
Abstract
Streptococcus agalactiae is well recognized to cause a variety of infections in many animal species and humans. We aimed to investigate genetic relatedness of S. agalactiae strains isolated from humans and animal origins, including cattle and fish, using capsular gene typing, pulsed-field gel electrophoresis (PFGE), and multilocus sequence typing techniques. Our results revealed that S. agalactiae strains with capsular type Ia and ST103 were observed from all bovine isolates (17/17) and one human isolate (1/5). S. agalactiae strains with capsular type III and ST283 were detected among isolates from fish (5/5) and from humans (2/5). Two PFGE clusters containing isolates from mixed origins were demonstrated: one cluster of five fish and one human isolate, and another cluster of one bovine and one human isolate. In conclusion, the close genetic relationship among S. agalactiae strains isolated from humans and animal origins was evident.
Introduction
S treptococcus agalactiae or group B streptococcus (GBS) is a pathogenic bacterium causing infections in domestic animals such as fish, chickens, and cattle, and pet animals such as dogs and cats (Evans et al., 2002, 2009). S. agalactiae has become one of the major causes of bovine mastitis worldwide (Keefe, 1997). S. agalactiae was confirmed to cause a high-mortality outbreak of up to 30% of mortality in tilapia culture in Thailand (Suanyuk et al., 2008; Kayansamruaj et al., 2014) and many countries (Anshary et al., 2014; Aisyhah et al., 2015).
In addition to causing infection in animals, S. agalactiae has been well recognized to cause invasive neonatal infection in humans worldwide (Nandyal, 2008). S. agalactiae can also be isolated from gastrointestinal and urinary tracts in healthy adults, and genital tracts of healthy women (Farley et al., 1993). Moreover, the incidence of S. agalactiae infection in the elderly has increased in many countries (Ho et al., 2006; Phares et al., 2008; Matsubara and Yamamoto, 2009).
The source of infection associated with septicemia from S. agalactiae is usually not clearly identified. However, a study in France was able to demonstrate cases of septicemia of S. agalactiae in patients due to the consumption of contaminated food (Van der Mee-Marquet et al., 2009). Therefore, cross-species infection of S. agalactiae can also occur.
S. agalactiae has been frequently isolated from the milk of dairy cows with subclinical and clinical mastitis, and ranked among the top two most common bovine mastitis pathogens with prevalence up to 20% in northern Thailand (Boonyayatra et al., 2007). S. agalactiae infection has also been reported in farmed tilapia in Thailand (Suanyuk et al., 2008). Moreover, the number of human patients infected with S. agalactiae has been increasingly reported in northern Thailand (Chaiwarith et al., 2011). Our objective was to investigate genetic variations of S. agalactiae strains isolated from bovine mastitis cases and compare with isolates from fish and humans.
Materials and Methods
S. agalactiae isolates
A total of 27 S. agalactiae isolates from Chiang Mai, Thailand, which were isolated during 2011 to 2014, were included in the study. Among these strains, 17 were isolated from clinical and subclinical bovine mastitis cases in 7 smallholder dairy farms. All bovine isolates were originally cultured from quarter milk samples either from cows showing clinical signs of udder inflammation or from cows with positive results using California Mastitis Tests. Five strains were isolated from internal organs of seven tilapia showing clinical signs of streptococcosis in three farms. Another five strains were clinical isolates from septicemia human patients. All of these isolates were confirmed to be GBS using Gram's staining, hemolysis activity, catalase reaction, CAMP test, hippurate hydrolysis, and Lancefield grouping (Facklam and Washington, 1991). All confirmed GBS were frozen at −80°C in brain heart infusion (BHI) broth with 5% glycerol before further examination.
