
Abstract
Subclinical Clostridioides difficile colonization in piglets could be a potential source of this bacterium for community-acquired C. difficile infection. The purposes of this study were to assess the effect of specimen type and processing on C. difficile isolation, culture, and detection by polymerase chain reaction (PCR), and to determine the occurrence of C. difficile in piglets of different ages. We compared different culture procedures—direct plating, ethanol shock, and an enrichment step—to isolate C. difficile from swine feces and rectal swabs. DNA was isolated directly from feces, processed feces, and bacterial isolates to detect the triose phosphate isomerase (tpi) gene and identify the toxins A and B genes. The results show that ethanol shock increased the C. difficile isolation from feces, while it decreased it for rectal swabs, in comparison with direct plating. The use of the enrichment broth gave the highest C. difficile recovery from both types of specimen. Our findings show low sensitivity for tpi gene detection after the DNA extraction directly from feces and an increase in PCR-positive samples when feces were processed before the DNA extraction. The overall prevalence of C. difficile was 16.9% (22/130), of which 100% were found to be toxigenic as assessed by the enrichment culture of fecal samples. The rate of isolation of positive samples decreased with the animal age, regardless of the presence or absence of diarrhea. Our results demonstrate the persistent reservoir of toxigenic C. difficile in fecal samples of piglets and support the impact of specimen processing on its isolation.
Introduction
Clostridium difficile, recently renamed Clostridioides difficile (Lawson et al., 2016), is a spore-forming human pathogen associated with serious enteric diseases around the world. C. difficile has been identified also as a cause of enteritis in many animal species, including pigs, and can be isolated from feces, from both diseased and healthy animals (Songer and Anderson, 2006). Thus, subclinical animals can be both hosts of C. difficile and possible carriers and might play an important role in the spread of the community-acquired infection (Hensgens et al., 2012). Furthermore, the presence of C. difficile spores in the feces of animals destined for human consumption may represent a risk for the contamination of meat products, with a possible impact on the economics of food production (EFSA BIOHAZ Panel, 2013).
The data available about C. difficile carriage in animals destined for human consumption in Spain are scarce. Detection of animal C. difficile isolates would help us to further understand the potential changes in the epidemiology of community-associated C. difficile infection in this country.
A multitude of assays are available for C. difficile testing (Arroyo et al., 2005). Traditionally, studies of enteric pathogens are based on stool specimens, although rectal swabs have been proposed for use in surveillance studies of multiple enteric pathogens, due to their easy collection, transport, and handling. The need for specimen pretreatment before culture is based on the fact that the presence of Clostridium spp. is quantitatively lower than that of other microorganisms in most fecal specimens. To facilitate the isolation and detection of C. difficile, methods to eliminate fecal contaminants are used, such as ethanol shock, based on the high survival capacity of the spores of C. difficile in ethanol, set against the inactivation of the vegetative forms of the contaminants present in the specimen (Marler et al., 1992). Molecular detection methods could potentially serve as a more rapid screening test, compared with conventional culturing techniques (Bélanger et al., 2003; van den Berg et al., 2006, 2007). However, many commercial assays for the detection of C. difficile have been systematically evaluated for use in human samples, but their performance with specimens of animal origin or other complex samples (soil, water, food, etc.) has not been validated.
The first objective of this study was to develop a specific and sensitive method for C. difficile isolation from fecal specimens of piglets and to assess if the C. difficile recoveries from feces and rectal swabs were comparable. The second objective was to evaluate the direct extraction of DNA from feces and processed feces for the detection of C. difficile by polymerase chain reaction (PCR). The third objective was to determine the occurrence and the toxigenic profile of C. difficile in different age groups of piglets from Murcia, Spain.
Materials and Methods
Animals and sampling
One hundred thirty samples of feces were obtained from 109 piglets belonging to 4 age-range populations (1–15 days, 16 days to 1 month, 1–2 months, and 2–3 months). Sampling included 114 feces from 93 piglets from “farm A” and 16 from 16 piglets from “farm B,” both located in Murcia (in the southeast of Spain), between February and October 2015.
All fecal specimens were collected directly from the animals in sterile plastic containers after rectal stimulation with a dry cotton-tipped swab (Deltalab).
The samples were preserved in sterile packs at −20°C until being processed, except for an aliquot of each feces sample that was used for procedure 3 (explained below); this was processed within 2 h of its collection.
