
Abstract
To describe a historical baseline of antimicrobial resistance (AMR) profiles for human clinical Campylobacter species isolates obtained by laboratory surveillance in the province of Saskatchewan from 1999 to 2006; to determine if there were differences in resistance between Campylobacter jejuni and Campylobacter coli; and to determine if there were changes in the annual resistance levels in the two species. One thousand three hundred seventy-eight Campylobacter isolates were subjected to antimicrobial susceptibility testing using the E-test method. Annual resistance levels in C. jejuni and C. coli were compared using logistic regression models. One thousand two hundred (87.1%) isolates were C. jejuni and 129 (9.4%) were C. coli. Resistance in C. jejuni isolates included ciprofloxacin (CIP: 9.4%), erythromycin (ERY: 0.5%), and tetracycline (33.3%). CIP resistance in C. jejuni was higher in 1999 (15.5%, odds ratio [OR] = 3.96, p = 0.01), 2000 (12.7%, OR = 3.10, p = 0.01), 2005 (10.2%, OR = 2.47, p = 0.05), and 2006 (13.0%, OR = 3.22, p = 0.01) compared with 2004 (4.4%). C. coli had significantly higher CIP resistance (15.5%, OR = 1.78, p = 0.03), ERY resistance (13.2%, OR = 60.12, p < 0.01), multidrug resistance (2.3%, OR = 36.29, p < 0.01), and CIP-ERY resistance (3.1%, OR = 50.23, p < 0.01) compared with C. jejuni. This represents the first and most current report of AMR of the collective human Campylobacter isolates from a province in Canada and provides a baseline against which current and future resistance patterns can be compared. Fluoroquinolone resistance in C. jejuni isolates fluctuated from 1999 to 2006, including an increased prevalence in 2005–2006, while macrolide/lincosamide resistance remained very low. Human clinical C. jejuni isolates from Saskatchewan demonstrated resistance to multiple antimicrobials but had significantly less fluoroquinolone and macrolide resistance than C. coli isolates.
Introduction
Campylobacter is a leading cause of acute bacterial gastroenteritis in humans in Canada and the world (Moore et al., 2005; Ruiz-Palacios, 2007; Thomas et al., 2013). Food, particularly raw or undercooked chicken, is the main source of human infection, although other sources have been implicated (Effler et al., 2001; Olson et al., 2008). Resistance to antimicrobials may increase the public health burden of campylobacteriosis. People infected with Campylobacter strains resistant to quinolones have a longer duration of diarrhea (Engberg et al., 2004; Nelson et al., 2007). Quinolone-resistant Campylobacter infections are linked to an increased frequency of hospitalization and risk of adverse health events and death (Helms et al., 2005; Nelson et al., 2007). Resistance to fluoroquinolones, often used for empirical treatment of Campylobacter in adults, has increased in prevalence around the globe (Engberg et al., 2001; Moore et al., 2006; Luangtongkum et al., 2009; Smith and Fratamico, 2010). Canadian studies documented concerning resistance in Campylobacter species from humans, food animals, and retail foods. Fluoroquinolone and macrolide resistance has been documented in Campylobacter isolates from chicken, swine, and humans in Canada (Johnson et al., 2008; Cook et al., 2009; Rosengren et al., 2009; Agunos et al., 2013; Riley et al., 2015; Government of Canada, 2017). Fluoroquinolone and erythromycin (ERY) resistance emerged and increased in human isolates from the late 1980s to 2013 (Gaudreau and Gilbert, 1998, 2003; Gaudreau et al., 2007, 2014).
Saskatchewan is 1 of 10 provinces and three territories in Canada with ∼3% of Canada's population. Since the 1990s, Saskatchewan has maintained a collection of isolates from all reported human cases of campylobacteriosis, which is unique for Canada. We tested antimicrobial susceptibility for those collected from 1999 to 2006. These data provide a historical baseline of antimicrobial resistance (AMR) surveillance among Campylobacter species in Saskatchewan. The objectives of this study were to (1) describe AMR profiles for human clinical Campylobacter species isolates from the province of Saskatchewan from 1999 to 2006; (2) identify differences in the prevalence of resistance to antimicrobials between Campylobacter jejuni and Campylobacter coli; and (3) detect changes in the annual prevalence of resistance in the two species.
