
Abstract
A study using sentinel broiler chickens was performed to address Campylobacter persistence in litter that was reused for successive flocks. Cloacal swabs, litter, drag swabs, darkling beetles, feed, and drinking water were weekly sampled and analyzed by standard microbiological procedures. Thermotolerant Campylobacter isolated strains were confirmed by polymerase chain reaction and subtyped by pulsed-field gel electrophoresis analysis. Campylobacter was not detected in samples collected immediately after downtime between broiler flocks. However, Campylobacter-positive samples were first detected at 21 d. After Campylobacter was initially isolated from the cloacal swabs, reused litter, drag swabs, or darkling beetles, these samples remained Campylobacter positive in the following weeks until the end of the rearing period. Campylobacter-positive cloacal swabs obtained from sentinel broilers ranged from 97.3% to 100% at 42 d. All isolated strains were identified as Campylobacter jejuni. Among the subtypes identified, an indistinguishable C. jejuni strain was predominant in sentinel broilers and was also detected in the other environmental samples analyzed, suggesting a common and persistent contamination source within the flocks. Sentinel broilers may have contributed to amplify the Campylobacter level, maintaining flock and broiler house contamination until the end of the production cycle.
Introduction
A variety of foodstuffs can be contaminated by Campylobacter; however, chicken meat is considered a main source of human campylobacteriosis (Hansson et al., 2018; Sibanda et al., 2018). Broilers can carry high numbers of thermotolerant Campylobacter species in their guts, the content of which might contaminate carcasses at slaughter despite efforts in processing to prevent contamination (Hutchison et al., 2017). Therefore, further reduction of Campylobacter depends on decreasing the levels of contamination in primary broiler production or processing (Battersby et al., 2016).
Campylobacter colonization of broilers is associated with particular husbandry practices (Bull et al., 2006). In Brazil, litter is composted to reduce microbial loads during downtime and is typically reused for multiple batches of broilers before being definitively removed from the broiler house (Vaz et al., 2017). Litter reuse avoids the disposal of large quantities of waste; however, there is concern about the carryover of pathogens in the broiler house (Smith et al., 2016; Kassem et al., 2017). Although Campylobacter remains viable in artificially contaminated litter (Brooks et al., 2015; Sandilands et al., 2018), it is believed that strains do not survive in dry litter in field conditions (Thakur et al., 2013). Nevertheless, Campylobacter-positive litter has been detected at the end of the rearing period on broiler farms (Bull et al., 2006).
It is noteworthy that stressor effect on Campylobacter strains might trigger the viable but nonculturable (VBNC) state (Newell and Fearnley, 2003; Kassem et al., 2010). Such VBNC cells lose their ability to grow on culture media but allow their survival in litter for a long time (Smith et al., 2016; Hansson et al., 2018), which might underestimate the detection of Campylobacter and represent a potential microbiological risk for the successive flock (Kassem et al., 2017). Stressed Campylobacter cells might multiply after colonization of a host (Newell et al., 2011; Sandilands et al., 2018; Sibanda et al., 2018), allowing more appropriate conditions for detection. Therefore, a study using sentinel broilers reared on composted litter was performed in a controlled broiler farm setting to address Campylobacter persistence in reused litter. Darkling beetles (Alphitobius diaperinus), drinking water, and feed were also sampled to analyze their relationship with Campylobacter retrieved from broilers and reused litter.
Materials and Methods
Broilers and broiler farm
Three successive sentinel broiler flocks were raised in the all-in/all-out system in a multi-pen broiler house at a broiler research farm in southern Brazil. Each flock comprised 180 one-day-old male broiler chicks distributed in six pens (11 broilers/m2) built with a concrete floor. Cobb 500 broiler chicks came from a commercial hatchery, were fed with a standard corn–soybean meal-based diet supplemented with 8.8 g per ton of tylosin phosphate, and received drinking water from a chlorinated supply. Litter consisted of pine wood shavings previously used to raise three clinically healthy broiler flocks. Just after depopulation at 42 d, superficial feathers were burned and caked litter was removed. The remaining litter was windrowed at the center of each given pen for 10 d and redistributed to a height of 10 cm for 3 d before rearing the successive flock. A layer containing 10% of fresh litter was replaced below the brooding chamber in each pen. To access the broiler house, staff took showers and dressed in dedicated boots and overalls. New clean disposable shoe covers were changed when entering each pen. Broilers were cared for according to the animal welfare and ethical standards set forth in the protocol approved by the Ethics Committee for Animal Experimentation (CEUA, No. 007/09).