Specimen pretreatment and C. difficile cultivation
Eighteen feces samples, previously known to be culture positive for C. difficile and their corresponding rectal swabs were selected for the first objective. As a preparation step for the specimens before culture, we tested an ethanol shock and an enrichment step and compared both with the direct plating.
Procedure 1: “direct plating”
A feces aliquot of ∼0.5 g was directly plated onto C. difficile selective agar “CLO” (bioMèrieux, Marcy l'Etoile, France). Each rectal swab tip was briefly scrubbed out onto solid CLO agar.
Procedure 2: “ethanol shock” before plating
A feces aliquot of ∼1 g was mixed with an equal volume of filter-sterilized 70% ethanol (v/v; Panreac Química S.L.U., Barcelona, Spain). Each rectal swab used in procedure 1 was submerged for 30 s in a microtube containing 1 mL of 70% ethanol. Both kinds of specimen-ethanol mixture were vortexed and incubated at room temperature (23–25°C) for 20 min. Then, each mixture was centrifuged at 4000 rpm for 10 min, and 75 μL of the pellet, resuspended with the remaining ethanol, and were plated onto CLO agar.
Procedure 3: “enrichment step+ethanol shock” before plating
A feces aliquot of ∼1 g was inoculated into 5 mL of C. difficile enrichment broth medium described by Blanco et al. (2013). Each rectal swab used in procedure 2 was also used in procedure 3 and was introduced into 5 mL of C. difficile enrichment broth. The inoculated broths were incubated anaerobically—80% N2, 10% CO2, and 10% H2—in an anaerobic chamber (bioMèrieux) at 37°C for 7 days. Then, 2 mL of each broth was mixed with an equal volume of filter-sterilized absolute ethanol (Panreac Química S.L.U.), vortexed briefly and incubated at room temperature for 1 h. Then, each mixture was centrifuged at 4000 rpm for 10 min, and 75 μL of the pellet, resuspended with the remaining broth, and were plated onto CLO agar.