Materials and Methods
From January 1, 1999, to December 31, 2006, the Saskatchewan Disease Control Laboratory (the Laboratory) in Regina received fecal samples for Campylobacter isolation and speciation. Samples were collected in Cary-Blair transport medium (prepared in-house) and cultured as soon as possible after arrival at the laboratory. All isolates received by the laboratory from the province of Saskatchewan were included in the study. Isolates were accompanied with the following information: unique, anonymous patient identification, sex, age, and date of sample collection. The study was approved by the Research Ethics Boards of the Universities of Guelph and Regina.
Fecal specimens were cultured on charcoal-based selective medium (Becton Dickinson, Mississauga, Canada) without further enrichment and incubated in a microaerobic atmosphere (Anaerocult™ C sachets, Oxoid, Ottawa, Canada) at 42°C for 48 h. Isolates were identified and speciated using standard methods (Gram stain, catalase, oxidase, hippurate hydrolysis, and indoxyl acetate) (Nachamkin, 2019). Minimum inhibitory concentrations (MICs) were determined for the Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS) antimicrobial panel and breakpoints for Campylobacter species (Government of Canada, 2007) using E-test strips (AB BIODISK, Sweden) as described (Baker et al., 1991) according to the Clinical and Laboratory Standards Institute standard M45-A for fastidious bacteria (CLSI, 2006). The panel and concentration ranges (μg/mL) included the following: ciprofloxacin (CIP: 0.032–32), azithromycin (AZM: 0.032–256), clindamycin (CLI: 0.032–256), ERY (0.032–256), gentamicin (GEN: 0.032–256), nalidixic acid (NAL: 0.032–256), chloramphenicol (CHL: 0.032–32), and tetracycline (TET: 0.032–256) (Government of Canada, 2009). The MIC breakpoints (μg/mL) for resistance/susceptibility for each antimicrobial were as follows: CIP (≥4/≤1), AZM (≥8/≤2), CLI (≥8/≤2), ERY (≥32/≤8), GEN (≥8/≤2), NAL (≥64/≤16), CHL (≥32/≤8), and TET (≥16/≤4).
AMR prevalence was calculated by categorizing MICs according to resistance breakpoints (resistant or not); intermediate isolates were classed as nonresistant. Resistance prevalence results for CIP, ERY, and TET in C. jejuni and C. coli were compared for each antimicrobial and for antimicrobial patterns, including resistance to CIP and ERY, pan-susceptibility, or multidrug resistance (MDR; resistance to three or more antimicrobial classes).
Prevalence differences by species and by year were assessed for inclusion in multivariable, logistic, or exact logistic regression models. Indicator variables were used for species and for year of the study, and significant model variables were assessed for interaction. The year 2004 was used as the referent value as it had the lowest CIP resistance. Variables were kept in models if two-sided p-values were ≤0.05. Variables that were not significant in a model were included if they acted as a confounder, defined as an alteration of any model coefficient by greater than 25%. Logistic regression models were assessed for fit using the Pearson goodness-of-fit test. Model residuals, leverages, and influences (delta/betas) were examined to determine their impacts on the models (Dohoo et al., 2009). All data were analyzed using Stata 11 ME (StataCorp, College Station, TX).
Results
From January 1, 1999, to December 31, 2006, 1378 Campylobacter species isolates from humans (804 male, 574 female) in Saskatchewan were tested for antimicrobial susceptibility by the laboratory. Of these, 1200 (87.1%) were C. jejuni and 129 (9.4%) were C. coli. There were 4 Campylobacter upsaliensis, 1 Campylobacter laridis, and 2 C. lari isolates, with the remaining 42 identified only as Campylobacter species. The median age of cases was 27 years (range 1.5 months to 97 years).
Resistance prevalence results and MIC distributions for C. jejuni are shown in Table 1. Resistance was highest for TET (33.3%), followed by CIP/NAL (9.4%), ERY/AZM/CLI (0.3%), and one isolate was resistant to CHL; all isolates were susceptible to GEN. Temporality of resistance (Fig. 1) was analyzed using annual logistic regression models (Table 2) for CIP, ERY, and TET. Annual observed CIP resistance in C. jejuni decreased significantly from 15.5% in 1999 and 12.6% in 2000 to 4.4% in 2004, but increased significantly to 10.2% in 2005 and 13.0% in 2006.
