
Abstract
Antimicrobial resistance (AMR) is a global issue for both human and animal health. Infections caused by antimicrobial-resistant bacteria present treatment option challenges and are often associated with heightened severity of infection. Antimicrobial use (AMU) in human and animal health is a main driver for the development of antimicrobial-resistant bacteria. Increasing levels of AMU and the development and spread of AMR in food-producing animals, especially in poultry and swine production, has been identified as a food safety risk, but dairy production systems have been less studied. A number of farm management practices may impact on animal disease and as a result can influence the use of antimicrobials and subsequently AMR prevalence. However, this relationship is multifactorial and complex. Several AMR transmission pathways between dairy cattle, the environment, and humans have been proposed, including contact with manure-contaminated pastures, direct contact, or through the food chain from contaminated animal-derived products. The World Health Organization has defined a priority list for selected bacterial pathogens of concern to human health according to 10 criteria relating to health and AMR. This list includes human pathogens such as the extended-spectrum beta-lactamase-producing Enterobacteriaceae (ESBL-E), which can be associated with dairy cattle, their environment, as well as animal-derived food products. ESBL-E represent a potential risk to human and animal health and an emerging food safety concern. This review addresses two areas; first, the current understanding of the role of dairy farming in the prevalence and spread of AMR is considered, highlighting research gaps using ESBL-E as an exemplar; and second, a New Zealand perspective is taken to examine how farm management practices may contribute to on-farm AMU and AMR in dairy cattle.
Introduction
Antimicrobial resistance (AMR) is a global issue for both human and animal health. Left unchecked it is estimated that by 2050, 10 million human deaths per year will be attributable to AMR globally (O'Neill, 2014). Infections caused by antimicrobial-resistant bacteria present treatment option challenges and are often associated with heightened severity of infection (Pitout and Laupland, 2008). The use of antimicrobials in human and animal health is considered the main driver for the development of antimicrobial-resistant bacteria (Laxminarayan et al., 2013). However, the reason for the emergence, persistence, and spread of AMR is more complex (Holmes et al., 2016). For example, the exposure to other compounds, such as heavy metals, biocides, and disinfectants may co-select for AMR (Horner et al., 2012; Jaglic and Cervinkova, 2012; Seiler and Berendonk, 2012). Because of the extensive use of antimicrobials in food-production animals, the contribution of the food–animal AMR nexus to the emergence and spread of AMR in the human population has also become an established consumer concern (George, 2018). Some studies suggest an epidemiological relationship between genetically similar antimicrobial-resistant bacteria found in both livestock and humans (Harrison et al., 2013; Liu et al., 2016). One such group of bacteria is the extended-spectrum beta-lactamase-producing Enterobacteriaceae (ESBL-E) (Wu et al., 2013).
Enterobacteriaceae are Gram-negative bacteria that form part of the natural intestinal microbiota of humans and dairy cattle as well as other animals, and are also able to persist within the environment. Some members of the Enterobacteriaceae family, such as Escherichia coli and Klebsiella pneumoniae, are opportunistic pathogens and can cause both hospital- and community-acquired infections such as sepsis and urinary tract infections (Pitout and Laupland, 2008; Heffernan et al., 2009; Freeman et al., 2012).
Enterobacteriaceae can acquire a wide range of AMR genes including resistance to colistin, mediated by the mcr1 gene (Liu et al., 2016), and resistance to tetracycline because of the tetABCDE determinants (Fluit et al., 2005). However, one of the most important and widespread mechanisms of AMR in Enterobacteriaceae is the production of ESBL enzymes. This review takes a wide definition of ESBL as described by Rubin and Pitout (2014) and Iredell et al. (2016) as being those beta-lactamases that provide resistance to the first- and third-generation cephalosporins, monobactams, and penicillins, but not the cephamycins. This includes variants of the CTX-M, SHV, and TEM (excluding parent type) enzymes, among other less frequent ESBL types as reviewed by Naas et al. (2008), but not the wider spectrum AmpC type beta-lactamases that have a broader activity against cephalosporins, providing resistance to first-, second-, and third-generation cephalosporins (Rubin and Pitout, 2014).
Genes encoding ESBLs are often associated with additional genetic determinants coding for other mechanisms of resistance, resulting in a multidrug-resistant (MDR) phenotype (Poirel et al., 2011; Ho et al., 2013; McGann et al., 2016). It is the development of MDR pathogenic strains of Enterobacteriaceae that is of particular concern to the health sector resulting in ESBL-E being identified as “critical” on the World Health Organization's (WHO) “Priority Pathogens List” (Tacconelli et al., 2018). ESBL-E pose a significant public health issue and have been detected in agricultural environments and associated food products. Therefore, we use ESBL-E in this review as an exemplar to investigate the role of dairy farming in the prevalence and spread of AMR.