Genomic DNA extraction
Genomic DNA extraction was performed using the GF-1 Bacterial DNA Extraction kit (Vivantis Technologies, Selangor Darul Ehsan, Malaysia). Briefly, all frozen S. agalactiae isolates were regrown in BHI broth and incubated at 37°C overnight. Cultured broth tubes were centrifuged at 6000 g for 2 min at room temperature (25°C). The cell pellet was resuspended with 100 μL of Buffer R1. Cells were lysed using 20 μL of lysozyme (50 mg/mL; Vivantis Technologies) and incubated at 37°C for 20 min. Lysed cells were centrifuged at 10,000 g for 3 min and resuspended in 100 μL of Buffer R2. Protein was digested using 20 μL of Proteinase K and incubated at 65°C for 20 min. Then, 400 μL of Buffer BG was added and mixed until a clear solution was observed. DNA was separated using 200 μL of absolute ethanol, and transferred into the DNA binding column. The columns were centrifuged at 10,000 g for 1 min. The DNA-bound columns were then washed twice using 750 μL of wash buffer. Bound DNA was eluted using 50 μL of elution buffer. Genomic DNA was kept at −20°C until use.
Identification of S. agalactiae using polymerase chain reaction
All GBS isolates were confirmed to be S. agalactiae by amplification of specific DNA sequences using primer V1: 5′-TTTGGTGTTTACACTAGACTG-3′ and V2: 5′-TGTGTTAATTACTCTTATGCG-3′ as previously described (Meiri-Bendek et al., 2002). The polymerase chain reaction (PCR) was conducted using 2 × Taq Master Mix (Vivantis Technologies). The PCR mixture contained 25 μL of 2 × Taq Master Mix (1.25 U of Taq DNA polymerase, 1 × ViBuffer A, 0.2 mM dNTPs, and 1.5 mM MgCl2), 1 μL of V1 and V2 (10 μM), and 5 μL of DNA template (50–100 ng/μL). The PCR cycles included initial denaturation at 94°C for 4 min, 5 cycles of denaturation at 94°C for 45 s, annealing at 50°C for 45 s, and extension at 72°C for 45 s, 20 cycles of denaturation at 94°C for 45 s, annealing at 46°C for 45 s, and extension at 72°C for 45 s, and final extension at 72°C for 5 min. The PCR products were separated using 1.5% agarose gel electrophoresis, stained with ethidium bromide, and visualized using a fluorescence camera (Gel Doc 1000; Biorad, Hercules, CA). The expected size of the PCR product was 120 bp.
Capsular gene typing
All confirmed S. agalactiae isolates were examined for their capsular gene using the two multiplex PCR method described by Poyart et al. (2007). Primers are listed in Table 1. The PCR mixture and cycles were similar to those described in the previous section. PCR products were randomly selected to represent each capsular gene, extracted using NucleoSpin® Gel and PCR Clean-up kit (MACHEREY-NAGEL GmbH & Co. KG, Düren, Germany), and sequenced. DNA sequences were compared with sequences in the NCBI database using BLASTn (
Sequences, Gene Targets, and Product Sizes of Primers Used for Capsular Gene Typing
Pulsed-field gel electrophoresis analysis
Pulsed-field gel electrophoresis (PFGE) was performed as described for Streptococcus uberis genotyping (Boonyayatra et al., 2018). Briefly, S. agalactiae isolates were inoculated in BHI broth and incubated at 37°C overnight. The inoculated broth was adjusted for its turbidity to reach OD610 = 1.5, and centrifuged at 13,000 g for 5 min. The cell pellet was mixed with 240 μL of cell suspension buffer (10 mM Tris-HCl [pH 7.2], 20 mM NaCl, and 50 mM EDTA) and 60 μL of lysozyme (10 mg/mL; Vivantis Technologies) and incubated at 37°C for 10 min. Pulse-field certified agarose (1%) in buffer (10 mM Tris-HCl [pH 7.2] and 0.1 mM EDTA) was added to the mixture. The DNA plugs were subsequently incubated with 500 μL of PK-PK buffer (1% sarkosyl, 50 mg/mL proteinase K, and 250 mM EDTA [pH 9]) at 50°C for 2 h, and washed thrice using wash buffer (10 mM Tris-HCl [pH 7.6] and 0.1 mM EDTA). DNA plugs were incubated with the restriction enzyme “SmaI” (10 U) at room temperature (25°C) overnight. The plugs were loaded into a 1% agarose gel. A contour-clamped homogeneous electric field device (CHEF DRII) (Biorad) with pulse times of 5.3 to 34.9 s at 6 V/cm for 20 h was applied. Gels were stained with ethidium bromide and visualized under UV light.
