that a 6-h treatment of E. coli O157:H7 with subinhibitory concentrations of the antimicrobial did not result in any significant changes (p < 0.05) in AI-2 production compared to the control (Fig. 8A). The antimicrobial at the concentrations used (0.0625% and 0.125%) did not have any effect on the growth of E. coli O157 (Fig. 8B). In addition, the antimicrobial showed no antimicrobial activity against V. harveyi at the concentrations used in this study (results not shown).
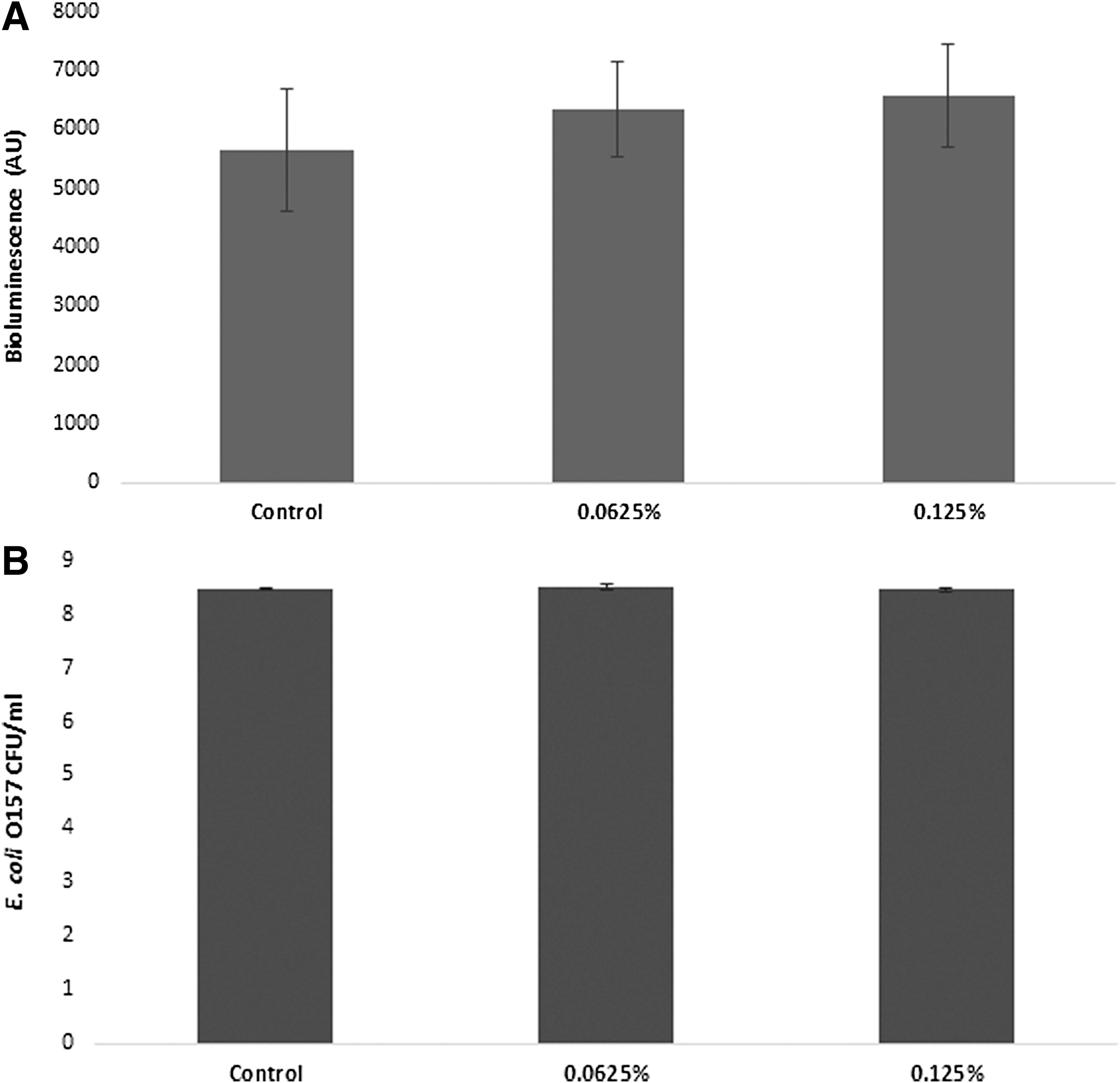
Production of the AI-2 by E. coli O157:H7 EDL 933 after exposure to different subinhibitory concentrations of the antimicrobial
Discussion
Enterohemorrhagic E. coli O157:H7 infection is a worldwide problem linked to many foodborne outbreaks (Vygen-Bonnet et al., 2017; CDC, 2018). Since ruminants are an important source of this pathogen (EFSA, 2010), reducing its ability to reside in the animal gastrointestinal tract would be desirable. In the present study we demonstrated that the natural antimicrobial is effective against E. coli O157:H7 with MIC and MBC values of 0.5% and 0.75% (v/v), respectively.