Figure 1 shows a schematic representation of the processing of the specimens and the culture of the feces and rectal swabs. The culture plating onto CLO agar was performed by four-way streaking, differentiating four areas: discharge area, first quadrant, second quadrant, and third quadrant. The plates were incubated anaerobically at 37°C for 72 h. The isolates were identified as C. difficile on the basis of morphological criteria (grayish, nonhemolytic, and rough colonies) and the typical horse-manure-like odor. They were subcultured onto brain heart infusion broth (BHI) agar plates (Scharlau; Scharlab, S.L.) and incubated anaerobically at 37°C for 24–48 h. The area where there was growth of suspected C. difficile colonies and other fecal microbiota, able to survive the ethanol and antibiotics used in the processing, was recorded.
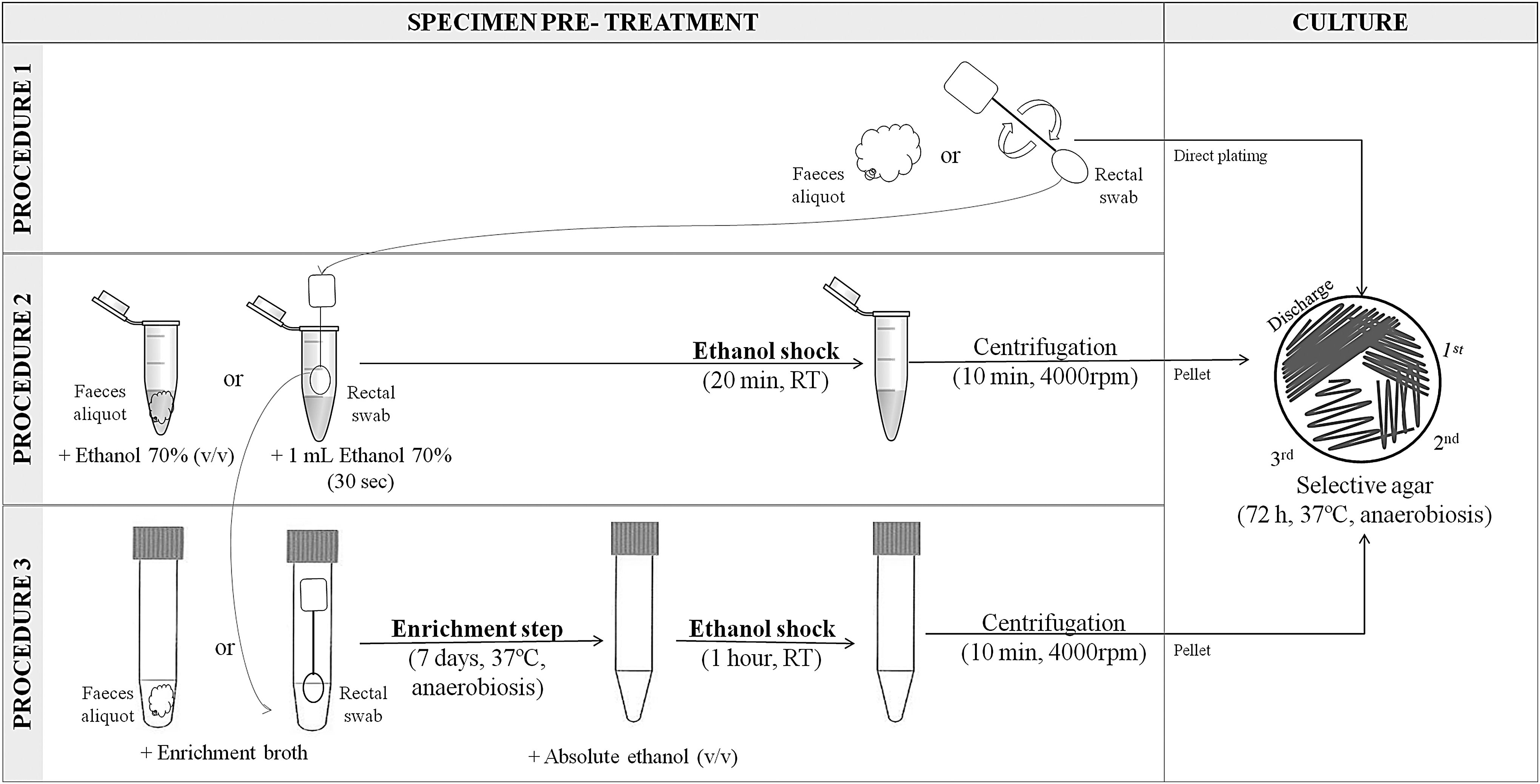
Schematic representation of specimen pretreatment and culture of feces and rectal swabs. A detailed description of procedures 1, 2, and 3 and the culture is given in the Materials and Methods section. RT, room temperature (23–25°C)
DNA extraction from bacterial isolates, feces, and enriched feces
Figure 2 shows a schematic representation of DNA extraction from bacterial isolates, feces, and enriched feces. The DNA from C. difficile isolates was purified from suspected C. difficile colonies grown on BHI agar by a manual method described by Lemee et al. (2004). In brief, bacterial suspensions, made using one colony and 0.5 mL of distilled water, were heated at 100°C for 20 min and then centrifuged for 2 min at 10,000 rpm to pellet bacterial debris. The supernatants were used as a source of DNA for PCR. This manual method was set to compare with a commercial kit designed for removing Taq polymerase inhibitors of complex sample matrices. The commercial kit -QIAamp DNA stool Mini Kit (Qiagen) was used for DNA extraction of 18 aliquots of feces (0.2 g) and 22 enriched—with C. difficile enrichment broth—feces aliquots (0.2 mL), according to the manufacturer's protocol. The DNA was quantified in both methods by absorbance spectrophotometry (NanoDrop-2000 Spectrophotometer; Thermo-Scientific, NanoDrop products, Wilmington, DE).
