
Abstract
Endophytic bacteria (EB) are a prospective source of natural and novel bioactive compounds with pharmaceutical relevance. In the present study, a total of 50 EB were isolated from the fruits and leaves of ginkgo tree (Ginkgo biloba L.), the only living species in the division Ginkgophyta and popularly known as a living fossil. All the isolated EB were screened for their antibacterial activity against five deleterious foodborne pathogenic bacteria namely Escherichia coli ATCC 43890, Salmonella Typhimurium ATCC 19586, Bacillus cereus ATCC 10876, Listeria monocytogenes ATCC19115, and Staphylococcus aureus ATCC 12600. Among the isolated EB, GbF-96, GbF-97, and GbF-98 exhibited antibacterial activity against all the pathogenic bacteria tested, with inhibition zone ranging from 33.47 to 9.55 mm. GbF-96, identified as Bacillus subtilis, exerted the highest antibacterial activity against all the tested bacteria. In contrast, the ethyl acetate extract of GbF-96 showed antibacterial activity against only B. cereus, E. coli, and Salmonella Typhimurium. Scanning electron microscopy results indicated cracked and irregular, swollen, shrunken, and lysed cell surfaces of the pathogenic bacteria treated with ethyl acetate extract of GbF-96 or B. subtilis, indicating that the metabolites of GbF-96 might penetrate the bacterial cell membranes and evoke pathways inducing cell lysis. Together, the data suggest that B. subtilis from G. biloba can be a potential candidate for controlling dreadful foodborne pathogenic bacteria, either by itself or by its metabolites.
Introduction
Endophytes living symbiotically with plants form a rich source of novel and bioactive compounds that are medicinally and economically significant to humans for their potential use in agriculture, medicine, and food industry (Strobel, 2003; Gutierrez et al., 2012; Kumar et al., 2016). Especially, endophytic bacteria (EB) can be used as active biological control agents (Kunoh, 2002; Backman and Sikora, 2008). Bioactive compounds obtained from endophytes have been used as key components in drug development (Demain and Sanchez, 2009; Brader et al., 2014; Kumari et al., 2015). Although various antimicrobial bioactive compounds are commercially available, there is a constant demand for the invention of new drugs with novel compounds (Kaaria et al., 2012), to overcome the growing drug resistance in different disease-causing pathogens, acquired by virtue of regular and irrational usage of various types of antibiotics.
Recently, foodborne diseases that are mainly caused by bacteria and viruses have become a major concern (Boore et al., 2010). The Centers for Disease Control and Prevention (CDC) in United States have estimated that 90% of all illnesses are caused by bacteria and viruses, including Escherichia coli, Listeria, Staphylococcus aureus, Salmonella, Campylobacter, Clostridium perfringens, and Norovirus (Deak, 1996; Addis and Sisay, 2015). With growing consciousness about food consumption, consumers now prefer natural source food products, to meet their demands for safer and preservative-free food. Despite being moderately harmful, food additives or preservatives are regularly used to preserve and prolong the shelf life of food products (Pandey et al., 2014). Due to the increasing microbial resistance against antibiotics, natural alternatives are becoming important; natural bioactive compounds, isolated from microbes, are being considered as prospective resources of food additives in food industry (Wiyakrutta et al., 2004). Endophytic microorganisms can be potential natural resources of bioactive antimicrobial compounds for controlling foodborne pathogens and/or toxin-producing microbes in foods (Strobel, 2003). There is an urgent need to discover unique antibacterial agents for controlling foodborne and related diseases (Četojević-Simin et al., 2010; Khaled et al., 2016).
Ginkgo biloba, commonly known as the maidenhair tree, is the only living species in the division Ginkgophyta (Singh et al., 2008). It is one of the oldest living tree species on earth and found in fossils dating back to >200 million years and, hence, referred to as a living Mesozoic fossil. Different tissues of this tree have long been used in China as a traditional medicine for treating various disorders (Chen et al., 2007), also in Korea and Africa as a folk medicine. The leaf extract is also one of the best-selling herbal supplements in the United States and Europe. Previous studies have reported that G. biloba plants are potent enough to affect important aspects of human physiology by refining the blood circulation to the tissues, including the brain, thereby enhancing cellular metabolism (Gaby, 1996).