In dairy cattle the beta-lactam antibiotics, including penicillins and first-generation cephalosporins, are extensively used for the treatment of mastitis and other diseases. There is also some use of third- and fourth-generation cephalosporins that have been identified as “critically important” for human health (Collignon et al., 2016) and as a risk factor for asymptomatic carriage of ESBL-E in dairy cattle (Gonggrijp et al., 2016). In New Zealand (NZ), beta-lactams have been used prophylactically to prevent mastitis during drying off post-lactation (New Zealand Veterinary Association, 2015).
For intensive high antimicrobial usage (AMU) farming systems such as pig and poultry production, both foodborne (Larsen et al., 2016) and occupational exposure (van Cleef et al., 2015) have been identified as potential transmission pathways for the spread of AMR to humans (Manson et al., 2003; Graveland et al., 2011; Leverstein-van Hall et al., 2011; Agerso and Aarestrup, 2013; Dohmen et al., 2017). However, data on the contribution of dairy farming as a source of antimicrobial-resistant bacteria and genes are less extensive. Dairy production systems provide frequent opportunities for human exposure to bovine microorganisms directly either through ingestion of food or through occupational exposure and indirectly through other pathways, such as environmental contamination.
Dairy farming in NZ is largely a pasture-based system (Holmes et al., 2002) which may contribute to the low AMU compared with other countries (Compton and McDougall, 2013). Data covering 30 countries indicated NZ was the third lowest user of antimicrobials in animal production and used ∼12.9 times less antimicrobials than in human medicine (per mg active ingredient/kg biomass) (Hillerton et al., 2017). As of 2017, NZ had 11,748 dairy herds with an average herd size of 414 cows (NZIER, 2017). Dairy farms are spread throughout NZ; however, the majority is located in the North Island (73%) (NZIER, 2017). NZ dairy farming is also associated with low prevalence of cattle diseases such as coliform mastitis (Lacy-Hulbert et al., 2012) and therefore provides a unique opportunity to study AMR in a pastoral-based dairy farm environment. Such studies also allow comparisons with overseas intensive farm management practices, involving high AMU, increased feed supplementation, and extensive periods of indoor housing.
The aims of this review were twofold: first, we review the current understanding of dairy farming as a potential source for the emergence and transmission of AMR and highlight research gaps using the ESBL-E as an exemplar for antimicrobial-resistant bacteria. Second, we take a NZ perspective on how variation in farm management practices may contribute to AMU and subsequently AMR in dairy cattle.
Bacterial Pathogens of Concern
The WHO has defined a priority list for 20 bacterial pathogens of concern to human health and categorized them as critical, high, and medium, according to 10 criteria relating to health and resistance (Tacconelli et al., 2018). This list includes several pathogens of concern that are also associated with dairy cattle including ESBL-E (discussed in next paragraph), and also fluoroquinolone-resistant Salmonella spp., fluoroquinolone-resistant Campylobacter spp., methicillin-resistant and vancomycin-intermediate and -resistant Staphylococcus aureus (Vandendriessche et al., 2013; Ali et al., 2016; Rodriguez-Rivera et al., 2016).
Enterobacteriaceae
ESBL-E include K. pneumoniae, E. coli, Enterobacter spp., Serratia spp., Proteus spp., Providencia spp., Salmonella spp., and Morganella morganii (Iredell et al., 2016). ESBLs confer resistance to a broad range of beta-lactam antibiotics, including penicillins and third-generation cephalosporins that are commonly used as first-line therapy against Gram-negative bacteria (Johnson et al., 2013; Liebana et al., 2013). Third-generation cephalosporins have been identified as “critically important” antimicrobials for use in human medicine; they are the antibiotics of last resort to treat serious human infections and pathogens causing such infections maybe transmitted through nonhuman sources (Collignon et al., 2009, 2016).
ESBL-E can cause urinary tract and bloodstream infections (Pitout and Laupland, 2008) and are associated with both hospital-acquired (Kaye and Pogue, 2015) and community-acquired infections (Laupland et al., 2008). Recently there has been an increase in the number of community-acquired infections caused by ESBL-E (Toubiana et al., 2016). In NZ, the annualized national incidence of human infections associated with ESBL-E was 95.5 per 100,000 people in 2014, and 49% (176/359) of these infections were community related (Dyet et al., 2014).
Many ESBL-E are resistant to multiple classes of antibiotics (Le et al., 2015; Zhang et al., 2015; Xu et al., 2018). Of particular concern are those that are also resistant to carbapenems. The WHO bacteria priority list used this resistance class as a marker for MDR and pandrug-resistant (resistance to agents in all antimicrobial classes) bacteria (Tacconelli et al., 2018), and carbapenems are often the “last line of defence” for treating serious infections (Collignon et al., 2016).
