
Abstract
Bacillus cereus sensu lato (s.l.) is a group of bacteria commonly found in diverse environments, including foods, with potential to cause emesis and diarrhea. In Colombia, it is one of the main foodborne pathogens. The aim of this study was to determine the genomic and toxigenic heterogeneity of B. cereus s.l. isolated from ready-to-eat foods and powdered milk collected in day care centers of Medellin, Colombia. Of 112 B. cereus s.l. isolates obtained, 94% were β-hemolytic. Toxigenic heterogeneity was established by the presence of nheABC, hblCDAB, cytK2, entFM, and cesB toxigenic genes. The nheABC operon and entFM gene were most frequently detected in the isolates, whereas the cesB gene was not found. According to the toxin genes content, nine toxigenic profiles were identified. A 44% of isolates had profiles with all genes for nonhemolytic enterotoxin, hemolysin BL, and enterotoxin FM production (profiles II and IV). Pulsed-field gel electrophoresis analysis indicated a high genomic heterogeneity among the B. cereus s.l., with 68 isolates grouping into 16 clusters and 33 placed separately in the dendrogram. This study provides useful information on the safety of ready-to-eat foods and powdered milk in day care centers where children, a susceptible population, are exposed and it should incentive for more studies to understand the distribution of different toxin-encoding genes among B. cereus s.l. isolates, enabling detailed risk assessment.
Introduction
The Bacillus cereus group or B. cereus sensu lato (s.l.) includes spore-forming, facultative anaerobic, Gram-positive and motile rod bacteria commonly found in diverse environments (Ceuppens et al., 2013). Nine members of the group are recognized, Bacillus anthracis, Bacillus cytotoxicus, Bacillus mycoides, Bacillus pseudomycoides, B. cereus sensu stricto (s.s.), Bacillus thuringiensis, Bacillus toyonensis, Bacillus weihenstephanensis (Jiménez et al., 2013), and Bacillus wiedmannii (Miller et al., 2016). In addition, new species have recently been proposed based on molecular analysis (Liu et al., 2017). The heat-resistant pathogen B. cereus s.l. is naturally found in soil, which constitutes a risk factor for food contamination and foodborne infection or intoxication, because the spores survive cooking or pasteurization processes (De Jonghe et al., 2008).
Among the foods susceptible to be contaminated are ready-to-eat foods, sausages, and dairy products (raw, pasteurized and dried whole milk, and yoghurt) (Banykó and Vyletelová, 2009; Kumari and Sarkar, 2014; Yang et al., 2017; Alvarenga et al., 2018). In Colombian households and schools, B. cereus s.l. has been detected in cheeses, raw vegetables, ready-to-eat cereals, and flours (Forero et al., 2018), also in rice and not industrially prepared ready-to-eat foods (INS, 2011). In 2016, it was referred as one of the main foodborne pathogens in the country (Forero et al., 2018).
B. cereus s.l. produces food poisoning with diarrhea and/or emesis and causes outbreaks that are often underestimated (Papan et al., 2019). The emetic syndrome is generated by the emetic toxin, cereulide (Logan, 2012), encoded by the ces gene cluster (cesHPTABCD), located on a plasmid in strains belonging to a particular lineage of B. cereus s.s. (Økstad and Kolstø, 2011). The diarrheal syndrome is associated to three different enterotoxins coded in the bacterial chromosome. The hemolysin BL (HBL) is encoded by the hbl operon (hblCDA or hblCDAB that includes the hblB pseudogene), and the nonhemolytic enterotoxin (NHE) is encoded by the nhe operon (nheABC) (Logan, 2012). The cytotoxin K includes two variants, CytK1, encoded by the cytK1 gene, has a high cytotoxic activity, and CytK2, encoded by cytK2, is the most common variant (Stenfors et al., 2008). In addition, most B. cereus strains are potential producers of enterotoxin FM (EntFM), codified by the entFM gene (Ceuppens et al., 2013).
Knowledge about genomic heterogeneity, including intraspecific diversity and toxigenic profiles of B. cereus, is important to differentiate potential clinical isolates, to find contamination sources, track isolates along the food chain, and elucidate strain distribution within foods (De Jonghe et al., 2008). In Colombia there are reports of the detection of B. cereus enterotoxin genes (hblC, nheA, and cytK2) directly in powdered foods such as dietary complement for children and cassava starch (Sánchez et al., 2014), and of B. cereus strains producing HBL in rice and ready-to-eat cereals (Forero et al., 2018).