Endophytic fungi, isolated from G. biloba, have been reported to produce important bioactive secondary metabolites (Xiao et al., 2013; Zhang et al., 2013). Although Chaetomium globosum and Bacillus amyloliquefaciens synthesized bioactive compounds that can defend host plants against pathogenic fungi and bacteria as bio-control agents (Yang et al., 2015), there has been no report of isolating EB from G. biloba to evaluate their antimicrobial activities. In this study, instead of identifying fungi to produce antimicrobial activities, we attempted to isolate EB from different parts of G. biloba to evaluate their antibacterial capacity against various foodborne pathogenic bacteria, including two Gram-negative bacteria, E. coli and Salmonella Typhimurium, and three Gram-positive bacteria, Bacillus cereus, Listeria monocytogenes, and S. aureus. The newly isolated EB with high antibacterial activities against foodborne pathogenic bacteria can be a powerful resource to control the pathogens, either by itself or by its active compounds.
Materials and Methods
Isolation and characterization of EB
Fresh leaves and fruits of G. biloba L. were collected from the campus of Yeungnam University, Gyeongsan, Republic of Korea, during October 2014. The tissues were collected and washed thoroughly under running tap water. EB were isolated as per the protocol reported by Costa et al. (2012) with a little modification. The washed tissues were dried, weighed, and sterilized, using 70% alcohol for 60 s, followed by 2% sodium hypochlorite for 3 min, and 100% ethanol for 30 s. The surface-disinfected samples were rinsed twice with sterilized distilled water, dried with sterilized filter papers, and ground with sterilized 3 mL/g of aqueous NaCl solution (0.9%) using a sterile mortar and pestle. The ground samples were incubated at 28°C for 3 h to allow complete release of EB from the host tissues.
For the isolation of EB, the ground extracts were diluted (10−1 and 10−2 times) with sterilized aqueous NaCl solution (0.9%) and spread onto different types of media plates, including 25% yeast extract, nutrient broth, and agar (YNA), 25% nutrient agar (NA), 25% de Man, Rogosa, and Sharpe (MRS) agar, and 25% Luria-Bertani (LB) agar, with a stainless steel spreader, to obtain single bacterial colonies. The media were prepared by dissolving only 25% (weight) of the original ingredients in water. The plates were incubated for up to 15 days at 28°C, and all the colonies were counted in terms of colony-forming units (CFU) per gram of the fresh organ. Single colony isolation was repeated at least thrice for purification of each isolated EB. Morphological characterization was done by recording colony characteristics based on their form, margin, color, and elevation. The isolated EB were cultured in different broth media and maintained as 50% glycerol stocks at −80°C until use.
The isolated EB were identified based on the comparison of 16S ribosomal RNA (rRNA) gene sequencing done by a colony polymerase chain reaction using 27F-1492R primers by Cosmo Genetech. The sequencing data were aligned and phylogenetic tree analyzed by Molecular Evolutionary Genetics Analysis (MEGA) software (version 6.0). Phylogenetic tree analysis was accompanied by the neighbor-joining method (Tamura et al., 2013; Susilowati et al., 2015).
Screening of antibacterial potentiality of EB against foodborne pathogenic bacteria
Various types of foodborne pathogenic bacteria were used in this study to determine the antibacterial capacity of EB isolated from G. biloba. The current study included two Gram-negative bacteria, E. coli ATCC 43890 and Salmonella Typhimurium ATCC 19586, and three Gram-positive bacteria, B. cereus ATCC 10876, L. monocytogenes ATCC 19115, and S. aureus ATCC 12600. These foodborne pathogenic bacteria were obtained from the ATCC (Manassas, VA) and maintained as glycerol stocks at −80°C.
Antibacterial activity of all the isolated EB was tested according to the method reported by Roh et al. (2009) with minor modification. The isolated EB were cultured in 96-well plates with 10 μL individual bacterial strain in yeast extract and nutrient broth (YNB; Becton, Dickinson and Company) and incubated for 48 h at 28°C. Next, 10 μL of EB culture at OD600nm = 1.0 was incubated in 25% YNA medium (containing 1.5% agar) for 36 h at 28°C, and the antibacterial activity was screened by the spot-on-lawn method (Roh et al., 2009). The spots of EB, grown on the plates, were killed by the addition of 1mL chloroform on the inner face of the Petri dish covers and incubated for 15 min. The lids were kept open for 20 min to remove any trace of chloroform. The Petri plates were then treated with ultraviolet light for 15 min to kill the bacteria completely. After the foodborne pathogens were grown in YNB medium at 37°C overnight, 30 μL cultures were suspended in 10 mL fresh YNA medium (0.75% agar) at 55°C and poured slowly and uniformly on the patch plate, without disturbing the plates. After solidification, all the Petri plates were sealed and incubated at 37°C for 24 h, and the diameter of inhibition zone was measured by an electronic digital caliper. Kanamycin and streptomycin at 10 μg/disc were used as the standard reference antibiotics.