Multilocus sequence typing
Multilocus sequence typing (MLST) was performed for all GBS isolates. Internal fragments (400–500 bp) of seven different housekeeping genes, as listed in Table 2, were amplified by PCR and sequenced as described previously (Jones et al., 2003).
Primer Sequences and Their Corresponded Targets Used for Multilocus Sequence Typing of Streptococcus agalactiae
Data analysis
DNA fragment profiles from PFGE were interpreted as described by Tenover et al. (1995) and analyzed using Bionumerics® version 7.6 software (Applied Maths BVBA, Kortrijk, Belgium). The similarity of the band patterns was calculated using Pearson's correlation coefficient and clustered using a dendrogram generated by the unweighted pair group of arithmetic mean method. Simpson's index of diversity (D) with 95% confidence intervals of the PFGE techniques was determined as described previously (Grundmann et al., 2001).
For MLST data, sequences were assembled and analyzed using Bio-edit version 7.2.1 (Hall, 1999). Each isolate was assigned a sequence type (ST) based on the seven integer numbers constituting its allelic profile. The sequences of all alleles and the composition of the STs identified in this study are available at
Results
Isolate collection and identification
All 27 isolates were β-hemolytic on bovine blood agar, Gram-positive cocci and positive for Lancefield Group B. Based on species-specific PCR, all isolates were confirmed to be S. agalactiae.
Capsular gene typing
All S. agalactiae isolates from fish (n = 5) and bovine milk (n = 17) origins were capsular type III and Ia, respectively, as shown in Table 3. Among five human isolates, four different capsular types were identified, including capsular type Ia (n = 1), II (n = 1), III (n = 2), and V (n = 1) as listed in Table 3.
Allelic Profiles, Sequence Types, and Capsular Types of S. agalactiae Isolates from Different Origins
ST, sequence type.
PFGE analysis
Among all 27 S. agalactiae isolates, 13 distinct pulsotypes composed of 6–14 DNA bands were identified. When pulsotypes were compared among S. agalactiae isolates from different origins, Dice similarity indices ranging from 39.3% to 100% were observed. Four clusters of genetically closely related isolates were observed, including clusters A, B, C, and D, as shown in Figure 1. Isolates within each cluster showed ≥80% similarity. Clusters A and B contained only bovine strains. All fish isolates were in cluster C. Two human isolates, NMC98 and NMC115, were closely related to five fish isolates and a bovine isolate in clusters C and D, respectively.

Dendrogram showing the genetic relatedness of the 27 Streptococcus agalactiae isolates determined by the UPGMA method for PFGE patterns. The dashed line represents the threshold of 80% Dice similarity index. Capital letters and boxes denote genetically closely related isolates sharing band similarities above the threshold. PFGE, pulsed-field gel electrophoresis; UPGMA, unweighted pair group of arithmetic mean.
MLST analysis
Four STs were identified. One new ST was discovered in this study: ST 652. All S. agalactiae isolates from fish (n = 5) and bovine milk (n = 17) origins were ST283 and ST103, respectively, as shown in Table 3. Among five human isolates, four different STs were identified, namely ST1 (n = 1), ST103 (n = 1), ST283 (n = 2), and ST652 (n = 1) as listed in Table 3. S. agalactiae ST652 was the founder of the genetic evolution among all examined isolates. The genetic evolution of ST652 was closest to ST283, followed by ST1 and ST103, as shown in Figure 2. Genotypes based on MLST analysis were similar to those using capsular gene typing, as illustrated in Figure 2.