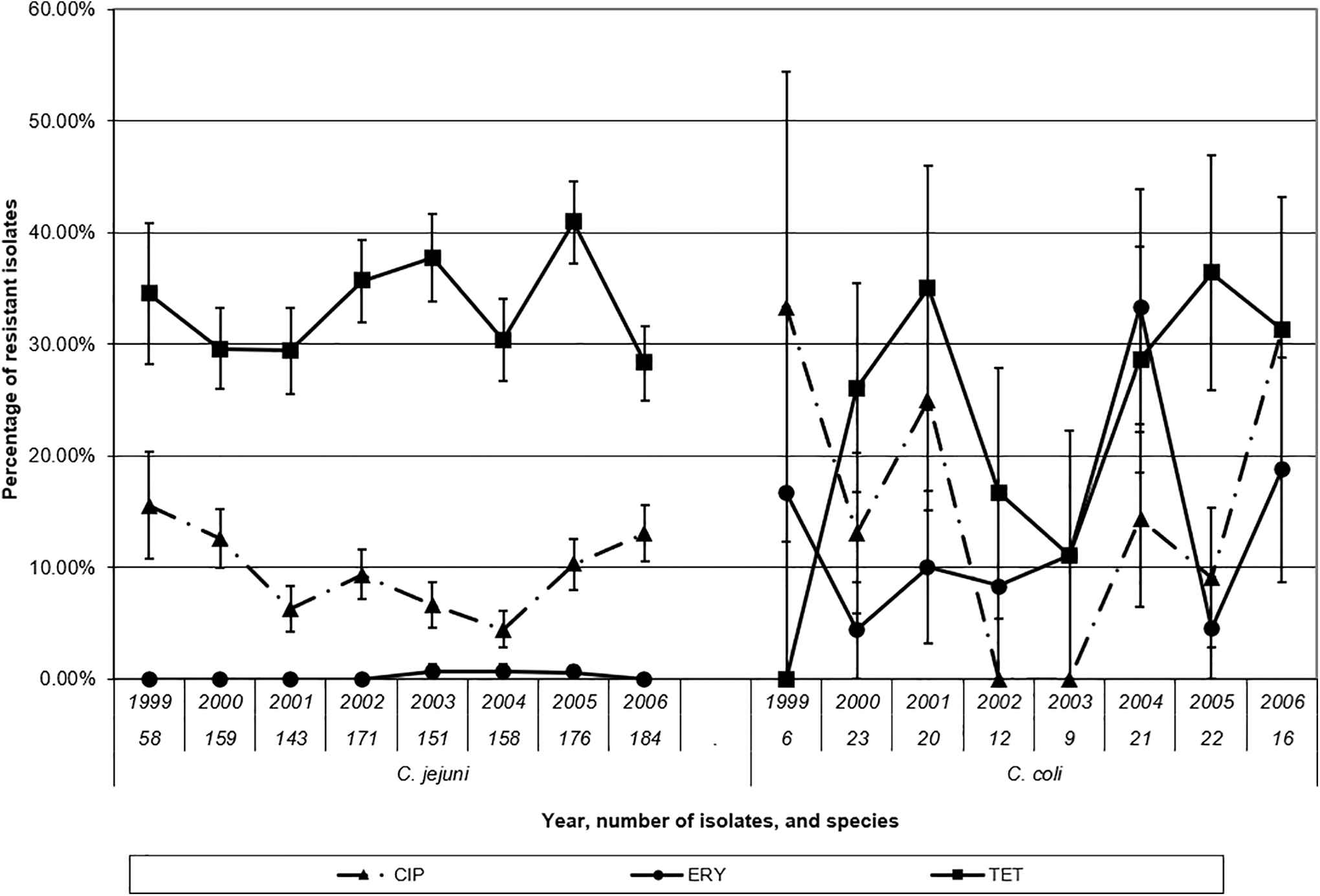
The annual prevalence of resistance to antimicrobials for human isolates of Campylobacter jejuni and Campylobacter coli from Saskatchewan, 1999–2006. Bars represent standard errors. CIP, ciprofloxacin; ERY, erythromycin, TET, tetracycline.
Antimicrobial Susceptibilities and Minimum Inhibitory Concentration Distributions of Human Clinical Campylobacter Species Isolates from Saskatchewan (1999–2006)

The Results of Ordinary Logistic Regression and Exact Logistic Regression Models for Antimicrobial Resistance in Campylobacter jejuni and Campylobacter coli from Humans in Saskatchewan (1999–2006)
Median unbiased estimate of odds ratio.