Antibacterial activity of EB solvent extract against foodborne pathogenic bacteria
Ethyl acetate solvent extraction of EB was conducted according to the protocol by Zubair et al. (2011). After the selected EB were subcultured on YNA medium (1.5% agar) for 36 h, a single colony was picked and inoculated in 200 mL YNB in 1 L Erlenmeyer flasks with continuous shaking (180 rpm) at 28°C for 36 h. The culture broths were centrifuged at 2000 g for 15 min at 4°C, the supernatant was collected, mixed with an equal volume of ethyl acetate, and kept overnight. Ethyl acetate layer was selectively collected using a separating funnel. The ethyl acetate extract (5 mg/well) was checked for its antagonistic activity against the foodborne pathogens by the agar well diffusion assay, and the diameter of inhibition zone was measured with an electronic digital caliper.
The cells harvested as pellets were suspended with 20 mL methanol, sonicated in a bath sonicator (WUC-A03H; Daihan Scientific) for 15 min to break the cells, and incubated overnight, and the supernatant was collected by centrifugation at 2000 g for 15 min at 4°C. The extract was dried in a rotary evaporator (A-1000S; EYELA) at 50°C, redissolved in 2 mL methanol, and dried at room temperature. The amount of extract was determined and dissolved in methanol with 5% dimethyl sulfoxide (DMSO) to make the desired concentration (0.1 g/mL).
Determination of the minimum inhibitory concentration and minimum bactericidal concentration
The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of ethyl acetate extract of antibacterial EB against the five foodborne pathogenic bacteria were determined by the twofold dilution method (Patra et al., 2015), where dilutions were made by mixing the ethyl acetate extract in 5% DMSO (in methanol); 5% DMSO was used as the control. Different dilutions of the ethyl acetate extract in the YNB medium were prepared, and the lowest concentrations of ethyl acetate extract that inhibited the visual growth of the tested foodborne pathogenic bacteria were defined as the MIC, expressed in mg/mL. The MIC and the next higher concentration of the sample were selected, spread on the YNA Petri plates, and incubated at 37°C for 24 h; the lowest concentration of the extract that did not allow for any growth of the pathogenic bacteria on the 25% YNA plates was defined as the MBC, expressed in mg/mL.
Scanning electron microscopy analysis
The effects of ethyl acetate extract of antibacterial EB were determined from the scanning electron microscopy (SEM) images of surface morphology of the foodborne pathogenic bacteria, including B. cereus, E. coli, and Salmonella Typhimurium. The pathogenic bacteria were treated with either 5% DMSO as the control or the MICs of GbF-96 ethyl acetate extract, and their morphology was analyzed using the standard protocol by Bajpai et al. (2009).
Statistical analysis
The results of all experimental data are expressed as the mean ± standard deviation from three independent replicates for each experiment. Statistical analysis was conducted by one-way analysis of variance, followed by Duncan's multiple range test at p < 0.05 using the Statistical Analysis Software (SAS; Version: SAS 9.4; SAS Institute, Inc., Cary, NC).
Results
Isolation and screening of antibacterial EB
Fifty EB were isolated from fresh leaves and fruits of G. biloba in different media. The EB were obtained either from tissues, such as leaves (11 isolates, 22%) and fruits (39 isolates, 78%), or from different media such as LB (30 isolates, 6%), 25% YNA (19 isolates, 38%), and MRS (1 isolate, 2%) (Supplementary Table S1). Tissue-specific bacterial cell numbers were counted as 1.8 × 103 CFU/gin leaves and 6.0 × 104 CFU/gin fruits (Supplementary Table S1). The EB exhibited various forms, elevation, margins, and colors on growth culture media (Supplementary Table S2). Almost all the isolated EB were either circular or irregular, appearing as white, off-white, transparent, brown, light brown, light yellow, yellow, or orange colonies with almost complete margins or undulate margins. More than 50% isolates were raised in elevation, while the rest were flat and convex.