ESBL-E may cause disease in dairy cattle and have been isolated from milk from cows with mastitis in France (Dahmen et al., 2013), Turkey (Pehlivanoglu et al., 2016), and China (Ali et al., 2016). Although uncommon, ESBL-E abundance may be underestimated if sampling of apparently healthy dairy cattle with subclinical levels of mastitis does not occur (Ali et al., 2016). In NZ, coliform mastitis (caused by members of the Enterobacteriaceae family) is rare (McDougall, 2002; Lacy-Hulbert et al., 2012; McDougall et al., 2018) and ESBL-E prevalence in livestock is unknown (Toombs-Ruane et al., 2017).
Mastitis-Associated pathogens
Numerous bacterial pathogens may cause bovine mastitis and can be categorized into two groups according to their main reservoir: contagious or environmental pathogens (Gomes and Henriques, 2016). Contagious mastitis-causing pathogens generally include S. aureus, Streptococcus agalactiae, and Streptococcus dysgalactiae (Gomes and Henriques, 2016), although Strep. dysgalactiae can also be considered as an environmental pathogen (Oliveira et al., 2013). Environmental pathogens also include members of the Enterobacteriaceae family where E. coli, K. pneumoniae, and Klebsiella oxytoca are of particular importance, as are Streptococcus uberis, coagulase-negative staphylococci (De Vliegher et al., 2012; Bal et al., 2016; Gomes and Henriques, 2016), Serratia spp., Enterococcus spp. (Oliveira et al., 2013), and Pseudomonas aeruginosa (Zadoks et al., 2011). Coliform mastitis is not a common cause of mastitis in NZ (Lacy-Hulbert et al., 2012).
The prevalence and diversity of mastitis-associated pathogens varies between studies, geographic regions, can be herd specific (McDougall et al., 2007; Oliveira et al., 2013; Sztachanska et al., 2016; Gao et al., 2017), and may be because of different methods used for the detection of these pathogens, variation in case definitions of clinical mastitis, and national and local policies regarding AMU in livestock animals and farming practices. The wide range of bacterial pathogens that may trigger bovine mastitis highlights the difficulty in identifying the causative organism(s) and incorrect treatment (either antibiotics to which the etiological agent is resistant to or using third-generation cephalosporins when penicillin would suffice) may contribute to the development and spread of AMR in dairy farm environments.
ESBL-E and mastitis-associated pathogens are both significant bacterial pathogens from dairy farm environments when considering human and animal health and the development and transmission of AMR. We focus hereunder on ESBL-E because of the important public health issue they pose, their indeterminate prevalence in agricultural environments, their detection in the food production chain, and the increasing levels of AMR found in Enterobacteriaceae.
Prevalence of ESBL-Producing Enterobacteriaceae
Prevalence in dairy cattle
Few studies have investigated the prevalence and incidence of ESBL-E in dairy cattle and of those, the prevalence and incidence varies widely (Table 1). Study comparisons however are often confounded because of the study-specific sampling and detection methods used and the sample types, often resulting in data from study populations that cannot be extrapolated (Gonggrijp et al., 2016). For example, culture-based selection methods have been used to determine the prevalence of ESBL-E in several studies (Table 1), but the enrichment protocols used differ, resulting in varying screening sensitivities. Farm selection criteria, smaller sample sizes, or studies investigating antimicrobial-resistant bacteria only from diarrheic samples may also influence the prevalence estimates, leading to estimates that differ from the actual prevalence. In NZ, very few studies have investigated the prevalence of antimicrobial-resistant bacteria, including ESBL-E, in animal-derived food products, dairy cattle, and their environment. One cross-sectional study however found that a low proportion of dairy farms (6%; 1/15) were positive for ESBL-producing E. coli (Aplin et al., 2018; unpublished data).
Prevalence of Extended-Spectrum Beta-Lactamase -Producing Enterobacteriaceae in Dairy Cattle
ESBL, extended-spectrum beta-lactamase.
The prevalence of ESBL-E has been demonstrated to be lower on organic dairy farms than on conventional dairy farms within the same country (Gonggrijp et al., 2016; Santman-Berends et al., 2017). The detection of ESBL-E on organic farms that have reduced AMU compared with conventional farms suggests that factors other than AMU may also contribute to the development of antimicrobial-resistant bacteria (Santman-Berends et al., 2017).
Prevalence in milk and bobby veal
The prevalence of ESBL-E in bulk tank milk and milk from cows with mastitis has not been investigated in NZ, but it varies widely between studies undertaken overseas (0.7–91%; Table 2). The prevalence in milk from cows with mastitis was low (1.5%) in a study in Switzerland (Geser et al., 2012) and high in milk samples (25.53%) from Chinese dairy farms (Ali et al., 2016). The prevalence of ESBL-E in bulk tank milk was relatively low (0–9.5%; Table 2), and although pasteurization of milk using established heat treatment processes will inactivate Enterobacteriaceae (Odenthal et al., 2016), these data indicate that raw milk consumption may pose a public health risk.