Considering that (1) the number of day care centers is rapidly growing around the world (Guirdham, 2019), (2) children deserve adequate care and special attention regarding food preparation and their safety, and (3) in Colombia B. cereus is an important cause of foodborne disease in scholar children (INS, 2007; Forero et al., 2018), this study aimed to determine the genomic and toxigenic heterogeneity of B. cereus s.l. isolates in ready-to-eat foods and powdered milk collected in day care centers of Medellin, Colombia. This information will contribute to food safety/risk assessment in institutions such as day care centers where children, a susceptible population, consume these foods.
Materials and Methods
Food sampling
A total of 327 food samples were randomly collected at day care centers (A–L) in Medellín, Colombia. Collections were performed four times spanning a month, 252 ready-to-eat foods were collected from September 2016 to November 2017 and included the following: fruit juice (n = 34), meat (n = 33), soup (n = 32), salad (n = 30), cooked rice (n = 29), milk beverage (n = 29), potato puree (n = 17), fried egg (n = 13), cheese (n = 12), corn-arepa (n = 8), fried ripe banana (n = 7), pasta (n = 5), bread (n = 2) and cooked cassava (n = 1). In addition, 75 powdered milk samples collected from May to August 2013 were included in the analysis.
Bacterial strains and processing
B. cereus s.l. were isolated and identified from ready-to-eat foods and powdered milk samples according to the ISO 7932:2004 method (ISO, 2004). B. cereus s.s. F4810/72 (emetic), ATCC 14579 (enterotoxigenic) and ATCC 21281 (enterotoxigenic), and B. thuringiensis 4D22 and 4AH1, were used as reference strains to standardize molecular procedures. Reference strains and food isolates were kept in cryo-tubes containing Luria-Bertani (LB) broth (Sigma, St Louis, MO) with 30% (v/v) glycerol (Merck, Darmstadt, Germany) and stored at −80°C. Lecithinase activity was determined on mannitol egg yolk polymyxin agar, MYP (Merck). Hemolytic activity was analyzed on blood agar with 5% sheep blood (Becton Dickinson, Heidelberg, Germany) (ISO, 2004).
Identification of B. cereus s.l. toxin genes by multiplex polymerase chain reaction
To identify B. cereus s.l. toxin genes, total DNA was extracted from 18 h grown LB broth cultures as previously described (D'Alessandro et al., 2007). DNA was quantified with a Nanodrop 2000 (Thermo Fisher Scientific, MA). To detect B. cereus s.l. toxin genes three multiplex polymerase chain reaction (mPCR) assays (1–3) were applied. The final reaction mixture for mPCRs consisted of 600 μM dNTPs mix, 4 mM MgCl2, 1.3 U of Taq DNA polymerase (Thermo Fisher Scientific), 1.6 μL 10 × reaction buffer, 100 ng of B. cereus s.l. DNA template, and 0.2 μM primer pairs added according to the genes amplified in each mPCR (Supplementary Table S1); for the internal transcribed spacer (ITS1), used as the internal amplification control in the first mPCR, 0.1 μM primers were added.
A mix of DNAs from B. cereus F4810/72 and B. cereus ATCC 14579 reference strains was used as positive control of toxin genes. A reaction mixture without DNA template was used as the PCR-negative control. Amplification in mPCRs was performed using a DNA thermocycler MyCycler (Bio-Rad, CA) with an initial denaturation at 94°C for 5 min, followed by 35 cycles of each, 20 s denaturation at 94°C, 40 s annealing at 50°C and 2 min elongation at 72°C, and a final incubation at 72°C for 5 min.
Genomic diversity of B. cereus s.l. isolates by pulsed-field gel electrophoresis
The pulsed-field gel electrophoresis (PFGE) method for characterization of B. cereus s.l. isolates used DNA plugs obtained as previously described (Gholamiandekhordi et al., 2006). The bacterial cultures had an optical density of 1.5 at 610 nm. DNA plugs were restricted using 20 U NotI endonuclease (Thermo Fisher Scientific) at 37°C for 4 h. DNA fragments were separated on a 1% SeaKem Gold agarose (Lonza, Basel, Switzerland) gel using a CHEF-Mapper (Bio-Rad), with the following optimized parameters: low molecular weight = 20 Kb and high molecular weight = 700 Kb, running time = 21 h, initial switch time = 5 s, final switch time = 80 s and tank temperature = 14°C. PFGE profiles were observed using a G:Box with a GeneSnap 7.12 (Syngene, Cambridge, United Kingdom) after staining the gels for 30 min in 400 mL of a 1 μg/mL ethidium bromide solution (Carl Roth, Karlsruhe, Germany) and destaining for 30 min in a 500 mL distilled water bath.