Evaluation of antibacterial activity of EB
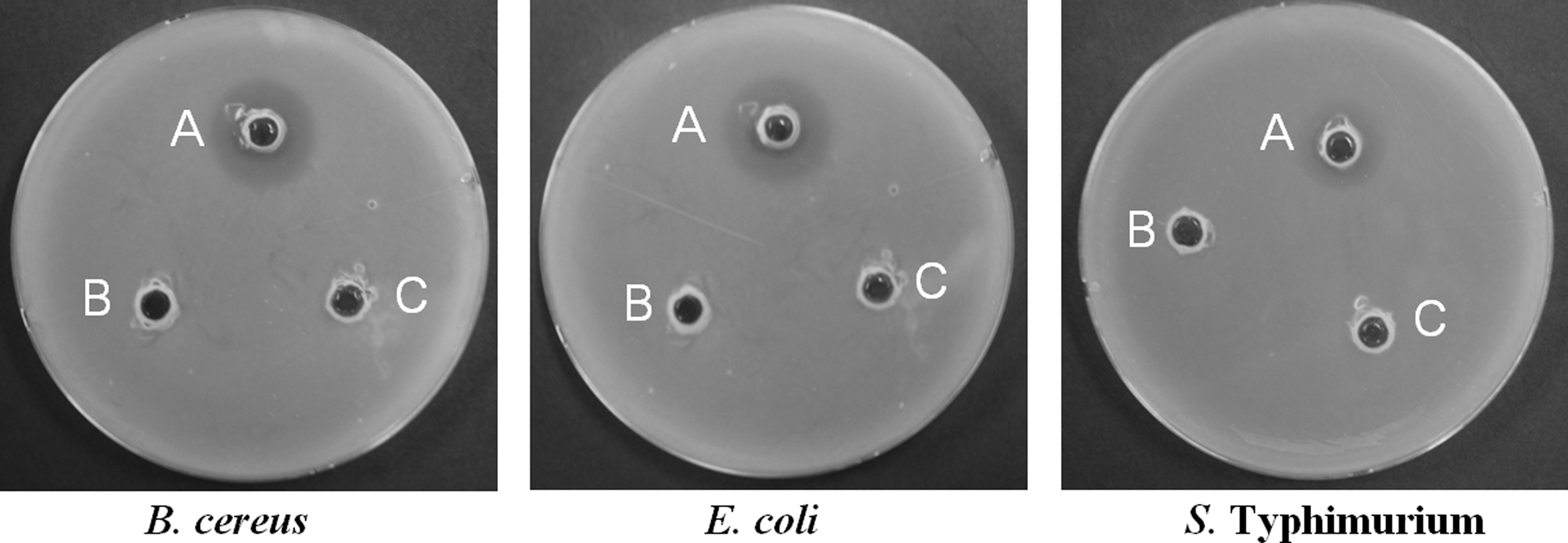
Among the isolated EB, total 12 EB showed positive antibacterial activity with different diameters (Table 1), but the rest 38 EB did not have any antibacterial activity. Especially, three isolates GbF-96, 97, and 98 having the highest antibacterial potentialities were selected for further study (Table 2 and Fig. 1). GbF-96 possessed the highest antibacterial activity against all the tested foodborne pathogens, including B. cereus, L. monocytogenes, S. aureus, E. coli, and Salmonella Typhimurium, with inhibition zones of 31.27 ± 3.01, 26.12 ± 0.95, 33.47 ± 0.95, 31.83 ± 0.01, and 29.51 ± 0.81 mm, respectively (Table 2). GbF-98 showed lower antibacterial activity against S. aureus and Salmonella Typhimurium with inhibition zones of 9.65 ± 0.18 and 9.55 ± 0.26 mm, respectively, compared to GbF-96 (Table 2). GbF-97 exerted antibacterial activity only against S. aureus (10.02 ± 0.30 mm of inhibition zone, Table 2).
Antibacterial activity of the endophytic bacterium Bacillus subtilis GbF-96 isolated from Ginkgo biloba against foodborne pathogenic bacteria. GbF-96 exerted antibacterial activity against all the foodborne pathogenic bacteria:
Antibacterial Screening of Endophytic Bacteria Isolated from Ginkgo biloba
The activity was estimated after 24-h incubation at 37°C by measuring the clear zone of EB inhibition: “−” indicates negative response (no activity during antibacterial screening test); “+” indicates positive response (<9 mm diameter of inhibition zone); “++” indicates medium positive response (9–11 mm diameter of inhibition zone); “+++” indicates strong positive response (>11 mm diameter of inhibition zone).
EB, Endophytic bacteria; GbF, G. biloba fruit.
Antibacterial Activity of the Selected Endophytic Bacteria, Isolated from Ginkgo biloba, and Standard Antibiotics Against Foodborne Pathogenic Bacteria
Diameter of inhibition zone (in mm) from three independent replicates expressed as the mean ± standard deviation
Values with different superscript letters in the same column are significantly different at p < 0.05 by Duncan's test.
10 μg/disc were used as standard reference antibiotics.
−, No inhibition zone.