Sampling
Litter, drag swabs, and darkling beetle samples were collected from pens before the placement of each sentinel broiler flock. Pooled samples of paper lining placed in a sterile plastic bag and swabs of internal surfaces placed in 150 mL of 1% buffered peptone water (BPW) were collected from day-old chick delivery crates. Next, flocks were sampled on days 7, 14, 21, 28, 35, and 42 when cloacal swabs, litter, drag swabs, darkling beetles, feed, and nipple water were collected. Litter sample (∼500 g) consisted of a pooled sample collected from each pen. Drag swab was collected from litter surface in each pen and placed as a pooled sample (six swabs) in 150 mL of 1% BPW. Pooled feed (∼500 g) and drinking water (∼500 mL) samples were collected from feed hoppers and nipples on the drinking lines in each pen, respectively. Darkling beetles were randomly collected inside the pens. A pooled cloacal swab was created by collecting swabs from each of three broilers, which were placed in tubes containing Cary-Blair (Oxoid, Hampshire, United Kingdom).
Thermotolerant Campylobacter isolation
Litter (10 g), feed (10 g), and drinking water (10 mL) samples were suspended in 90 mL of Bolton broth (Oxoid). Swabs of crates and drag swabs suspended in 1% BPW were mixed thoroughly, and 10 mL was inoculated in 90 mL of Bolton broth. Paper lining and macerated darkling beetles were suspended in 1% BPW at a ratio of 1:4 (mass/volume) following inoculation into nine times its volume of Bolton broth. Pooled cloacal swabs were inoculated in 15 mL of Bolton broth. Samples were incubated for 24 h (±4 h) at 41.5°C under a micro-aerobic atmosphere (5% O2, 10% CO2, and 85% N2) before being plated onto modified charcoal cefoperazone deoxycholate agar (mCCDA; Oxoid) and Preston agar (Oxoid) for 24–48 h at 41.5°C in micro-aerobic conditions. With the exception of the cloacal swabs, the samples were also directly streaked onto mCCDA and Preston agar following incubation for 24–48 h. Colonies exhibiting Gram-negative curved rods were confirmed as Campylobacter as described (Vaz et al., 2014).
Polymerase chain reaction
A subset of 96 Campylobacter strains isolated from broilers and environmental samples from the first (n = 33), second (n = 30), and third (n = 33) flocks at each sampling time were tested by polymerase chain reaction (PCR). Genomic DNA was extracted from Campylobacter cells according to Boom et al. (1990). PCR was based on a 287 bp sequence of the 16S rRNA gene from Campylobacter jejuni, Campylobacter coli, and Campylobacter lari (Josefsen et al., 2004). PCR mixture contained 1 × GoTaq Flexi buffer (Promega, Wisconsin), 2.25 mM MgCl2, 200 mM of each dNTP (Promega), 12 pM of each forward (5′-CTGCTTAACACAAGTTGAGTAGG-3′) and reverse (5′-TTCCTTAGGTACCGTCAGAA-3′) primer (IDT, Iowa), 1 U of GoTaq DNA polymerase (Promega), 2 μL of DNA, and water in a total volume of 25 μL. Reaction was amplified after denaturation at 94°C for 2 min followed by 35 cycles of 94°C for 30 s, 60°C for 15 s, and 72°C for 30 s. C. jejuni (ATCC 33560) was used as a positive control, whereas Campylobacter fetus (ATCC 19438) served as negative control.
DNA macrorestriction analysis
Campylobacter strains analyzed by PCR were subtyped by pulsed-field gel electrophoresis (PFGE) as described (Ribot et al., 2001). Briefly, DNA digested with SmaI (New England Biolabs, Hertfordshire, United Kingdom) was electrophoresed with an initial switch time of 6.75 s and a final switch time of 38.35 s in a CHEF Mapper XA system (Bio-Rad, California) for 22.5 h at 6.6 V/cm and an angle of 120°. DNA macrorestriction patterns were analyzed with the BioNumerics software version 6.1 (Applied Maths, East Flanders, Belgium). Similarity was calculated by the Dice coefficient with a position tolerance of 1.5%. A dendrogram was generated by cluster analysis using unweighted-pair group method with arithmetic mean (UPGMA).