Data analysis
Cluster analysis was performed using BioNumerics v 7.6 software (Applied Maths, Sint-Martens-Latem, Belgium). The PFGE banding patterns were normalized by alignment with the Salmonella ser. Braenderup H9812 chromosome restricted with XbaI endonuclease, used as size standard marker (Dréan et al., 2015). A similarity analysis was conducted using the Dice coefficient and the unweighted pair group method with arithmetic mean, optimization, and tolerance settings at 1%. A similarity value of 84% was used to group the isolates in clusters (Gdoura-Ben Amor et al., 2018), based on visual determination.
Results and Discussion
B. cereus s.l. isolates recovered from foods
A total of 112 B. cereus s.l. were isolated from foods. There are no reports that these isolates were involved in food poisoning cases. The B. cereus s.l. were isolated from cheese (n = 22), cooked rice (n = 14), fried eggs (n = 4), fruit juice (n = 27), milk beverage (n = 13), potato puree (n = 2), powdered milk (n = 6), salad (n = 11), and soup (n = 13). The β-hemolytic activity was detected in 105 B. cereus s.l. isolates (94%), observed as a transparent halo around the colony. Similar to this result, other studies have reported high frequencies (>80%) of hemolytic activity in B. cereus isolates from ice cream (Arslan et al., 2014) and milk, ready-to-eat dishes and spices (Chaves et al., 2011). Of interest, of 47 isolates that were negative for the hbl genes, 45 presented β-hemolysis, which suggests that they probably produce other proteins with hemolytic activity such as cereolysin O, hemolysines II, or/and III (Ramarao and Sanchis, 2013).
All B. cereus s.l. isolates were able to degrade lecithin. This characteristic constitutes the first step to identify presumptive B. cereus according to international standards (ISO, 2004). However, some B. cereus isolates may lack the capacity to produce lecithinase (Chaves et al., 2011); therefore, these strains would not be identified in standard media as MYP (Fricker et al., 2008). The fact that hemolytic and lecithinase activities can vary among B. cereus strains suggests that although phenotypic analysis is a very important initial step for strain identification, it should be complemented with genomic characterization.
Toxigenic heterogeneity of B. cereus s.l. isolates
Toxigenic characterization of the isolates by mPCR showed that the nhe operon was present in all isolates, entFM gene in 92.9% of isolates, cytK2 gene in 24.1%, hblCDAB in 28.6%, hblCDA in 29.5%, and hblA in one isolate (0.9%); isolates carrying cesB gene were not identified. Similar to these results, a large distribution of toxin genes is observed in other countries all over the world such as Korea, Latvia, and Canada, with the enterotoxin genes (particularly, nhe operon and entFM) present in higher frequencies, and the emetic toxin genes were not detected (Lee et al., 2017; Saleh-Lakha et al., 2017; Fogele et al., 2018).
The nhe operon being present in all B. cereus s.l. isolates is in agreement with the suggestion that its high frequency is owed to its vertical inheritance, promoted by an unknown but relevant fitness function; conversely, the hbl operon and the cytK are commonly mobilized and horizontally transmitted (Böhm et al., 2015). In addition, the cytK2 gene detected in this study is the most common variant of the cytK gene present in B. cereus group members (Stenfors et al., 2008); the cytK1 gene is exclusive of B. cytotoxicus initially reported in France (Lund et al., 2000).
None of the isolates carried cesB gene, therefore, they were not able to produce the emetic toxin. This is a frequent finding (Altayar and Sutherland, 2006) with few exceptions of studies that have found B. cereus emetic strains in various foods (López et al., 2015; Park et al., 2016; Lee et al., 2017; Lin et al., 2017; Mudagza and Buys, 2017; Yang et al., 2017; Frentzel et al., 2018). In Colombia, the cesB gene was detected in DNA directly extracted from cassava starch (Sánchez et al., 2014); however, there are no reports about isolated emetic strains. Probably the reason for this low frequency is because the emetic strains belong to particular lineage of B. cereus s.s. (Økstad and Kolstø, 2011) that may have a low circulation in Colombia; in addition, the cereulide synthetase gene (ces) cluster is harbored on a megaplasmid horizontally transferred, called pBCE (Logan, 2012).