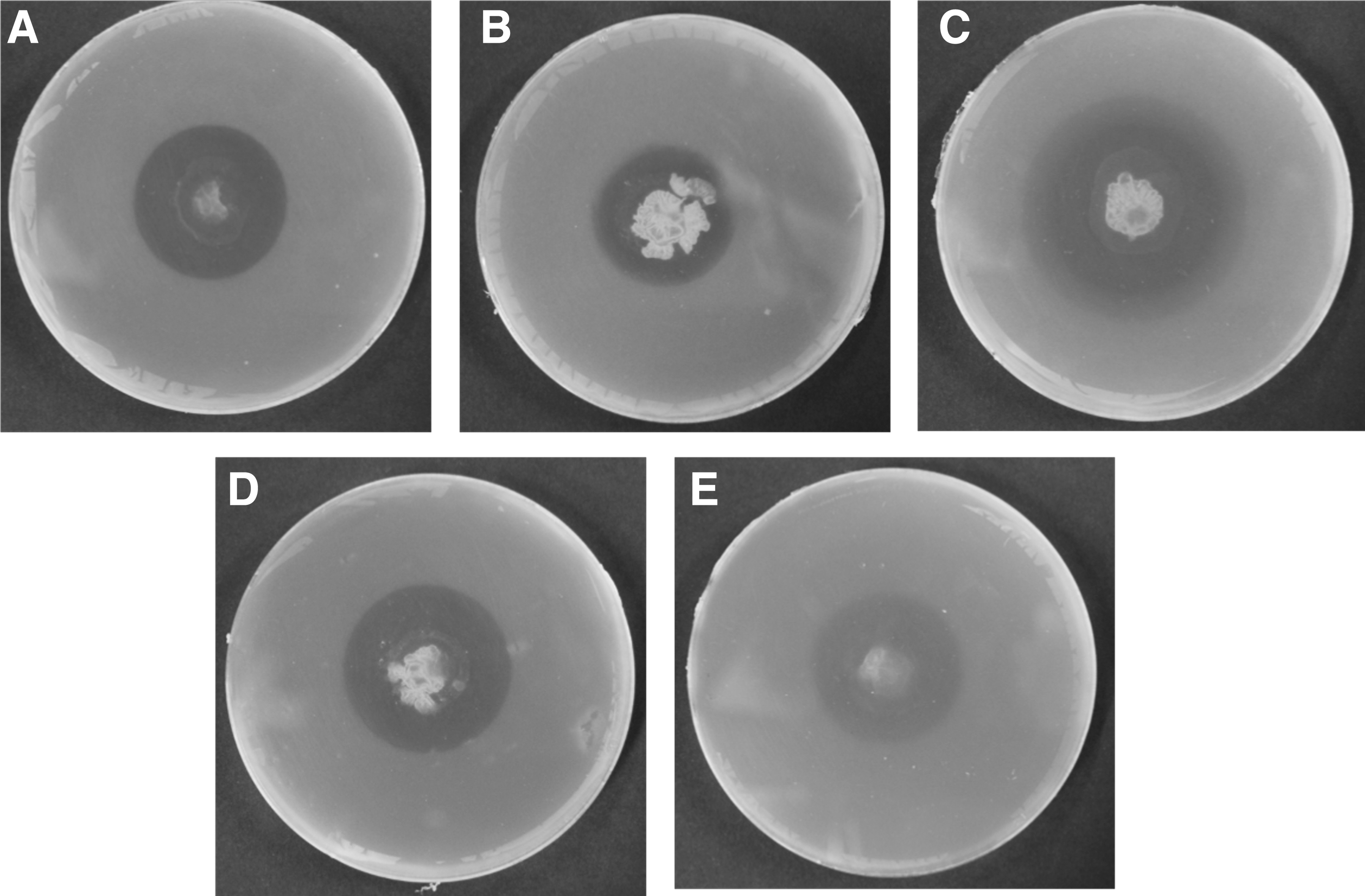
Antibacterial activity of the ethyl acetate extracts from the media broth or methanol extract from the bacterial cells was tested. The three EB (GbF-96, 97, and 98) were further tested for their antibacterial activity with the ethyl acetate extracts. In the assay of ethyl acetate extracts of the three selected EB, only GbF-96 showed antibacterial activity against the three foodborne pathogenic bacteria (B. cereus, E. coli, and Salmonella Typhimurium) (Table 3 and Fig. 2). Especially, the extract of GbF-96 exerted the highest antibacterial activity against B. cereus, with the inhibition zone of 17.54 ± 2.81 (Table 3 and Fig. 2). The methanol extract from the bacterial cells did not exert any antibacterial activity against the foodborne pathogens.