Statistical analysis
Analysis of the percentage of Campylobacter-positive cloacal swabs was performed through logistic regression using the LOGISTIC procedure of Statistical Analysis System (SAS Institute, Inc., 2012, North Carolina) considering flock and broiler age effects.
Results
Campylobacter in broilers
Samples collected from the day-old-chick delivery crates tested negative for Campylobacter. Nevertheless, Campylobacter-positive broilers were detected during the rearing period and persisted up to depopulation at 42 d when positive cloacal swabs obtained from each broiler flock sampled ranged from 97.3% to 100% (Table 1). The total number of pooled cloacal swabs taken in the first week (14 swabs) was established to detect at least one Campylobacter-positive sample, considering an expected prevalence of 20%, with a 90% sensitivity of the diagnostic method and 95% confidence level (Cannon, 2001). After the first positive-pooled cloacal swab detection, intra-flock sampling was estimated considering a 30% expected prevalence at 28 d (35 pooled cloacal swabs) and 50% at 35 and 42 d (37 pooled cloacal swabs), with 95% confidence level and 10% accuracy (Thrusfield, 2010). Differences (p < 0.0001) were found among the sentinel flocks; nevertheless, no difference (p > 0.05) was detected between the first and the third flock. However, sentinel broilers from the second flock were detected Campylobacter positive earlier than the other flocks (Fig. 1). Based on the estimates of the model parameters, the broiler age to detect 50% and 95% of the Campylobacter-positive cloacal swabs was determined according to Agresti (2002). Fifty percent of the Campylobacter-positive cloacal swabs were detected at 17.9 d in the second flock and 28.4 in the first and third flocks, whereas 95% of the Campylobacter-positive cloacal swabs were detected at 24.3 d in the second flock and 34.8 d in the first and third flocks.
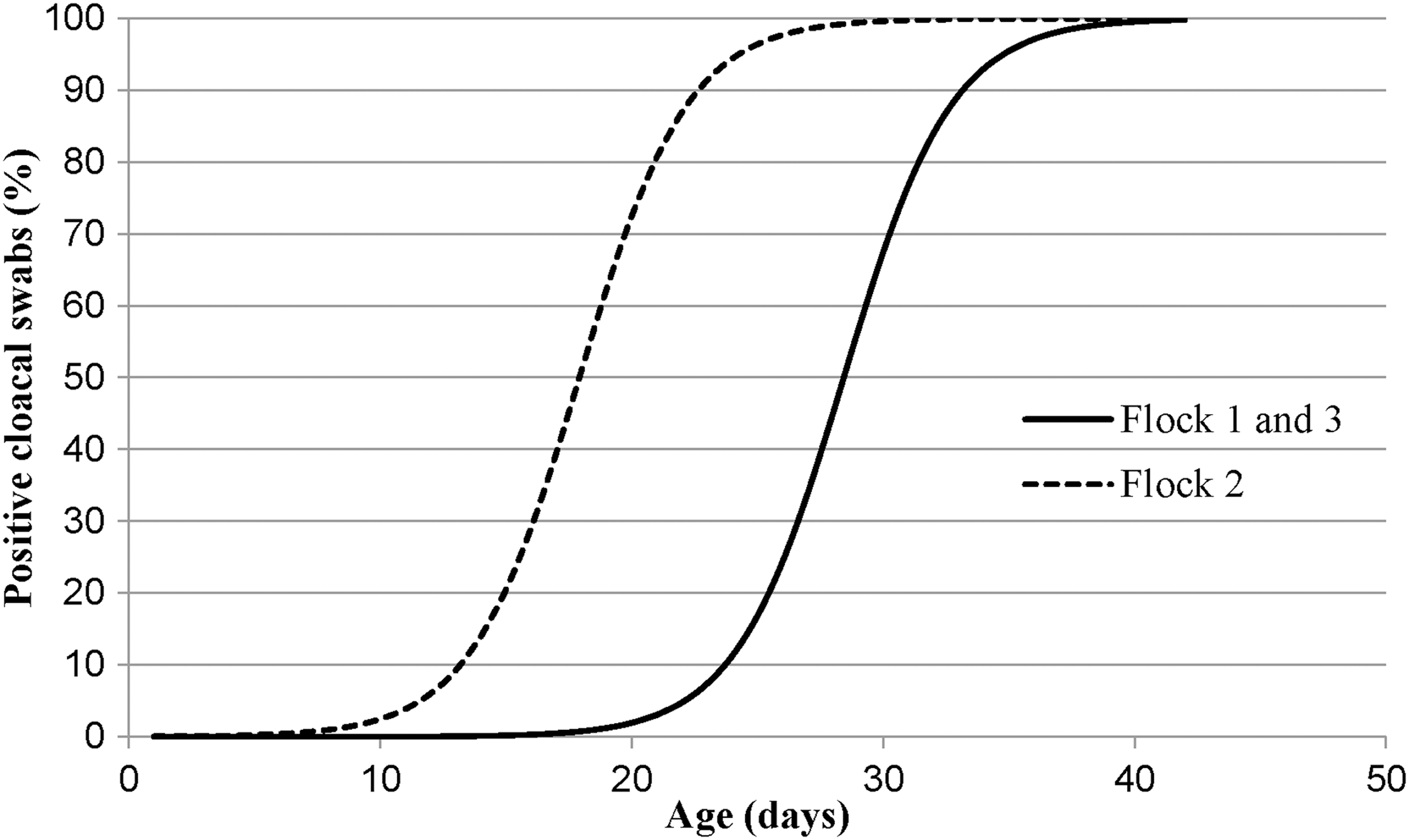
Age of broilers for detection of Campylobacter-positive cloacal swabs in each sentinel flock.