In total, nine toxigenic profiles were identified according to toxin genes presence (Table 1). Most of the isolates (43.8%) had profiles with all genes for NHE, HBL, and EntFM production (profiles II and IV), whereas 12.5% of the isolates had profiles with all genes for HBL, NHE, CytK, and EntFM (profiles I and III). The toxigenic profile VIII (25%) included isolates carrying genes for NHE and EntFM. Of interest, one isolate harbored an incomplete hbl operon (only hblA, profile VI), suggesting the presence of a toxin gene polymorphism and therefore, the production of a nonfunctional HBL because the three components, hblCDA, are necessary to generate an active toxin (Stenfors et al., 2008).
Toxigenic Profiles of Bacillus cereus sensu lato Isolated from Foods in Day Care Centers
CH, cheese; CR, cooked rice; FE, fried eggs; FJ, fruit juice; LS, lentil soup; MB, milk beverage; OS, oat soup; PM, powdered milk; PP, potato puree; SA, salad; SS, spinach soup; VS, vegetables soup.
The toxigenic profiles obtained are in agreement with those reported for B. cereus isolated from foods worldwide (Ankolekar et al., 2009; López and Alippi, 2010; Chaves et al., 2011; Samapundo et al., 2011; Arslan et al., 2014; Kim et al., 2014; Saleh-Lakha et al., 2017; Fogele et al., 2018), with B. cereus carrying predominantly genes encoding diarrheal toxins. Although many of these studies reported that the most common B. cereus toxigenic profile harbored all the genes for the NHE, HBL, and EntFM plus the cytotoxin K gene (Chaves et al., 2011; Samapundo et al., 2011; Fogele et al., 2018), in this study the predominant toxigenic profiles did not include cytK2 (profiles IV and VIII). The cytK2 gene was found in 24.1% of profiles; this result is in agreement with previous reports in which the cytK gene is found in low frequency (Lin et al., 2017; Mudagza and Buys, 2017), or is not even detected (Arslan et al., 2014; Silva et al., 2014; Frentzel et al., 2018).
When observing the distribution of B. cereus toxigenic profiles in relation to food type (Fig. 1A), or day care center (Fig. 1B), the results showed that a specific toxigenic profile could be isolated from various foods or day care centers; this indicates that there is no relationship between these variables, probably because the foods came from raw materials from diverse sources.