Prevalence of Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae in Milk and Veal
Few studies have investigated the prevalence of ESBL-E in veal calves and carcasses. No E. coli isolates (n = 300) from bobby calf carcasses in NZ isolated between 2009 and 2010 were ESBL positive (Heffernan et al., 2011). The detection of antimicrobial-resistant bacteria from veal calf fecal samples and bobby veal in published studies overseas (Table 2) suggest that carcass contamination and transfer to the food chain may represent a public health concern if good hygienic slaughter practices are not adhered to. This food safety concern may be enhanced if nonoptimal transport, cooking, and storage conditions are used.
Prevalence of AMR determinants using Culture-Independent methods
Culture-independent metagenomic methods targeting total DNA from an environmental sample for the detection of antimicrobial-resistant bacteria and their genetic determinants provides a holistic detection approach. Culture-independent methods include quantitative real-time PCR for targeted amplification of AMR genes and shotgun metagenomics involving sequencing of the collective genetic material in an environment (Crofts et al., 2017). Specific AMR gene databases such as the Comprehensive Antibiotic Resistance Database (CARD) (Jia et al., 2017) and ResFinder (Zankari et al., 2012) are available to assign sequencing reads to DNA sequences to infer resistance. Metagenomic methods allow for community analysis to determine the abundance and diversity of microbes present in a sample, unlike culture-based methods that are often limited to specific bacteria according to the isolation methods used.
Culture-independent methods may be limited by the presence of low read numbers of AMR DNA sequences (Pitta et al., 2016) and databases may only classify AMR genes to certain phyla, rather than to the species level (Pitta et al., 2016), although shotgun metagenomics provides improved taxonomical classification at the genus level (Thomas et al., 2017). Finally, although a gene is known to confer resistance to antimicrobials in one organism, it may not be functional in another (Crofts et al., 2017). Thus, the use of both culture-based and metagenomics methods in parallel may provide an improved indication of AMR.
Few studies (Table 3) have investigated the prevalence of beta-lactam resistance genes and the resistome (the entire suite of microbial resistance genes and their precursors in a particular environment [Crofts et al., 2017]) in dairy farm environments using culture-independent methods. Beta-lactam resistance genes have not been commonly detected (Table 3); this may be because of AMR resistance genes being present at low read numbers and/or environmental samples being examined at insufficient DNA sequencing depths. For example, in one study population, <1% of the sequence reads were annotated as AMR genes (Pitta et al., 2016). In another culture-independent study a higher proportion of beta-lactam resistance genes were detected in the feces of dairy cattle treated with a third-generation cephalosporin compared with untreated cattle (Chambers et al., 2015). The number of sequences associated with the transfer of genetic material such as plasmids and bacteriophage was also increased in antibiotic-treated cows (Chambers et al., 2015). This finding suggests AMU may contribute to enhanced levels of horizontal gene transfer and the potential spread of AMR genes in the microbial resistome of the animal host.
Detection of Extended-Spectrum Beta-Lactamase-Producing Antimicrobial Resistance Determinants Using Culture-Independent Methods
AMR, antimicrobial resistance; BLAST, Basic Local Alignment Search Tool.
In several studies (Table 3) the dairy cattle microbial resistome differed between herds; however, animals within the same sample population have been shown to have a resistome with a similar repertoire of AMR gene families (Wichmann et al., 2014). The fecal resistome varied between adult cattle and calves, beef and dairy operations, and calves raised on either conventional or organic dairy farms (Noyes et al., 2016). However, comparisons between culture-independent studies are confounded because of contrasting sample selection, small sample sizes, the analysis pipelines used, and differing AMR gene detection methodologies. No studies have investigated the prevalence of AMR determinants in NZ dairy farm environments and few studies have been undertaken in more intensive farming systems.
AMU on Dairy Farms
There has been a global movement to reduce AMU in livestock animals to prevent the development and transmission of AMR, to improve AMR surveillance, more prudent AMU, and to develop and adopt antimicrobial alternatives (McDougall et al., 2016; Lhermie et al., 2017; Schar et al., 2018). For example, to encourage both prudent use and the reduction of antimicrobials in animal husbandry, the NZ Veterinary Association has aspired that by 2030 NZ will not require antimicrobials for the maintenance of animal health and wellness (Anonymous, 2016b) but rather reserve their use for the treatment of disease. In contrast to swine and poultry, dairy production is not so reliant on antimicrobials. A sample of dairy herds in NZ (n = 477 herds) demonstrated that the antibiotic use rate was generally low compared with international standards, but herd-to-herd variation occurred, with some high-use herds identified (Compton and McDougall, 2013). The ubiquity of a largely pasture-based system may be a contributing factor to the low AMU in NZ dairy cattle.