Distribution of the Campylobacter jejuni-Positive Samples in Sentinel Broiler Flocks
Bold values represent Campylobacter-positive results.
Percentage of positive pooled cloacal swabs.
Positive or negative pooled sample.
All samples collected on days 7, 14, and 21 tested Campylobacter-negative.
All samples collected on days 7 and 14 tested Campylobacter-negative.
Campylobacter in environmental samples
Environmental samples collected before the placement of each sentinel broiler flock tested Campylobacter negative but became Campylobacter positive over the course of the rearing period (Table 1). Campylobacter was isolated from water collected from nipple drinkers in the second broiler flock (Table 1), whereas all feed samples tested Campylobacter negative.
Campylobacter strains
Strains isolated from sentinel flocks were phenotypically characterized as C. jejuni. PCR analyses confirmed all tested isolates as thermotolerant Campylobacter.
PFGE typing
Four PFGE subtypes (A to D) were distributed among the C. jejuni strains analyzed (Fig. 2). The majority of the strains (n = 92) shared a predominant profile (A) and were isolated from samples taken from flocks at different sampling times. This subtype showed 87.5% similarity to the profile (B) found in a strain isolated from a litter sample taken from the second flock. The remaining subtypes were found in strains isolated from the first flock (C) and from the third flock (D), with similarity levels below 60% in relation to the predominant profile found in C. jejuni strains.
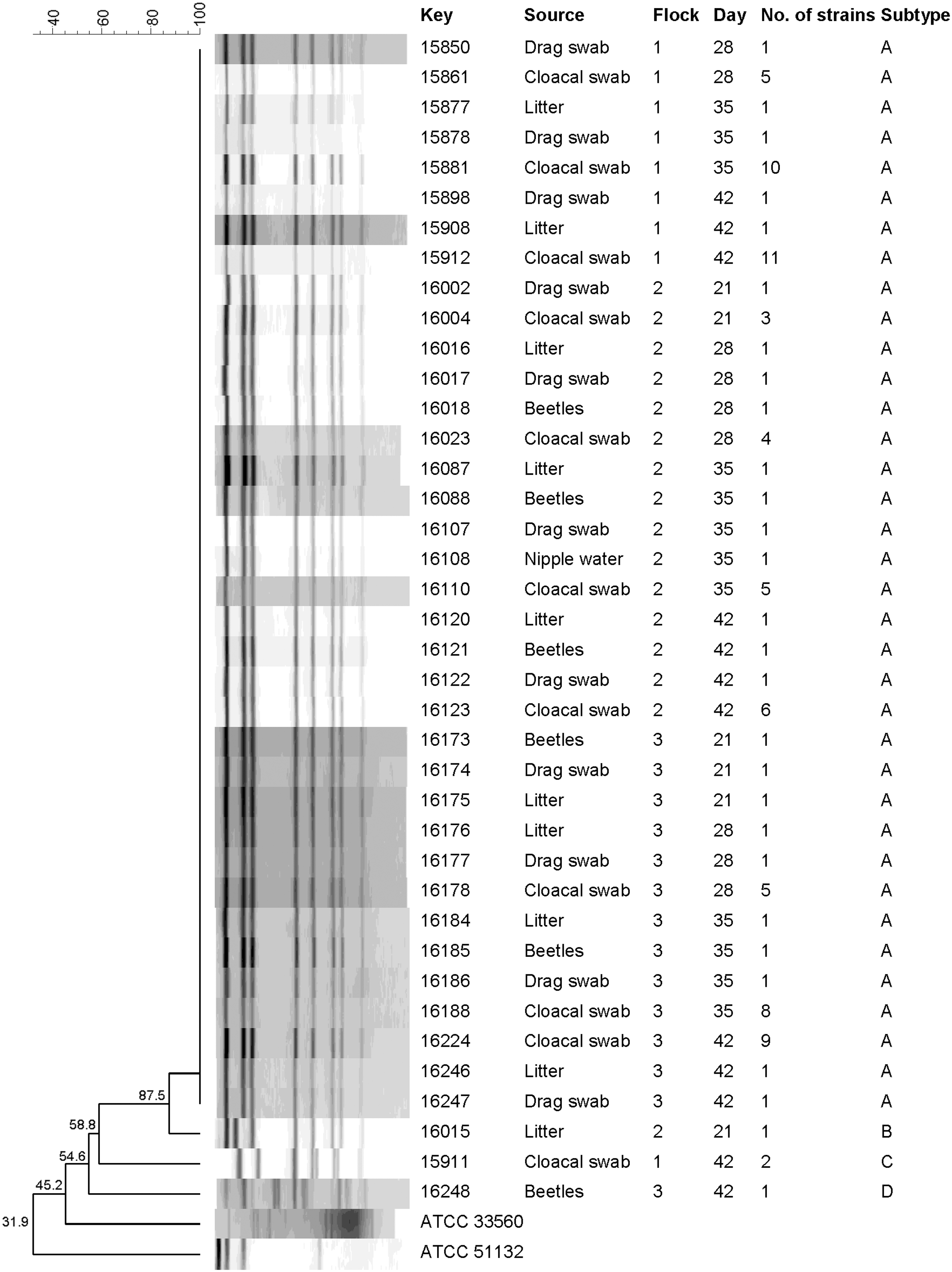
Dendrogram derived from PFGE analysis of Campylobacter jejuni strains isolated from sentinel broiler flocks (C. jejuni ATCC 33560 and Arcobacter skirrowii ATCC 51132 were included as control strains). PFGE, pulsed-field gel electrophoresis.
Discussion