Resistance in three or more drug classes (quinolones: CIP or nalidixic acid).
CIP, ciprofloxacin; ERY, erythromycin; MDR, multidrug resistance; TET, tetracycline.
Resistance prevalence results, MIC distributions, and the logistic regression models for C. coli are shown in Tables 1 and 2. Resistance was highest for TET (27.1%), which did not differ significantly from C. jejuni (Table 2). For C. coli, the prevalence of resistance to NAL, CIP, and ERY was 17.1%, 15.5%, and 13.2%, respectively. CIP resistance in C. coli was significantly higher than in C. jejuni (Table 2). Year was significant to the CIP model that included both species (p = 0.02) and showed a similar annual trend in CIP resistance to the model with C. jejuni alone. The interaction between species and year was not significant (p = 0.10). ERY resistance was also significantly higher in C. coli compared with C. jejuni (Table 2), but year was not significant to the model with both species (p = 0.16) or the model with C. coli alone (p = 0.16), nor was it acting as a confounder.
Antimicrobial susceptibility profiles of C. jejuni and C. coli are shown in Table 3. There were 9 and 11 distinct profiles for C. jejuni and C. coli, respectively. The most common pattern for both species was pan-susceptibility to all eight antimicrobials (Table 3). There were 62.8% pan-susceptible C. jejuni and 56.6% C. coli isolates (p > 0.05) (Table 2). Single resistance to TET was the next most common profile for both C. jejuni and C. coli. Isolates resistant to NAL-CIP or those to TET-NAL-CIP were also reasonably common in both species. No C. jejuni isolates were resistant to both CIP and ERY or were MDR. In contrast, 2.3% of C. coli isolates were MDR and 3.1% were resistant to both CIP and ERY, half of which were also resistant to TET. Exact logistic regression models found that the prevalence of C. coli isolates with CIP and ERY resistance or MDR was significantly higher compared with C. jejuni (Table 2).
The Antimicrobial Resistance Profiles of Campylobacter jejuni and Campylobacter coli Isolates from Humans in Saskatchewan (1999–2006)
Quinolones: CIP, ciprofloxacin; NAL, nalidixic acid.
Macrolides/lincosamides: AZM, azithromycin; CLI, clindamycin; ERY, erythromycin.
CHL, chloramphenicol; GEN, gentamicin; TET, tetracycline.
Diagnostics for all ordinary logistic regression models did not detect any lack of fit or obvious outliers or influential covariate patterns.
The AMR profiles of the remaining Campylobacter species isolates other than C. jejuni and C. coli are shown in Table 4. Both C. lari isolates were resistant to NAL, while only one was resistant to CIP. The single C. laridis isolate was resistant to TET-NAL-CIP and the single C. upsaliensis was pan-susceptible to all eight antimicrobials. Of the 42 isolates designated as Campylobacter species, 23 were pan-susceptible. Seven of these isolates were resistant to CIP, while one of these also harbored resistance to all eight antimicrobials.
The Antimicrobial Resistance Profiles of Other Campylobacter Species Isolates from Humans in Saskatchewan (1999–2006)
AZM, azithromycin; CHL, chloramphenicol; CIP, ciprofloxacin; CLI, clindamycin; ERY, erythromycin; GEN, gentamicin; NAL, nalidixic acid; TET, tetracycline.
Discussion
This study described the AMR profiles for human clinical Campylobacter species isolates for the province of Saskatchewan from 1999 to 2006. This represents the first report of the AMR of all reported human isolates of Campylobacter in a province in Canada over an extended period and is important baseline data against which future AMR trends in Saskatchewan can be compared. Unfortunately, surveillance of AMR among human clinical isolates of Campylobacter in Canada is very limited. These represent the most current, collective provincial data in the country. This organism was among the first priority organisms identified for AMR surveillance in Canada (Amaratunga et al., 2016) and CIPARS has recently included limited data from selected provinces (Government of Canada, 2019). The overall prevalence of CIP resistance in C. jejuni was 9.4%. In C. jejuni, there was a significant decrease in CIP resistance in 2004 compared with 1999 (15.5%) and 2000 (12.7%), with a subsequent significant increase in the last 2 years of the study (10.2% in 2005 and 13.2% in 2006).