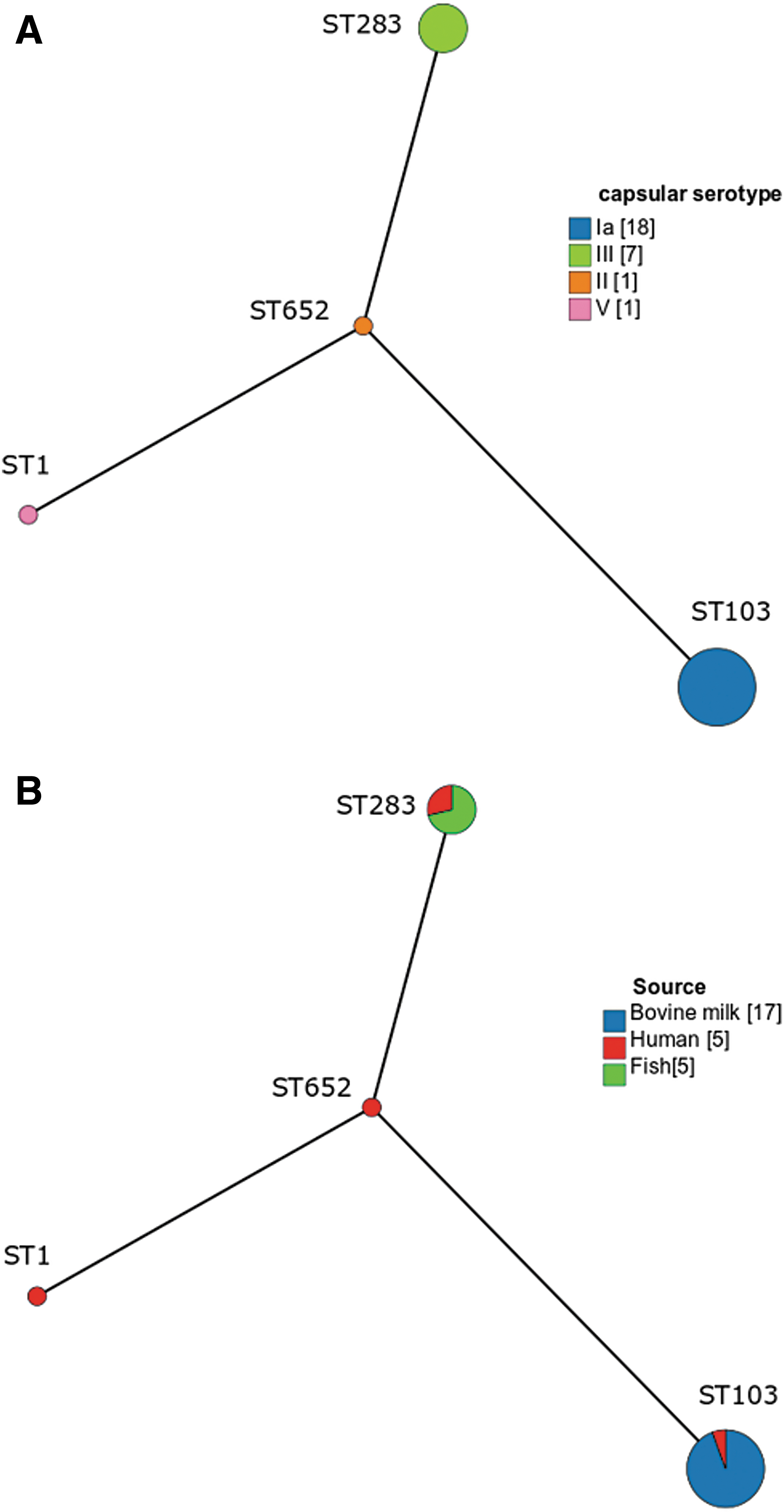
Minimum spanning trees demonstrating evolution of S. agalactiae ST1, ST103, ST283, and ST652 isolated from bovine milk, humans, and fish in northern Thailand. Sizes of circles are related to the number of isolates of each ST. Single locus variants are represented as connected lines.