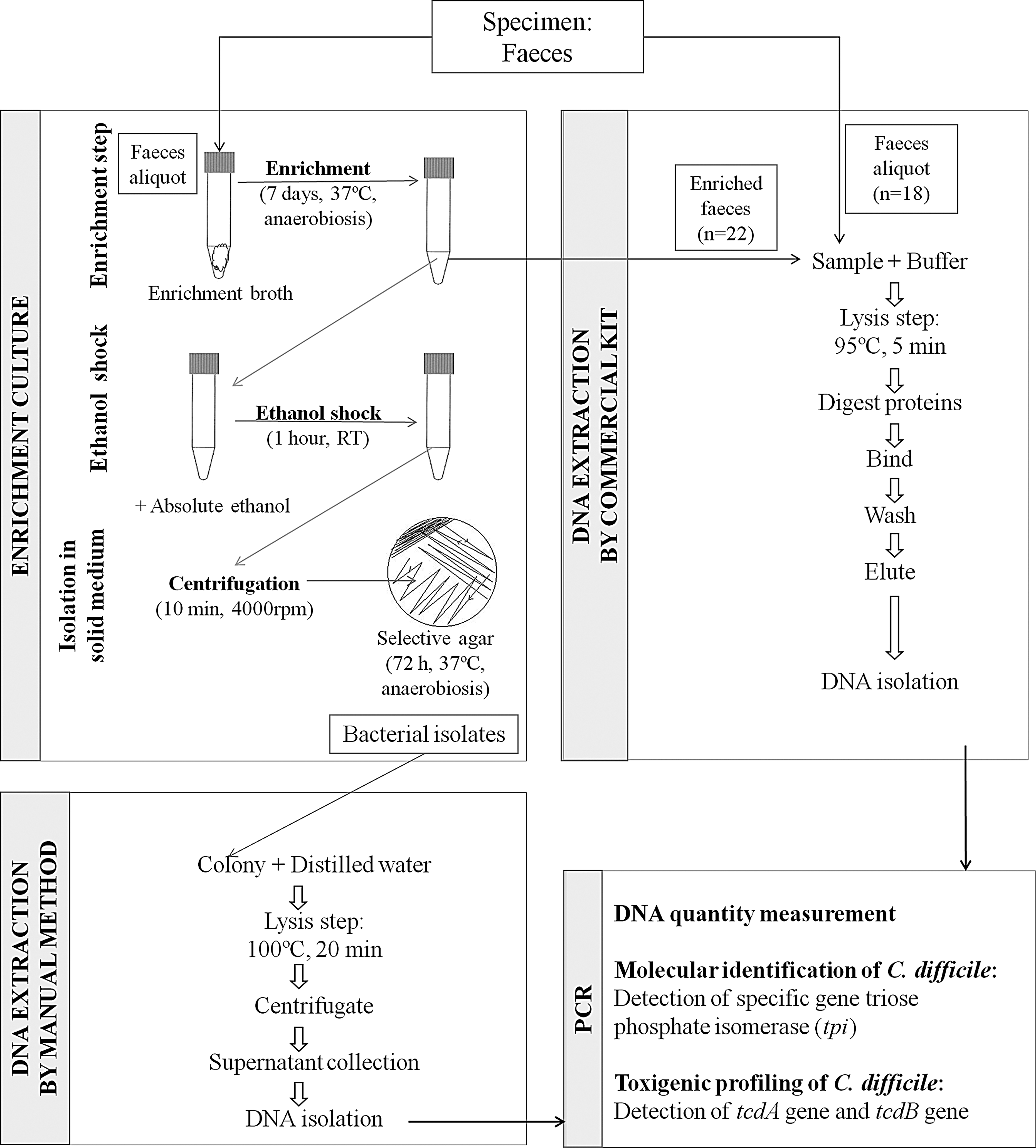
Schematic representation of DNA extraction from bacterial isolates, feces, and enriched feces.
Molecular identification and toxigenic profiling of DNA isolates
All the DNA extracts were evaluated by real-time PCR for the presence of the triose phosphate isomerase (tpi) housekeeping gene, an alternative marker to 16S ribosomal DNA for genotypic and phylogenetic characterization of bacterial species (Verlag et al., 2003). This gene allows the differentiation of pathogenic Clostridium species by revealing some variable gene regions. Positive tpi gene isolates were screened by PCR for the tcdA and tcdB genes, encoding toxin A and toxin B, respectively. The sequences of the amplification primers are listed in Table 1. The PCRs were performed in a CFX96 thermal cycler (Bio-Rad), in a final volume of 25 μL containing 0.5 μL of the forward primer (10 μM), 0.5 μL of the reverse primer (10 μM), 1 μL of DNA template, 10.5 μL of PCR-grade water (Panreac Química S.L.U.), and 12.5 μL of SensiMix SYBR No-ROX kit (Bioline). Detection of tpi and tcdB were done using the primers and thermal cycling conditions as described by Lemee et al. (2004). Detection of the nonrepeating region of tcdA gene was done using the NK2 and NK3 primers as described by Kato et al. (1991) and thermal cycling conditions as described by Justin and Antony (2015). DNA manually extracted from an isolate of C. difficile ATCC 9689 (Colección Española de Cultivos Tipo, Universitat de València, Spain)—a strain positive for toxins A and B—and PCR-grade water were used as positive and negative controls, respectively. Finally, melting curve analysis was performed to determine the specificity of the amplified products.
Sequences of the Specific Primers Used in Polymerase Chain Reaction Amplification
All primers were purchased from Sigma-Aldrich (St Louis, MO).