When bacteria are exposed to antimicrobial agents, different physical changes occur to their cell membranes (Bajpai et al., 2013). We show that the bactericidal effect observed is linked to the ability of the antimicrobial to disrupt E. coli O157:H7 membranes as described in Figure 1. This indicates a potential leakage of ions even at the MIC level because the bacterial membrane acts as a barrier to the movement of electrolytes that take part in cell functions and metabolism (Diao et al., 2014). Different studies have investigated the effect of plant extracts on E. coli O157. Exposing E. coli O157 to eugenol has also been observed to cause membrane damage and subsequent leakage of intracellular components (Di Pasqua et al., 2007).
Commercial citrus fruit extracts have also been found to cause physical damage to Salmonella enterica and E. coli membranes and leakage of cellular materials (Alvarez-Ordonez et al., 2013).
Stratakos et al. (2018) also found that carvacrol was able to increase cell membrane permeability in different STEC strains leading to cell death. Release of proteins and nucleic acids (260 nm absorbing materials) is used as an indirect way to assess membrane damage after application of an antimicrobial treatment (Bajpai et al., 2013; Stratakos et al., 2018). The natural antimicrobial caused a rapid release of proteins and 260 nm absorbing material, which increased over time, suggesting damage to the cytoplasmic membrane of the pathogen, which is consistent with the increase in relative electric conductivity. Zhang et al. (2017) also observed a dose dependent increase (0, MIC, 2 × MIC) in the protein and nucleic acid release of nonpathogenic E. coli cells after treatment with black pepper essential oil. Carvacrol and cinnamon essential oil have also been found to cause protein and nucleic acid release in E. coli and Staphylococcus aureus (Zhang et al., 2016). In general, the novel antimicrobial was found to interfere with E. coli O157 ion homeostasis and membrane integrity.
The in vitro rumen model showed that it is possible to reduce E. coli O157 EDL 933 shedding in ruminants with the natural antimicrobial. The 2 × MBC was used in the experimental design as it has been found in other studies that antimicrobials exhibit significantly reduced activity when applied in complex systems since these systems can contain different inhibitors (Burt, 2004; Rydlo et al., 2006). McDonnell et al. (2012) found that although caseicin A was able to reduce E. coli O157 in a broth system the same concentrations (≥2 mg/mL) had no effect when applied in an in vitro model system. The study of Rivas et al. (2010) showed that carvacrol applied at a concentration of 1000 μg/mL reduced E. coli O157 by >5 log CFU/mL in an in vitro rumen system after 24 h incubation. However, more work is required to elucidate further the effects of the antimicrobial on rumen microbiota and thus rumen fermentation.
It is possible that during ingestion of an antimicrobial present in the feed or water, its concentrations can be reduced due to partial inactivation when coming in contact with other compounds in the animal digestive system or simply by dilution. These lower concentrations might not have an effect on growth of pathogens residing in the gut but can potentially have an effect on different virulence factors (García-Heredia et al., 2016). Although a potential increase in the virulence of the pathogen due to exposure to subinhibitory concentrations of the antimicrobial will not cause any clinical symptoms to the animal, it could potentially result in cells with increased virulence being transferred to the meat or food producing surfaces during the slaughter process, increasing the risk and/or severity of human infection. Therefore, the present study also investigated the effect of subinhibitory concentrations of the antimicrobial on different virulence factors of E. coli O157:H7.
E. coli O157 is also able to form biofilms on various biotic and abiotic surfaces. These biofilms are resistant to host immune responses, antimicrobial agents, and other stresses (Kim et al., 2016). Therefore, subinhibitory concentrations of the natural antimicrobial were used to investigate the effect on biofilm formation. Both concentrations used (0.0625% and 0.152%) reduced E. coli O157 biofilm formation in a dose dependent manner without affecting the growth of planktonic cells (Fig. 5) Previously, eugenol was found to inhibit biofilm formation by Listeria monocytogenes and E. coli O157 (Perez-Conesa et al., 2006; Upadhyay et al., 2013). Similarly, subinhibitory concentrations of citral have been found to inhibit Cronobacter sakazakii biofilm formation at 12°C and 25°C (Shi et al., 2017). Carvacrol has also been found to inhibit biofilm formation in Campylobacter jejuni (Burt et al., 2014).