Discussion
The relationships of S. agalactiae isolated from humans and animals have been investigated in several studies. Even though human and animal S. agalactiae isolates are largely distinct populations as suggested in various studies (Martinez et al., 2000; Bohnsack et al., 2004; Sukhnanand et al., 2005), a number of studies were able to demonstrate genetic relationships among human and animal strains, such as dairy cattle (Bisharat et al., 2004; Bohnsack et al., 2004; Oliveira et al., 2006), tilapias (Suanyuk et al., 2008), aquatic mammals, and fish (Delannoy et al., 2013).
In 2009, a molecular epidemiological study in France revealed cases of S. agalactiae septicemia caused by consumption of contaminated food items (Van der Mee-Marquet et al., 2009). Another study to investigate an outbreak of S. agalactiae infection in Singapore confirmed that S. agalactiae ST283 is a zoonotic pathogen that was related to the consumption of freshwater fish causing an outbreak of foodborne illnesses in 2015 (Kalimuddin et al., 2017). Recently, a study has demonstrated that Thai S. agalactiae ST283 were closely related to the Singaporean outbreak strain in 2015 (Kayansamruaj et al., 2018). In this study, we were able to demonstrate a strain of human S. agalactiae with an identical capsular type and ST, and closely related PFGE pattern (one different band) to S. agalactiae ST283 isolated from tilapia. This evidence indicates the possibility of interspecies GBS transmission between animals and humans in Thailand.
The most common ST among bovine S. agalactiae isolates reported in this study is ST103. S. agalactiae ST103 has occasionally been isolated in several species (Brochet et al., 2006; Bohnsack et al., 2008; Haenni et al., 2010). Recently, a study in China revealed that human S. agalactiae ST485 might evolve from ST103, which could infect both human and cattle (Wang et al., 2018). In 2006, Oliveira et al. had demonstrated that a bovine S. agalactiae ST103 was very different from a human S. agalactiae ST26, whereas these two strains showed a close genetic relationship based on their PFGE patterns (Oliveira et al., 2006). Similarly, S. agalactiae ST103 strains reported in this study were from both bovine and human origins. Moreover, a human isolate (NMC115) had a close genetic relationship with one of our bovine isolates using PFGE, while they were different STs. This finding supports the genetic relatedness among bovine S. agalactiae ST103 and human S. agalactiae. It is unclear how ST103 has become the predominant ST in bovine milk. Zadoks et al. suggested that an adaptation of this clone being able to survive in bovine udder might be the reason, which needed to be investigated (Zadoks et al., 2011).
In this study, more STs were observed among a limited number of human S. agalactiae, compared with animal strains. A study in human patients in northern Thailand reported that the most common site of S. agalactiae infection among nonpregnant patients was bloodstream infection (Chaiwarith et al., 2011), which could be originated from different primary colonization of various organ tissues. In contrast, invasion into the mammary glands of cattle and the internal organs of fish may require less genetic adaptation, which should be further investigated.
Conclusion
S. agalactiae isolated from animals contained less genetic diversity compared with human isolates. Evidence of close genetic relationships among S. agalactiae isolated from human, bovine milk, and fish was illustrated based on capsular gene typing, MLST and PFGE analyses. In northern Thailand, where various kinds of livestock farming are presented, cross-species transmission, especially zoonotic infection from consumption of contaminated milk and fish, can potentially occur. Genetic investigation of a pathogen using molecular tools will help to identify the mode of transmission and thereby improve control and prevention of such infectious diseases.
Footnotes
Acknowledgment
We thank Dr. Adithep Konputtar for his technical assistance.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by a grant from Chiang Mai University (R000012710).