Nonrepeating region.
tpi, triose phosphate isomerase.
Results
The first objective of this study was to compare the performance of three culture procedures—direct plating, ethanol shock before plating, and an enrichment step before ethanol shock—used to isolate C. difficile from piglet specimens and to assess if the C. difficile recovery rates from feces and rectal swab samples were comparable. C. difficile was recovered from 66.67% (12/18) of feces and 94.44% (17/18) of rectal swabs with the direct plating procedure (procedure 1). For feces, the rate of recovery was increased when specimen pretreatment was used, obtaining isolation rates of 94.44% (17/18) with ethanol shock (procedure 2) and 100% (18/18) with the enrichment step (procedure 3). By contrast, for rectal swabs, the ethanol shock decreased the recovery rate to 83.33% (15/18), and the enrichment step increased the isolation rate to 100% (18/18).
Table 2 shows the growth area results of pure and contaminated C. difficile-positive cultures from feces and the corresponding rectal swab, according to the pretreatment. In the direct plating procedure, C. difficile was isolated from nonpure cultures contaminated with fecal microbiota in 91.67% of cases (11/12) for feces and in 88.23% of cases (15/17) for rectal swabs. This contamination rate decreased when specimen pretreatment was used, especially with ethanol shock, obtaining contamination rates of 47.06% (8/17) in feces and 46.67% (7/15) in rectal swabs, while the contamination rate in the enrichment treatment was 72.22% (13/18) in feces and 83.33% (15/18) in rectal swabs. Regarding the growth area, in most cases C. difficile reached the third quadrant in the selective medium, mainly in the feces samples.
Growth Area Results of Pure and Contaminated Clostridioides Difficile-Positive Cultures from Feces and the Corresponding Rectal Swab, According to the Pretreatment
C. difficile pure culture.
+, first quadrant; ++, second quadrant; +++, third quadrant; d, discharged area; L, liquid; NG, no growth; S, solid/semisolid.
The second objective of this study was to assess the performance of DNA extraction—directly from feces, from enriched feces, and from bacterial isolates—for the detection of C. difficile by PCR. A mismatch between the PCR results for bacterial isolates and those of feces was found. Positive PCR results obtained from bacterial isolates of a selection of feces were taken into account to determine true positive samples. On this basis, the specific C. difficile tpi gene was amplified in 11.11% (2/18) of samples when the DNA used in the PCR was extracted directly from feces. When DNA extract from a fecal aliquot was enriched with C. difficile enrichment broth, C. difficile was detected in 27.27% (6/22) of samples.
The third objective of this study was to determine the occurrence of toxigenic C. difficile in different age groups of piglets. Table 3 shows the occurrence of C. difficile-positive feces cultures according to the age population and feces consistency. Of the 130 feces samples processed with the enrichment culture (procedure 3), bacterial growth was obtained in 111 samples, of which 31 were suspected C. difficile colonies. The presence of the tpi gene was confirmed in 16.9% (22/130) of samples, all of them from the same farm. All the positive samples belonged to different animals. Both the tcdA gene and the tcdB gene were amplified by PCR (A+B+) in 100% (22/22) of C. difficile isolates.
Occurrence of Clostridioides Difficile-Positive Feces Cultures, According to the Pig Age Population and Feces Consistency
Discussion
In the present study, the ethanol shock treatment of specimens before plating decreased the growth of fecal bacteria, favoring the identification of C. difficile and accelerating its isolation. The incubation of specimens in an enrichment broth supplemented with lysozyme and compounds such as bile salts improves C. difficile recovery, facilitating the germination of its spores on a solid medium (Buggy et al., 1985). We obtained the 100% C. difficile recovery both in feces and swabs with the enrichment incubation before plating, in agreement with other authors (Arroyo et al., 2005; Blanco et al., 2013). In this study, the enrichment treatment of rectal swabs gave sensitivity results identical to those of feces, in agreement with other authors (McFarland et al., 1987; Curry et al., 2011; Kundrapu et al., 2012; Shakir et al., 2012). Unlike the feces culture, direct plating of swabs yielded better recovery rates than the ethanol shock procedure. This may be due to the fact that the same swab was used for procedures 1 and 2, being the first step in the direct plating. During the rolling of the swab tip over the culture medium surface, the microbial load may have been reduced for the next step. If two different swabs were used, further studies would be necessary to conclude whether a causal association existed between the order of swab collection and the order of the pretreatment used.