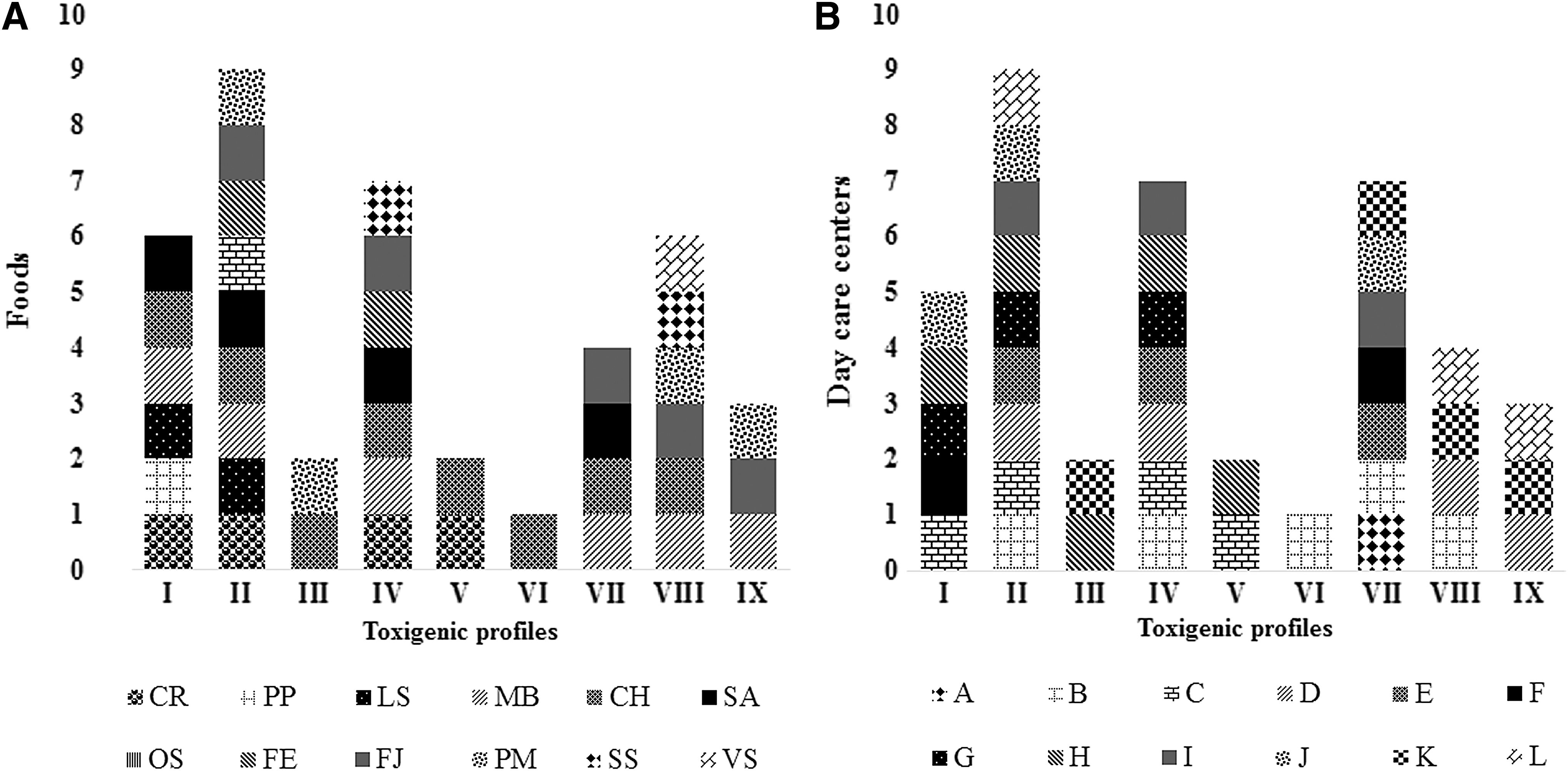
Distribution of Bacillus cereus sensu lato toxigenic profiles according to food types and day care centers.
Genomic heterogeneity of B. cereus s.l. isolates
PFGE fingerprinting was successful in separating the B. cereus s.l. isolates and a high heterogeneity was detected. In total, 105 PFGE patterns are shown in the dendrogram that correspond to 101 B. cereus s.l. isolates from foods and four reference strains (Fig. 2). No well-defined PFGE profiles were obtained for 11 isolates, even after several assays, making them untypeable and therefore, were not included in this analysis. Sixty-eight of the 101 isolates grouped into 16 clusters, the other 33 isolates and 4 reference strains clustered separately.
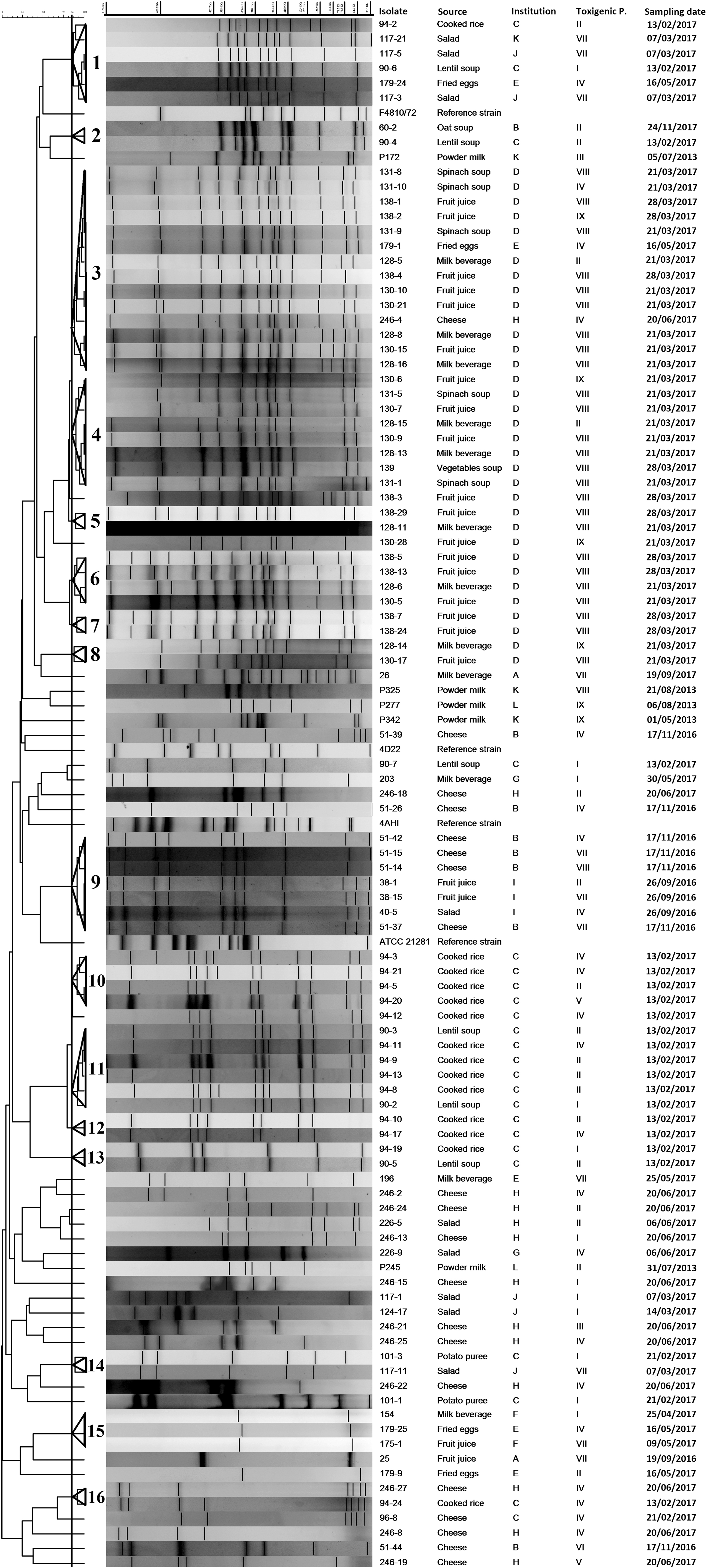
Pulsed-field gel electrophoresis dendrogram showing the relationship among Bacillus cereus sensu lato isolated from foods. The delineation level for clusters (indicated by bold vertical line) is visually set at 84% and clusters are shown in triangles.
Overall, a large genomic heterogeneity was detected and toxigenic profiles were widely distributed among the strains, with isolates from the same food type or harboring the same toxigenic profile presenting distinct banding patterns. In addition, particular genomic clusters presented different toxigenic profiles (e.g., clusters 1, 3, and 4) (Fig. 2 and Table 2). These findings indicate that the toxigenic profile is not related to the genomic background; in other words, no toxigenic profile was exclusively found on a specific cluster, as previously reported (Chaves et al., 2011; Lee et al., 2012).
Genomic Diversity and Toxigenic Profiles in Bacillus cereus sensu lato Isolates from Different Day Care Centers and Food Samples
CH, cheese; CR, cooked rice; FE, fried eggs; FJ, fruit juice; LS, lentil soup; MB, milk beverage; OS, oat soup; PFGE, pulsed-field gel electrophoresis; PM, powdered milk; PP, potato puree; SA, salad; SS, spinach soup; VS, vegetables soup.