Antibacterial potentiality of the ethyl acetate extract of EB isolated from Ginkgo biloba against foodborne pathogenic bacteria, Bacillus cereus, Escherichia coli, and Salmonella Typhimurium, respectively. The EB were (A) GbF-96; (B) GbF-97; and (C) GbF-98. EB, Endophytic bacteria.
Antibacterial Potentiality of the Ethyl Acetate Extract of Endophytic Bacteria Isolated from Ginkgo biloba Against Foodborne Pathogenic Bacteria
Diameter of inhibition zone (in mm) from three independent replicates expressed as the mean ± standard deviation.
Values with same superscript letters are not significantly different at p < 0.05 by Duncan's test.
−, No inhibition zone produced.
Phylogenic analysis of the antibacterial EB
A phylogenetic tree constructed by analyzing the 16S rRNA sequences indicated that the antagonistic GbF-96 against all pathogenic bacteria was closely related to the species of Bacillus subtilis (Fig. 3). GbF-96 had the 99% highest similarity with the sequence of 16S ribosome of the B. subtilis IAM 12118 (accession No. NR 112116).

Neighbor-joining phylogenetic tree, based on 16S rRNA gene sequence analyses, showing the relationship among the isolated GbF-96 and the standard strains. Evolutionary distances were computed using the maximum composite likelihood method. The numbers at the nodes are bootstrap values (in %; based on 1000 samples). Genebank accession numbers are shown in parentheses. Bar: 0.0005 nucleotide substitutions per nucleotide position. rRNA, ribosomal RNA.
MIC and MBC of ethyl acetate extract
Since the ethyl acetate extract of GbF-96 was the only one to exert antibacterial activity against the three foodborne pathogens, its MIC and MBC against B. cereus, E. coli, and Salmonella Typhimurium were determined. The ethyl acetate extract of GbF-96 contained moderate-to-strong antibacterial effect against all the tested foodborne pathogenic bacteria. The MIC and MBC values of the ethyl acetate extract of GbF-96 ranged from 0.625 to 2.50 and 1.25 to 5.00 mg/mL, respectively (Table 4). The lowest values of MIC and MBC were against Salmonella Typhimurium ATCC 19586 (0.625 and 1.25 mg/mL, respectively) (Table 4).
Minimum Inhibitory Concentration and Minimum Bactericidal Concentration of the Ethyl Acetate Extract of Bacillus subtilis GbF-96 Against Foodborne Pathogenic Bacteria on Yeast Extract and Nutrient Broth Plates
MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration.
SEM analysis with the extract of GbF-96