Motility plays an important role in E. coli biofilm formation (Ren et al., 2001) and also contributes to colonization of host tissues and enables pathogens to move toward favorable environments and away from repellents (Zhu et al., 2015). Therefore, the motility of E. coli O157:H7 during exposure to subinhibitory concentrations of the antimicrobial was also investigated. Both subinhibitory concentrations of the antimicrobial significantly reduced the motility of the pathogen (Fig. 6). The presence of functioning flagella is required for motility and biofilm formation in Gram-negative bacteria (Chelvam et al., 2014). Burt et al. (2016) showed that organic acids, specifically cinnamic (0.2%), lactic (0.6%), and propionic acid (0.4%), can reduce motility in Salmonella Typhimurium. Mith et al. (2015) also found that oregano essential oil and carvacrol are able to significantly downregulate genes involved in motility of E. coli O157:H7. Carvacrol has also been found, after overnight incubation, to prevent the development of flagella in E. coli O157:H7 leading to aflagellate and nonmotile cells. Coumarins were also found to suppress curli genes and motility genes by causing reductions in fimbria production and swarming motility in E. coli O157:H7 (Lee et al., 2014). Thus, a possible explanation for the reduced motility observed is inhibition of the flagellar synthesis by the subinhibitory concentrations of the antimicrobial.
To further elucidate the effect on E. coli O157:H7 virulence, EPS production by treated and nontreated bacteria was determined. E. coli O157:H7 produces EPS which acts as a physical barrier to protect cells against different stresses. EPS is also involved in cell adhesion and the formation of three-dimensional biofilm structures (Ryu and Beuchat, 2005). Therefore, any changes in the EPS production by the pathogen can have an effect on biofilm formation and architecture and also reduce resistance to drugs. In this study our results revealed that EPS was reduced significantly and to the same extent by the two subinhibitory concentrations of the antimicrobial (Fig. 7). The study of Al-Shabib et al. (2016) also showed that EPS was reduced significantly when Listeria monocytogenes and E. coli were treated with subinhibitory concentrations of zinc oxide nanostructures from Nigella sativa. Cellulose is one of the EPS components that enable biofilm formation in Enterobacteriaceae (Priester et al., 2006). Shi et al. (2017) found that treating Cronobacter sakazakii cells with citral downregulated the expression of bcsA and bcsG that encode the cellulose biosynthesis operon. In the present study, we hypothesize that the novel antimicrobial could potentially affect the synthesis of components (e.g., cellulose) which EPS consists of and thus lead to inhibition of E. coli O157:H7 biofilm formation.
Accordingly, our results showed that subinhibitory concentrations of the novel antimicrobial can reduce the pathogen's biofilm forming capability, motility, and exopolysaccharide production. The data also suggest that the antimicrobial does not interfere with the AI-2 synthetic pathway, and thus, the antibiofilm effect of the antimicrobial is not closely linked to AI-2 production.
Conclusion
Our study showed that the natural antimicrobial has a potent antimicrobial effect against all tested E. coli O157:H7 strains and exerts its antimicrobial effect by cell membrane disruption. Our data also show that the antimicrobial can reduce the levels of the pathogen in an in vitro rumen model and revealed that subinhibitory concentrations of the antimicrobial attenuate E. coli O157:H7 reduced biofilm formation, motility, and exopolysaccharide production, potentially reducing the risk and severity of disease. However, expression of virulence could be influenced by other factors during the slaughter process which should be also investigated in the future. Future studies should also elucidate the effect of the antimicrobial on Shiga toxin production as it is one of the main E. coli O157:H7 virulence factors. These findings indicate that the natural antimicrobial can potentially be used as an antimicrobial to reduce the presence of E. coli O157:H7 in the gastrointestinal tract of ruminants and help control human infection.
Footnotes
Acknowledgments
This study was supported by a grant awarded to NC and AS by Envirotech Innovative Products Ltd. Funders had no role in study design, data collection, analysis, interpretation, and publication.
Disclosure Statement
No competing financial interests exist.