Dairy farm management practices involving AMU to maintain animal welfare and reduce the prevalence of foodborne pathogens and antimicrobial-resistant bacteria are interrelated (Barkema et al., 2015). Regulations for AMU in dairy cattle vary considerably, with some countries (e.g., United States of America and China) permitting the use of antimicrobials for growth promotion in addition to prophylactic use for the treatment, control, and prevention of animal disease without a veterinary prescription (Maron et al., 2013). In comparison the use of antimicrobials for growth promotion is not permitted in NZ (Anonymous, 1997) and European Union (EU) countries, and their prophylactic use is only permitted in animals with a veterinary prescription (Maron et al., 2013). There is currently a lack of reliable surveillance data on AMU in livestock globally (Van Boeckel et al., 2015) and in many developing countries AMU is unregulated in food-producing animals (Maron et al., 2013).
AMU also differs between conventional and organic farm management systems. In contrast to conventional dairy farms, organic farms must adhere to AMU regulations aimed at improved environmental-based management practices. For example antimicrobial therapy is banned in dairy cattle on United States organic farms; if an animal requires treatment they must be removed from the herd (Barkema et al., 2015; Sorge et al., 2016). In the EU however, organic dairy farms are permitted to use antimicrobials for disease treatment with an increased withholding milk period compared with conventional farms (Barkema et al., 2015). In NZ the regulations are designed to meet export market requirements, therefore organic dairy farms are banned from using antimicrobials (with some specified exceptions) to enable organic milk products to be exported to countries including the United States and the EU members (BioGro, 2009; AsureQuality, 2018). Although the global market for organic milk is increasing, organic dairy production in NZ is in a relatively small scale, with only 2.5% of the total milk sold in NZ in 2015 being organic (Anonymous, 2016a).
The range of antimicrobials approved for veterinary use differs between countries and there is some overlap between those approved for use in both human and animal health. In dairy cattle, antimicrobials are used to treat diseases such as mastitis, endometritis, neonatal diarrhea, and for dry cow therapy (DCT) (Compton and McDougall, 2014). Treatment with DCT (a long-lasting intramammary antibiotic preparation) at the end of lactation is intended to both treat existing mastitis infections and prevent new cases during the dry period. When DCT is applied to the whole herd, it is a farm management practice that may contribute to the development of antimicrobial-resistant bacteria (Kuipers et al., 2016). In NZ, antimicrobial DCT was not economically viable until the introduction of somatic cell count (SCC) penalties (as SCC is an indicator for intramammary infections) in 1993 (Woolford and Lacy-Hulbert, 1996; Lacy-Hulbert et al., 2011).
According to antibiotic sales data from the dairying Waikato region of NZ, 86% of the total antibiotics used (measured as daily doses/animal/year) were for DCT and mastitis treatment (Compton and McDougall, 2014). However, using antibiotic sales data as a proxy for true AMU can be unreliable as farmers may receive bulk antibiotic prescriptions during a season; therefore, it is possible the total antibiotics sold are not always used in one season. In addition, some antibiotics are not species specific; therefore, the sales data cannot be used to infer which animal production system it has been used in. A survey of NZ farmers and veterinarians determined that AMU was influenced by veterinarian advice and personal experience, with farmers generally having limited knowledge of the development and spread of AMR (McDougall et al., 2017). These data suggest mitigation strategies aimed at preventing mastitis and targeted AMU for mastitis treatment may be the most effective measure to reduce AMU on NZ dairy farms.
In the dairy industry alternatives to antimicrobials for the treatment and prevention of mastitis have been recommended including good hygienic practice and teat sealants for the prevention of new mastitis infections during the nonlactation period (Rabiee and Lean, 2013). Whole herd application of teat sealants during the dry-off period has been shown to have a protective effect against the development of ESBL-E-positive herd-level status (Gonggrijp et al., 2016). Other alternatives to antimicrobials for mastitis treatment and prevention (as reviewed by Gomes and Henriques, 2016) include the use of phage therapy and vaccines. These methods are more commonly utilized in the more intensive farming systems of the northern hemisphere where there are higher rates of mastitis. However, these alternatives have limitations because of the complexity and wide range of etiological agents causing mastitis in dairy cattle.
Other Farm Management Practices That May Influence AMR on Dairy Farms
In addition to AMU, a number of other farm management practices are likely to impact the prevalence of foodborne pathogens, antimicrobial-resistant bacteria, and AMR genes in agricultural environments (Barkema et al., 2015). Farm management practices (Table 4), such as AMU, the use of biocides or disinfectants, or the presence of heavy metals, may influence the prevalence of AMR by driving the selection of antibiotic-resistant bacteria or genes, or feed type may influence the prevalence of diseases such as clinical mastitis potentially leading to increased AMU. However these relationships are multifactorial and complex.
Factors That May Influence Antimicrobial Resistance in Agricultural Environments
Housing and feed type are farm management practices that may influence AMR in dairy farm environments, as a higher prevalence of certain pathogens have been associated with these more intensive farming practices. Northern hemisphere dairy farming systems often use indoor barns, such as tie-stall or freestall barns to house cattle during the winter, with limited access to pasture (Saini et al., 2012; Barkema et al., 2015; Sorge et al., 2016). In comparison, dairy farming in NZ uses a majority pasture-based system where cattle are routinely grazed on ryegrass/clover (Holmes et al., 2002); however, there are examples of farms where off-pasture systems and open feed pads are becoming more common.