The antibacterial effects of ethyl acetate extract of GbF-96 on the cell morphology of B. cereus, E. coli, and Salmonella Typhimurium were obtained by SEM analysis. Treatment of the pathogenic bacteria with ethyl acetate extract of GbF-96 induced cracked and irregular surfaces with an elongated, shrunk, lysed, and puffy morphology compared to the regular, uniform, and smooth surfaces in the untreated control bacterial cells (Fig. 4).
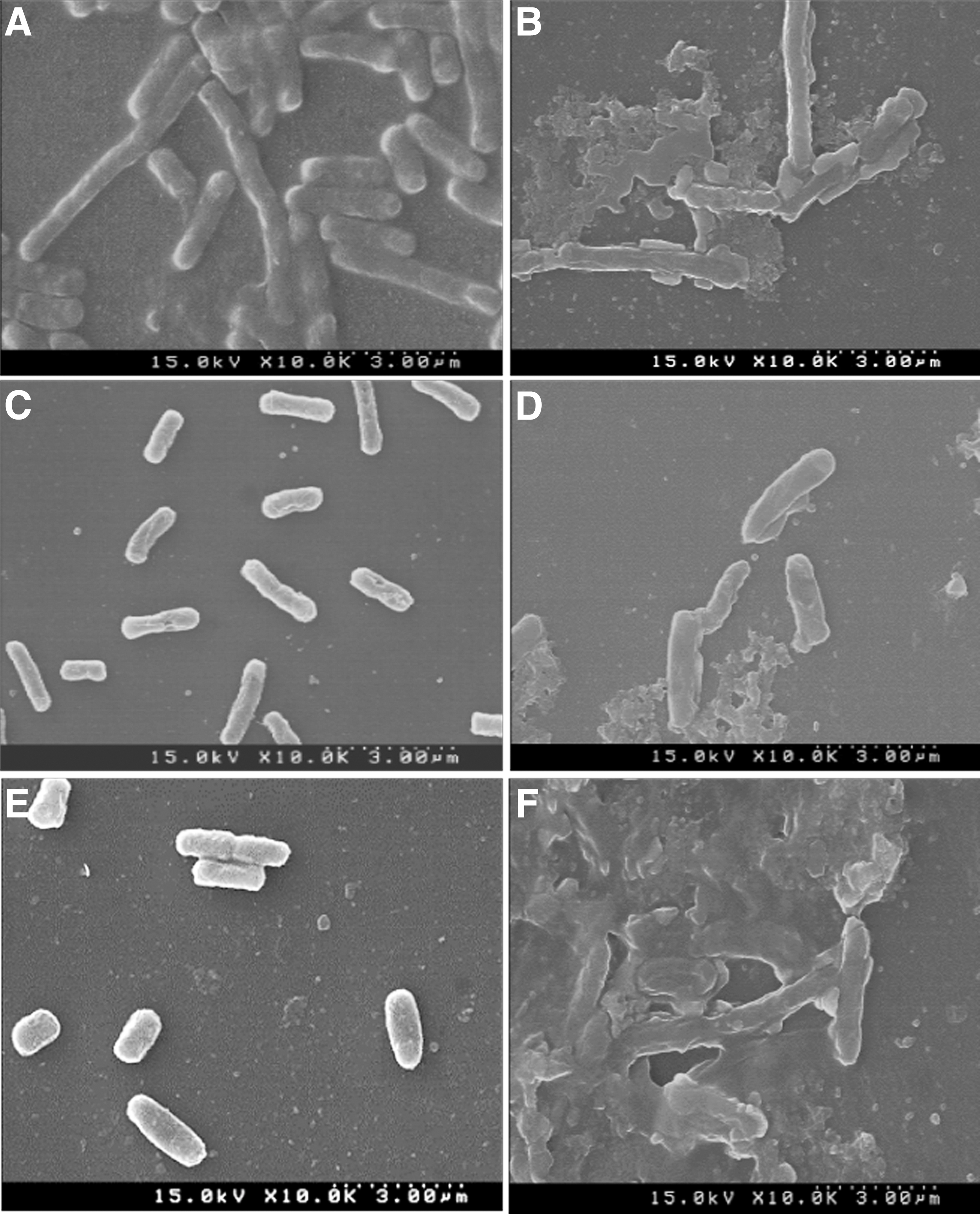
Scanning electron microscopy images of foodborne pathogenic bacteria treated with 5% DMSO as the control
Discussion
Food is an important medium for growth and spread of various types of foodborne pathogens and diseases under specific conditions (Torgerson et al., 2015). Food serves as an outstanding culture and protective medium for various pathogenic microorganisms, which may be capable of causing serious health problems such as foodborne diseases (Ganguly et al., 2012). Among the pathogenic microorganisms, the most important foodborne pathogenic bacteria, namely B. cereus, E. coli, L. monocytogenes, S. aureus, and Salmonella Typhimurium, may cause life-threatening diseases such as food poisoning, hemorrhagic colitis, bloody diarrhea, meningitis, osteomyelitis, endocarditis, gastroenteritis, and sepsis (Lim et al., 2010; Ganguly et al., 2012; Seo et al., 2016). Therefore, our research goal was to discover effective bioactive compounds from EB, isolated from Gingko tree, which would provide a new drug formulation for the economic and sustainable management of foodborne diseases.
Microbial endophytes are renowned as rich sources of natural and unique bioactive secondary metabolites with potential properties against pathogenic infections (Strobel, 2003; Krohn et al., 2007; Christina et al., 2013). Many of them can synthesize novel bioactive secondary metabolites that can be used for controlling foodborne pathogens. In this study, 50 EB were isolated from the leaves and fruits of G. biloba (Supplementary Table S1) and were found to range from 1.8 × 103 to 6.0 × 104 CFU in the culture media of different tissues (Supplementary Table S1). Similar data on the difference in densities and variety of bacteria had been reported earlier, based on the host plant species and related tissues, with varying environmental conditions, including the places and weather conditions (Quadt-Hallmann et al., 1997; Hallmann et al., 2001). All isolates having antimicrobial activity were derived from fruits, not from leaves. Although the leaves were also a substantial source of EB, only the fruits of G. biloba contained the EB with antimicrobial activity, suggesting the pulp of G. biloba fruits as an important resource for isolating EB with antibiotic activity.