Although the sentinel flocks tested Campylobacter negative upon chick placement, positive samples were detected over the course of rearing in reused litter. Broilers may be colonized by multiple Campylobacter species (Newell and Fearnley, 2003); however, only C. jejuni was detected in the analyzed samples. Other studies have shown as well commercial broiler flocks colonized exclusively by a single Campylobacter species (Messens et al., 2009; van Gerwe et al., 2009), which might be gradually replaced by others (Bull et al., 2006). Although a limited Campylobacter variety would be expected in this study as it was performed in a controlled environment, this finding highlights the predominance of C. jejuni in relation to other species.
C. jejuni was first detected in the sentinel broilers at 21 d. Although the Campylobacter status of the breeder flocks was unknown, Campylobacter-specific maternal antibodies can provide partial protection against colonization when challenged by homologous and heterologous C. jejuni strains (Sahin et al., 2003). Such protective antibodies would explain the Campylobacter-negative broilers in the first week. The model used in this study estimated between 24.3 and 34.8 d as necessary to detect 95% of positive-sampled broilers. Therefore, after the first Campylobacter detection, the sentinel broilers remained positive until the end of the production cycle in the majority of cloacal swabs analyzed. This result is in agreement to other findings of highest Campylobacter prevalence at the end of the production cycle of commercial flocks (O'Mahony et al., 2011; Hutchison et al., 2017).
The majority of C. jejuni strains analyzed shared an indistinguishable pattern among the PFGE subtypes detected in sentinel flocks. Although genetically diverse Campylobacter populations are generally found on commercial farms (O'Mahony et al., 2011), identical subtypes have previously been described in C. jejuni isolated from broilers and their environment (Bull et al., 2006; Ridley et al., 2011; Thakur et al., 2013). The predominance of a particular subtype suggests the carryover of strains from one flock to the next (Newell and Fearnley, 2003). Moreover, this is consistent with a common Campylobacter contamination source within the broiler house.