Intensive farming practices (e.g., supplementary feed and indoor housing) within northern hemisphere farming systems has been associated with an increase in pathogens in livestock including coliform mastitis that may result in increased AMU to prevent and treat such infections (Riekerink et al., 2008; Barkema et al., 2015). Cows that were confined in a more contaminated area were observed to have a higher incidence of coliform mastitis compared with pasture-based cows (Lacy-Hulbert et al., 2002). Although the relationship between cattle housing and AMR has been assessed (Gonggrijp et al., 2016), there has been a lack of correlation between these two factors that is likely because of the complexity and multiple factors involved in the development and transmission of AMR in agricultural environments. Feed regimes may also influence teat infections and consequently the use of antimicrobials to treat such infections; cows fed a total mixed ration (silage and concentrates) diet had a higher incidence of clinical mastitis than all-pasture-based diets (Lacy-Hulbert et al., 2002). In contrast, a metagenomic study of the rumen gut microbiome found no correlation between the prevalence of AMR genes in feedlot steers fed antimicrobial feed additives compared with steers where antimicrobial additives were withheld (Thomas et al., 2017).
Transmission of AMR Enterobacteriaceae in Dairy Farm Environments
The epidemiology of AMR transmission within the dairy farm environment and to humans is complex and several potential pathways for this transmission have been identified (Fig. 1). Theoretical pathways for AMR transmission from dairy cattle and their environment to humans include contact with feces-contaminated environments, through direct contact (e.g., farmers are at high risk during close contact with animals during milking or calving) or through the food chain from animal-derived products including raw milk and veal (Boerlin and Reid-Smith, 2008). However, the actual contribution of each pathway is undetermined.
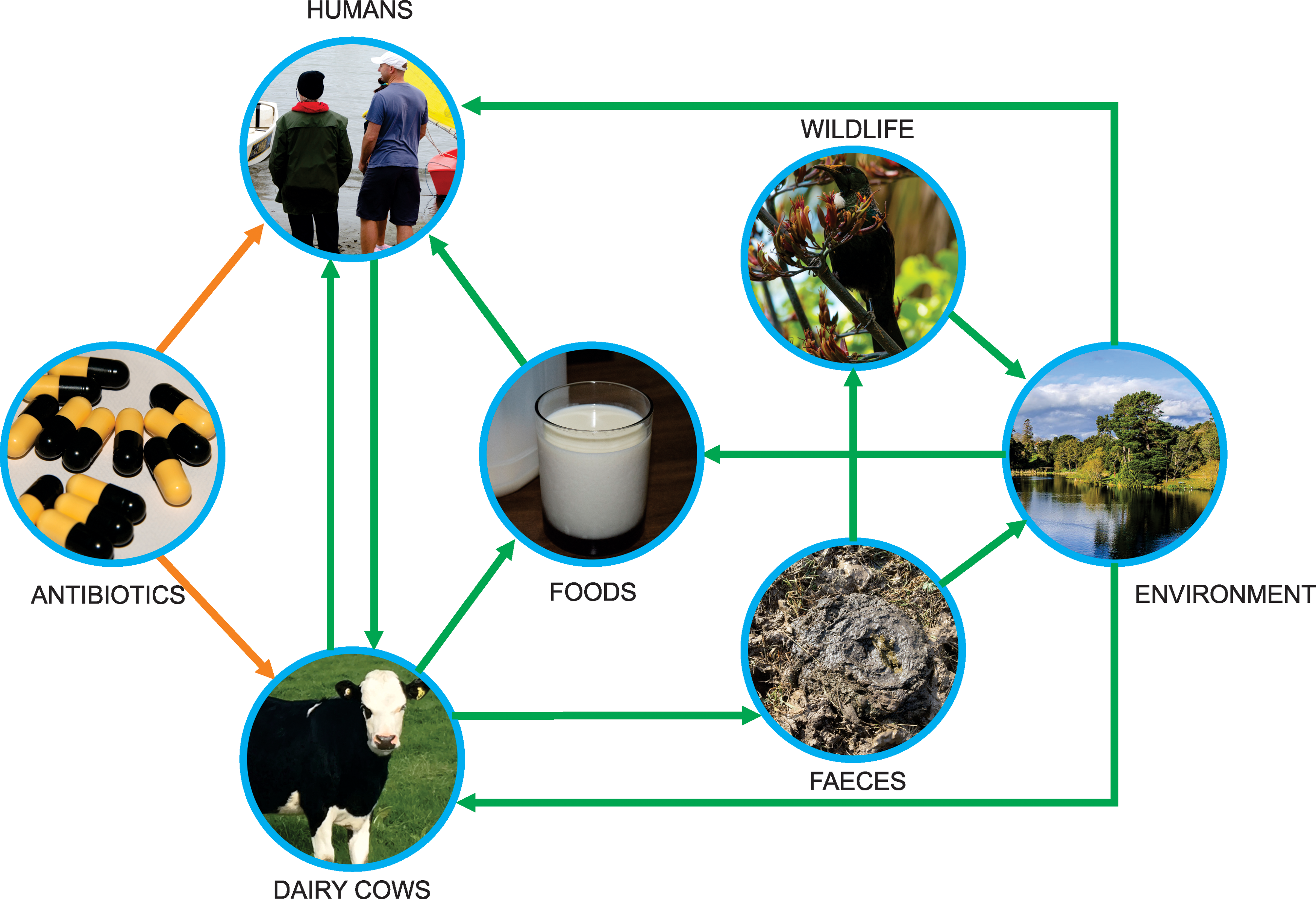
Potential transmission pathways of ESBL-producing Enterobacteriaceae and antimicrobial-resistant bacteria in dairy farm environments. Orange arrows indicate drivers of AMR, green arrows indicate potential transmission pathways, and arrow orientation indicates the direction of the relationship. AMR, antimicrobial resistance; ESBL, extended-spectrum beta-lactamase. Color images available online at
Within agricultural environments, the ruminant digestive tract has been suggested as a reservoir for the development and spread of AMR because of the diverse range of bacterial species and AMR genes present in both the bovine and ovine ruminant gut (Auffret et al., 2017; Hitch et al., 2018). Antimicrobial-resistant bacteria and resistance genes can be excreted in cattle feces (Wichmann et al., 2014; Chambers et al., 2015) that can subsequently contaminate the dairy farm environment, such as fecal contamination of pasture/soil during the application of manure or dairy effluent as a fertilizer to pastures (McKinney et al., 2018).