These data suggest that the prevalence of CIP resistance in human clinical isolates of Campylobacter from Saskatchewan was lower than other parts of Canada reported during the same time period. In 2001, the prevalence of CIP resistance in C. jejuni was 6.3% compared with 47% in Québec (Gaudreau and Gilbert, 2003). Further work in Québec from 2002 to 2006 found 44.3% of isolates to be CIP resistant (Gaudreau et al., 2007) compared with a high of 13.2% in 2006 in Saskatchewan. In Alberta, 31% of all human cases of Campylobacter species from 2004 to 2005 were resistant to CIP (Johnson et al., 2008) compared with 5.6% (2004) and 10.6% (2005) for C. jejuni in Saskatchewan. These differences in AMR may be due to varying Campylobacter strains, food consumption, travel patterns, testing practices, or antimicrobial use in humans or animals in these geographical regions. The Alberta study obtained cases from two of five health regions within the province and did not report resistance by Campylobacter species, although 196/210 isolates were C. jejuni. The Québec studies obtained isolates from hospitalized patients in Montréal. Resistant C. jejuni infections may be more severe and as a result, more likely to seek or require medical attention at a hospital (Nelson et al., 2004). In contrast, the Saskatchewan data comprised all culture-confirmed human isolates from the province and are therefore highly representative of a provincial sample of cases of a notifiable enteric disease obtained by passive surveillance.
Among C. jejuni, the observed prevalence of resistance to macrolide/lincosamide antimicrobials (ERY, AZM, and CLI) was low compared with other studies in Canada and some developed countries (Engberg et al., 2001), but was comparable with surveillance results from the United States in 2004/5 and the United Kingdom in 2002 (Campylobacter Sentinel Surveillance Scheme Collaborators, 2002; Gupta et al., 2004; CDC, 2007, 2008). The interpretive breakpoint for ERY resistance increased from ≥8 to ≥32 μg/mL by CIPARS in 2005 to harmonize with the U.S. National Antimicrobial Resistance Monitoring System in 2004 (CDC, 2007; Government of Canada, 2007). The 2001 Québec study reported 12% ERY resistance using the old breakpoints. When the old ERY breakpoints (≤0.5 μg/mL/≥8 μg/mL) were applied to this complete Saskatchewan collection of 1200 C. jejuni isolates (1999–2006), the prevalence of resistance did not change from 0.3%, although 85.1% had intermediate susceptibility (1–4 μg/mL). Care must be taken when comparing studies that use different MIC breakpoints, highlighting the importance of breakpoint harmonization to allow for meaningful comparisons of resistance prevalence between regions and over time.
C. jejuni was the most common isolate from human cases, which is comparable with other studies, including other provinces in Canada (Engberg et al., 2001; Johnson et al., 2008; Cook et al., 2009; Gaudreau et al., 2014). There was significantly higher resistance to quinolones and macrolides among C. coli compared with C. jejuni from Saskatchewan. The basis for this difference is not known, but may be attributed to different agricultural sources and their varied antimicrobial use patterns. The majority of Campylobacter in poultry are C. jejuni, compared with C. coli in pigs (Varela et al., 2007; Deckert et al., 2010, 2013; Rosengren et al., 2010). Although there has been no fluoroquinolone approved for use in poultry in Canada since 1997 (Engberg et al., 2001), its extralabel use is not prohibited. Fluoroquinolone resistance may offer a fitness advantage to C. jejuni, allowing it to persist in poultry even after withdrawal from use (Luo et al., 2005; Price et al., 2007). In contrast, Rosengren et al. reported high ERY resistance in Campylobacter isolates from swine in Alberta and Saskatchewan associated with macrolide treatment, which is common in the industry (Rosengren et al., 2010). While quinolone and macrolide resistances in C. jejuni and coli are both the result of chromosomal mutation (Engberg et al., 2001), there may be species differences in the rate of acquisition or retention of such mutations in the population, warranting further study (Aarestrup et al., 2008).
In 2005/2006, CIPARS started testing retail chicken for Campylobacter in Saskatchewan. Fluoroquinolone resistance in C. jejuni and coli was 0% for both in 2005, with 1/42 C. jejuni isolates being CIP-resistant in 2006 (Government of Canada, 2007, 2009; Agunos et al., 2013). However, CIP resistance from human clinical C. jejuni isolates in this study was higher in 2005 (10.2%) and 2006 (13.0%) compared with retail chicken. Interestingly, fluoroquinolone resistance in retail chicken Campylobacter species subsequently increased to similar levels in 2009/10 (Agunos et al., 2013), but there are no comparable human data.