Bacillus spp. represent a well-known and leading bacterial heterogeneous taxon with abundant spread in nature (Alina et al., 2015). Different Bacillus spp. are known as bio-control agents in the food industry since they are considered Generally Recognized As Safe (Pedersen et al., 2002; Romero et al., 2007). A number of species in the Bacillus genus, including B. subtilis, Bacillus licheniformis, Bacillus mojavensis, and B. amyloliquefaciens, are economically very important due to their capability of producing a variety of secondary metabolites (Gomaa, 2013; Kaewklom et al., 2013; Khochamit et al., 2015), which exert antibacterial activity against foodborne pathogenic microbes (Földes et al., 2000; Melnick et al., 2008; Torres et al., 2015). Among the Bacillus spp., B. subtilis can be classified as one of the most important species due to its ability to produce important secondary metabolites, including antimicrobial agents (Földes et al., 2000; Torres et al., 2015). The bioactive secondary metabolites such as bacteriocin, surfactin, and iturins are antimicrobial compounds produced by various B. subtilis strains (Földes et al., 2000; Sabaté et al., 2009; Compaoré et al., 2013; Torres et al., 2015).
In this study, ethyl acetate extracts of B. subtilis GbF-96 exerted strong antibacterial activity against B. cereus, E. coli, and Salmonella Typhimurium, but not against L. monocytogenes and S. aureus, which are controlled by the live GbF-96 (Figs. 1 and 2 and Tables 2 and 3). These data indicate that the mechanism for controlling the pathogenic bacteria by GbF-96 not only involved secondary metabolites but also other living components, such as antibacterial proteins or some yet unknown molecules, excluded during the process of ethyl acetate separation. High inhibition zones were reported against foodborne pathogenic bacteria such as S. aureus, L. monocytogenes, B. cereus, and E. coli in Bacillus spp. treatment (Sabaté et al., 2009; Lee et al., 2010; Torres et al., 2015). B. subtilis produces various kinds of antibiotics and antimicrobial secondary metabolites, including the bacteriocin sublancin 168 (Paik et al., 1998), bacillocin 22 (Zheng and Slavik, 1999), and subtilosin A (Shelburne et al., 2007), which exert differential levels of antibacterial activity, based on the pathogenic bacterial species, for example, E. coli, Salmonella enteritidis, Salmonella Typhimurium, and C. perfringens (Asai et al., 2002; Fairbrother et al., 2005). Another possibility may be the growth conditions, being differed by the growth on plates or in broth; therefore, further research would be required for the clear identification for the growth determinants.
The ethyl acetate extract of GbF-96 showed moderate-to-strong ranges of values of MIC and MBC against the foodborne bacteria (Table 4). High inhibition zones and the low MIC values have also been reported for Paenibacillus sp. against foodborne pathogenic bacteria, including B. subtilis (NCIM2063), S. aureus (MTCC 737), E. coli (MTCC739), and Pseudomonas aeruginosa (MTCC741) (Tayung et al., 2011; Gogoi et al., 2008). In addition, strong antimicrobial activity of Paenibacillus sp. against E. coli and Candida albicans has been reported (Aw et al., 2016).
Growth of foodborne pathogens in culture media may be suppressed by the diffusion of EB metabolites. B. subtilis GbF-96 exhibited robust antibacterial activity against the three tested foodborne pathogens, the possible reason being attributable to bioactive secondary metabolites. The metabolites might be simply passed into the bacterial cell membrane through the general bacterial porins, which are involved in several metabolic functions of the cell, or they might have generated various pores in the outer cell membrane of bacteria, thereby resulting in the leakage of internal substance to the exterior, subsequently leading to cell lysis and death (Kotzekidou et al., 2008). The mechanism was also validated by SEM analysis, suggesting that B. subtilis GbF-96 does possess potential bioactive compounds (Fig. 4). Cell morphology of the pathogens treated with ethyl acetate extract of B. subtilis GbF-96 was observed to be irregular, with rough surfaces, deformities, and eventual collapse. In agreement with our results, similar morphological deformities and collapse were also observed earlier in different test organisms treated with endophyte-produced metabolites (Mingma et al., 2014; Keerthi et al., 2016). Overall, the isolated GbF-96 demonstrated inhibitory effect on pathogenic bacteria, as confirmed by the severe morphological distortion on the cell surface of the three tested foodborne pathogenic bacteria, leading to destruction and cell lysis.
Conclusions
Endophytes produce a large amount of novel and bioactive secondary metabolites that are not only beneficial to the host plant but also economically important to humans for the potential applications in pharmaceutical and food industries. The newly isolated EB, B. subtilis GbF-96 isolated from G. biloba, can be a potential resource for investigating antibacterial agents against foodborne pathogenic bacteria applied as solvent extract or the pure bioactive compounds. Nevertheless, further studies would be required to identify and purify the bioactive metabolite(s) in large scale, for it to be used as an antibiotic, as well as for various biotechnological applications.
Footnotes
Acknowledgments
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Agri-Bio Industry Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA; 117044-3).
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.