Evidence for ESBL-E transfer from food-producing animals to humans remains equivocal, with poultry and swine being suggested as more likely sources for the transfer of AMR E. coli compared with cattle (Lazarus et al., 2015), probably because of the routine use of antimicrobials in these production systems. ESBL-E may also be isolated from humans and human sewage (Mesa et al., 2006), indicating a potential for human–human transmission routes.
A German study assessing the risk of animal–human direct contact for the cross-transmission of ESBL-E identified two isolates, one from dairy cattle and the other from a farm worker, shared identical multilocus sequence types (MLST) and CTX-M alleles, potentially indicating zoonotic transfer (Dahms et al., 2015). Whereas a comparison of ESBL- and AmpC-producing E. coli isolated from animal-derived food products, animal gut microbiota, and human clinical cases such as urinary tract infections found little similarity between animal and human isolates at the level of MLST and virulence and AMR gene profiles (Wu et al., 2013), only 1.2% (3 of 258) of the animal isolates were ≥70% similar to the human isolates in relation to MLST clonal complex and microarray gene profiles (Wu et al., 2013). Specific cases were however observed in which animal- and human-derived isolates were similar, with a United Kingdom cattle isolate and Dutch human isolate sharing >90% similarity in virulence and AMR microarray gene profiles, although such similarities only indicate genetic similarity and do not provide sufficient epidemiological evidence supporting zoonotic transfer (Wu et al., 2013).
A systematic review of the role of food-producing animals as a source of human infections caused by expanded-spectrum cephalosporin-resistant extraintestinal E. coli described studies both supporting and rejecting food-producing animals as a source of infection (Lazarus et al., 2015). Where there are associations between resistance in food-producing animals and humans, this seems to be sporadic and limited to specific strains. For example, whole genome sequencing (WGS) identified the recent emergence of a distinct lineage of third-generation cephalosporin-resistant Salmonella enterica serovar Typhimurium in Australia, from both human infections and dairy cattle, suggesting transmission between dairy cattle and humans (Sparham et al., 2017). At present, there are no published studies investigating the transmission and zoonotic transfer of ESBL-E in NZ environments. The evidence for direct contact as a transmission route of antimicrobial-resistant bacteria from cattle to humans is inconsistent partly because of the complexity of the transmission pathways and the methodologies used to infer transmission.
ESBL-E have been detected in numerous animal-derived food products overseas including in bulk tank milk from dairy farms (Sudarwanto et al., 2015; Odenthal et al., 2016; Tepeli and Zorba, 2018), raw milk and cheese during the production process (Tepeli and Zorba, 2018), and veal calves and carcasses (Hordijk et al., 2013b; Haenni et al., 2014; Hutchinson et al., 2017), indicating their occurrence along the food chain. A recent study of veal calves on-farm and pre- and postslaughter isolated AMR E. coli from both farm fecal and preevisceration samples sharing the same pulsed-field gel electrophoresis patterns and resistance phenotypes (Hutchinson et al., 2017), highlighting a potential pathway for the transfer of antimicrobial-resistant bacteria to the human food production chain.