This apparent disparity in CIP resistance between C. jejuni from humans and retail chicken samples in Saskatchewan during this time period raises questions about the source of infection. There may be other sources of resistant infection than retail chicken purchased in the province. Some resistant infections may be due to human fluoroquinolone use (Adler-Mosca et al., 1991; Wretlind et al., 1992; Smith et al., 1999). A Danish retrospective, registry-based, case/control study of humans with Campylobacter species infections found that fluoroquinolone use increased the subsequent risk of culture-confirmed diagnosis of Campylobacter species (Koningstein et al., 2011). This risk was significantly higher for isolation of fluoroquinolone-resistant Campylobacter species compared with susceptible isolates. Clinical knowledge suggests that CIP was commonly prescribed to humans in Saskatchewan for uncomplicated diarrhea and respiratory infections in the mid-1990s, but decreased starting in 1999 (G.B. Horsman, personal communication).
Some of these resistant isolates may have been obtained during foreign travel, which is a well-documented risk factor for acquiring a resistant Campylobacter infection in developed countries (Gaunt and Piddock, 1996; Smith et al., 1999; Engberg et al., 2004; Kassenborg et al., 2004; Johnson et al., 2008; Evans et al., 2009; Gaudreau et al., 2014). Travel destinations in these studies included the following: Europe, the Mediterranean, Southeast Asia, the Middle East, and Latin America. However, the U.S. studies indicated that while foreign travel and prior fluoroquinolone treatment were potential risk factors, the majority of resistant infections were acquired domestically and were not the result of fluoroquinolone treatment (Smith et al., 1999; Kassenborg et al., 2004). There is also the possibility for selection bias for resistant cases, as they may have more severe signs of disease and be more likely to seek medical attention, artificially increasing the prevalence of CIP resistance in the group of human isolates compared with retail chicken (Nelson et al., 2007).
This study was limited by incomplete individual epidemiologic information available for cases. This prevented identification of potential exposures for people colonized with CIP-resistant Campylobacter in Saskatchewan, including differentiation of domestic from travel-related strains. This may explain the apparent asynchrony between CIP resistance in human compared with retail poultry isolates (Riley et al., 2015). These baseline data represent the first complete provincial collection of isolates in Canada for an extended time. The only other comparable report is from Ontario (2011–2013) (Riley et al., 2015) that tested a portion of the provincial isolates for this period (Public Health Ontario, 2014). The Saskatchewan data represent a provincial sample of Campylobacter cases based on passive surveillance. However, resistance may be higher in this isolate collection due to the relative underreporting of human Campylobacter infection in Canada (Thomas et al., 2013) and the potential selection bias for resistant cases due to a higher likelihood of severe illness (Nelson et al., 2004).
Conclusions
This is the first report of AMR of all reported human isolates of Campylobacter species from a province in Canada and is important baseline data against which future AMR trends in Saskatchewan can be compared. This study detected increased CIP resistance in C. jejuni in 2005–2006 and very low macrolide/lincosamide resistance. Resistance to fluoroquinolones and macrolides/lincosamides and MDR was significantly higher in C. coli than C. jejuni. Human isolates of C. jejuni in Saskatchewan had higher resistance than that in retail chicken in 2005 and 2006. Further investigation is warranted to elucidate risk factors and the role of agricultural and human antimicrobial use in human infection with antimicrobial-resistant Campylobacter.
Footnotes
Acknowledgments
Support for this work included the following: the Blake Graham Fellowship (Ontario Veterinary College), the Laboratory for Foodborne Zoonoses and the Centre for Foodborne, Environmental and Zoonotic Infectious Diseases (Public Health Agency of Canada), the Saskatchewan Disease Control Laboratory (Saskatchewan Ministry of Health), and computational infrastructure obtained through the Canada Foundation for Innovation and Ontario Ministry of Research and Innovation (D.L. Pearl).
Disclosure Statement
The authors do not have a commercial or any other association that might pose a conflict of interest to this work.
Funding Information
This work was supported by the Blake Graham Fellowship at the Ontario Veterinary College, the Laboratory for Foodborne Zoonoses and the Centre for Foodborne, Environmental and Zoonotic Infectious Diseases at the Public Health Agency of Canada, and the Roy Romanow Provincial Laboratory (formerly the Saskatchewan Disease Control Laboratory) of the Saskatchewan Ministry of Health.