Intestinal digesta from broilers reared on reused litter showed numerous bacterial genera, such as Lactobacillus, Faecalibacterium, and Ruminococcus (Wang et al., 2016). An increased number of Campylobacter in litter has been found for each day a broiler is growing (Hutchinson et al., 2017). Therefore, the accumulation of excreta on reused litter would expose the successive flock to residual bacteria. Interestingly, reused litter and drag swabs tested Campylobacter negative after the downtime and in the early rearing period, despite the detection of positive samples in the previous sentinel flock raised on the reused litter. Environmental contamination readily appeared once the broilers began shedding Campylobacter (Bull et al., 2006; Sibanda et al., 2018). Due to its fastidious growth requirements, Campylobacter usually depends on multiplication within the gastrointestinal tract of a host (Newell et al., 2011), suggesting that the Campylobacter growth in broilers was required to maintain environmental contamination in this study.
It is noteworthy that microbiota in reused or fresh litter might be affected by management (Wang et al., 2016). Similar to the management practiced in commercial farms, reused litter was windrowed during the downtime in this study. Such a procedure has become accepted to reduce the residual contamination before rearing the successive flock (Brooks et al., 2015) and has been previously shown to be effective against the proliferation of Salmonella Enteritidis in reused litter (Vaz et al., 2017). The finding that reused litter and drag swab samples tested negative after the downtime would suggest a reduction in residual Campylobacter after depopulation. Nevertheless, a previous study has shown that Campylobacter survived in reused litter (Kassem et al., 2010). Moreover, detection of negative litter samples in broiler farms with low Campylobacter prevalence might reflect the nonculturability of strains under environmental stress (Kassem et al., 2017). Shortened C. jejuni survival in litter has been related to it entering the VBNC state due to stressors, such as desiccation (Smith et al., 2016). Therefore, the risk for Campylobacter persistence should be taken into consideration.
Darkling beetles usually persist in broiler houses after cleaning and disinfection. Previous studies have shown a connection between Campylobacter-positive darkling beetles and Campylobacter-positive broiler flocks (Hazeleger et al., 2008; Hutchinson et al., 2017). However, darkling beetles tested C. jejuni positive only after broilers and other environmental samples were detected positive. The majority of C. jejuni strains isolated from darkling beetles shared the predominant subtype previously found in strains from other samples. Nevertheless, the finding of a particular subtype in a strain isolated from darkling beetles highlights their potential to carry alternative Campylobacter strains in the broiler farm and the ability to survive in specific niches.
Feed samples tested Campylobacter negative. Such finding might result from an inefficient detection of the isolation procedures used. However, feed is adverse to Campylobacter survival due to the low moisture content and low water activity associated with the temperature reached in the pelleting process (Shane, 2000). Although C. jejuni was isolated from one nipple water sample, subtyping revealed that it shared the main pattern found in strains isolated from other samples in the flocks. Previous studies have found Campylobacter-contaminated drinking water in flocks with Campylobacter-positive status (Messens et al., 2009; O'Mahony et al., 2011), which suggests that Campylobacter may survive in biofilms on water pipes (Hansson et al., 2018). Nipple water contamination, indeed, usually follows rather than precedes the colonization of a flock, revealing the consequence that Campylobacter shed from the broilers can be tracked through the water lines (Newell and Fearnley, 2003).
Although Campylobacter colonization of broilers might be postponed by standard biosecurity measures (Newell et al., 2011; Battersby et al., 2016), the effectiveness of such procedures depends on the continuous contamination pressure in farms. In this study, the predominance of an indistinguishable strain suggests a common and persistent contamination source within the flocks. Sentinel broilers most likely amplified the Campylobacter level, which maintained flock and broiler house contamination until the end of the producing cycle. Because the reuse of litter is critical to the environmental sustainability of broiler production, it is clear that there is a need to better understand the impact of such management on the ecology of Campylobacter on broiler farms.
Footnotes
Acknowledgments
The Brazilian Agricultural Research Corporation (Embrapa, Grant No. 030860400) funded this study. In memory of our colleague, Gláucio Mattos.
Disclosure Statement
No competing financial interests exist.