Although PCR could serve as a rapid screening test, the literature on this topic is limited to a few studies, which reported varied performance with respect to the detection of C. difficile directly in feces of animal origin (Alvarez-Pérez et al., 2009; Avbersek et al., 2011, 2017). Our findings show low sensitivity for tpi gene detection after DNA extraction directly from piglet feces, using the commercial kit, compared with the manual method for extraction from bacterial isolates. The most critical step in PCR assays is the processing of the specimens. Although it is not essential to isolate DNA of great purity, it is necessary to remove or inactivate substances that may inhibit PCR assays and that are common to complex samples, such as feces. The enrichment of samples before PCR analysis, consisting of sample incubation in a selective broth for 5–7 days, and subsequent DNA extraction from these broths, has been shown to be effective in the detection of DNA in complex samples (Tansuphasiri et al., 2005; Houser et al., 2010). In accordance with this, the data presented in this study show a 16.16% increase in PCR-positive samples when the feces were enriched before DNA extraction, compared with nonenriched feces. A likely explanation for this result is that this enrichment procedure includes the multiplication of C. difficile in a broth to a PCR-detectable concentration. Despite the enrichment step used in this study, 16 samples were positive for C. difficile culture and negative for tpi gene detection. Although the reason for this observation remains unclear, we speculate that these false-negative results are due to the duration of the lysis step used in DNA extraction and the difficulty in achieving lysis of the spores. As C. difficile is sporogenic, its resistant spore coat is difficult to lyse; therefore, the release of nucleic acids is limited and this reduces the effectiveness of C. difficile DNA extraction methods (Avbersek et al., 2017). These sensitivity analysis results agree with the Society for Healthcare Epidemiology of America and the Infectious Diseases Society of America, which recommend feces culture as the most sensitive test (Cohen et al., 2010). Although traditional culture is time-consuming and laborious, it provides an isolate for further genotyping and susceptibility testing, essential for epidemiological studies (Cohen et al., 2010).
Previous European reports showed that the mean carriage rate of C. difficile in symptomatic and subclinical piglets was 24.3%, with the highest rate reported in Sweden (67%) (Rodriguez et al., 2016). Our findings show an overall prevalence of C. difficile of 16.9%. This is lower than that reported in other European studies, but comparable to previous results from other farms in Spain, which showed an isolation rate of 17.9% for the studied population (Alvarez-Pérez et al., 2009). Worldwide, the overall prevalence ranges from 0% to 73%, with the highest rate reported in Ohio (Rodriguez et al., 2016). All the 22 isolates found in this study were toxigenic. This was in agreement with the isolation rates obtained in previous reports that toxin genes were found at 83.6–100% of isolates (Rodriguez et al., 2016).
Hopman et al. (2011) demonstrated a cumulative prevalence of 100% within 48 h after the birth of piglets. This intestinal colonization in new-born piglets occurs during the first hours of life, although clinical manifestations only occur in specific circumstances (Hopman et al., 2011). An important age effect was observed in the C. difficile colonization and several studies reported a significant decrease in isolation rates over time for piglets after the first week of life (Alvarez-Pérez et al., 2009; Weese, 2009; Rodriguez et al., 2012). Our results indicate a decrease in colonization rates to below 10% after the first month of life. Among the population groups analyzed in the present study, carriage rates ranged from 7.69% of 2- to 3-month-old pigs to 32.56% of piglets <15 days old. A likely explanation for this age effect is that the bacterium is better able to colonize and proliferate in the intestinal tract of younger animals, where the gut microbiota is less developed.
Conclusion
This study demonstrates that subclinical colonization in piglets could be a significant reservoir of C. difficile and a potential source of bacteria for community-acquired C. difficile disease. Currently, no guidelines are available for the detection of C. difficile in animals. Our results support the use of an enrichment broth supplemented with sodium taurocholate before plating on a solid medium; therefore, this step is recommended to maximize the C. difficile recovery. The establishment of a standardized method could be useful in the implementation of measures to prevent C. difficile transmission to farm personnel or its entry into the food chain by contamination of carcasses at slaughterhouses.
Ethics Statement
The study received approval from the University Ethics Committee for Animal Experimentation and the Autonomic Community of Murcia Region Ethics Committee (Study protocol No. A13150101). The study was carried out in accordance with EU Directive 2010/63/EU for the care and use of animals for scientific purposes.
Footnotes
Acknowledgment
This work was supported by the University of Murcia and the Regional Campus of International Excellence Campus Mare Nostrum.
Disclosure Statement
No competing financial interests exist.