Although numerous potential AMR transmission pathways have been identified, the importance of each of these routes still remains unclear. Variation in the food production chains, farm management practices, and policy regarding AMU in agricultural environments may influence the transmission pathways of antimicrobial-resistant bacteria from animals to humans and vice versa (Lazarus et al., 2015).
Research Needs
ESBL-E pose both a public and animal health concern, and are a potential food safety issue. Dairy farms and the wider farm environment may play an important role in the development and transmission of these pathogens under some conditions; however, these processes are not yet fully understood. This review has highlighted the following research priorities for understanding AMR in the dairy farm environment globally: A major limitation of current studies is the lack of reliable surveillance data on AMU in dairy farm environments (e.g., sales data may be used as a proxy for true AMU). Enhancing surveillance of AMU in livestock would provide an improved understanding of how antimicrobials are used and how this may influence AMR. There is at present a lack of high-resolution transmission and source attribution studies for ESBL-E in dairy farm environments globally. Further research should address the potential transmission of ESBL-E from food-producing animals such as cattle to humans, food products, and the environment using techniques such as WGS to provide high-resolution genomic evidence for the relative contribution of different transmission pathways. AMU and farm management practices likely drive the prevalence and diversity of ESBL-E in agricultural environments and associated food products. Most studies investigating the prevalence of ESBL-E on dairy farms have been conducted in the northern hemisphere and are cross-sectional studies (Table 1). Therefore, additional studies investigating the prevalence of ESBL-E on dairy farms and associated food-products are required in the southern hemisphere, including in NZ. Such studies would provide useful insight into any effects farm management practices (such as those described in Table 4) and seasonal variation have on AMR in agricultural environments. Few longitudinal studies globally, and none in NZ, have been undertaken using a culture-independent metagenomics approach to investigate the abundance and diversity of AMR genes in dairy cattle and the dairy farm environment. Such studies would be beneficial to examine the transmission and development pathways of AMR by providing a high level of detection of AMR genetic determinants in these environments, and this would aid AMR surveillance.
Conclusion
AMU in dairy farm environments and the consequent emergence and transmission of antimicrobial-resistant bacteria, such as ESBL-E, is an increasing concern for both public and animal health and a food safety issue. Several bacterial pathogens of concern to human health, such as ESBL-E, have been detected overseas in the dairy farm environment and associated food products. The prevalence of ESBL-E in cattle, dairy food, and veal products, and the wider farm environment varies between studies and is likely because of contrasting farm management practices, national and local veterinary AMU regulations, food-processing standards, pathogen surveillance, and the sensitivity and specificity of detection methods to identify antimicrobial-resistant bacteria. Therefore, caution should be used when comparing data between studies.
Evidence of transmission of antibiotic-resistant bacteria from food-producing animals to humans and vice versa remains equivocal. This review has highlighted a number of global research needs for AMR studies in dairy farm environments. Further surveillance of AMR in agricultural environments is required to fully understand the mechanisms through which transmission of antimicrobial-resistant bacteria occur in the NZ dairy farm environment and whether ESBL-E, for example, are less prevalent in NZ pasture-based dairy farm systems. Enhanced knowledge of the prevalence, distribution, and transmission of antimicrobial-resistant bacteria in the dairy farm environment is needed to understand the transmission pathways and risk factors for subsequent development of mitigation strategies to reduce AMU and AMR in agricultural environments. Additional high-resolution WGS or metagenomic studies would provide important transmission and prevalence data.
Finally, NZ has policies and guidelines for the judicious use of antimicrobials in animal health and is a comparatively low user of such antimicrobials. However, education, outreach, and stakeholder engagement (by both veterinary professionals and farmers) is required to meet the targets set by the NZ Veterinary Association, which aspire that by 2030 NZ will not require antimicrobials for maintenance of animal health and wellness. Addressing the rising threat of AMR requires a holistic “One Health” approach across multiple sectors and further studies are required to understand the role of the dairy farm environment in the development, persistence, and transmission of AMR bacteria.
Footnotes
Acknowledgments
The authors thank Dr Ahmed Fayaz for his help generating the figure (![]() ) used in this review. This work was supported by the New Zealand Food Safety Science & Research Centre, AgResearch Ltd. (Strategic Science Investment Funding to the Food Provenance and Assurance program) and Massey University. Rose Collis is the recipient of a New Zealand Food Safety Science & Research Centre PhD scholarship. This work was informed by a review of antimicrobial-resistant bacteria in dairy cattle funded by the New Zealand Food Safety Science & Research Centre.
) used in this review. This work was supported by the New Zealand Food Safety Science & Research Centre, AgResearch Ltd. (Strategic Science Investment Funding to the Food Provenance and Assurance program) and Massey University. Rose Collis is the recipient of a New Zealand Food Safety Science & Research Centre PhD scholarship. This work was informed by a review of antimicrobial-resistant bacteria in dairy cattle funded by the New Zealand Food Safety Science & Research Centre.
Disclosure Statement
No competing financial interests exist.