Despite the wide genomic diversity, it was observed that clusters 10–13 were related; they included isolates exclusively from day care center C; furthermore, four different toxigenic profiles (I, II, IV, and V) were found in these clusters. Similarly, clusters 3–8 were related, and included B. cereus s.l. isolated mostly from day care center D with toxigenic profile VIII predominantly detected. In particular, clusters 5–7 carried highly related isolates, with a unique toxigenic profile; they were also from day care center D and differed in sampling date and food. This result may indicate a cross-contamination by persistent strains originating from the environment, raw materials, or the food-processing machinery in this day care center. Indeed, the analysis of the genomic diversity of isolates makes it possible to find contaminations with particular B. cereus strains in foods, as previously indicated (Samapundo et al., 2011; Mudagza and Buys, 2017).
Conversely, in day care center C, a particular food, for example cooked rice, was contaminated by a mixture of isolates belonging to various genomic groups (1, 10–13, and 16) and toxigenic profiles (II and IV predominantly) (Fig. 2). A possible explanation for this finding is related to the various steps taking place in the processing chain, in which diverse food contamination sources may be involved, as it was previously shown for pasta (Dierick et al., 2005).
Conclusions
Our results showed that B. cereus s.l. isolates present in ready-to-eat foods and powdered milk in the day care centers represent a potential risk for food poisoning in the children population given that all isolates contained enterotoxin genes. In addition, the high number of pulsotypes found indicates a high genomic heterogeneity among the B. cereus isolated from the various foods; such heterogeneity may even suggest that species within the B. cereus group may be present among the isolates and further studies should be carried out for their characterization and identification. Finally, this study provides useful information on the safety of ready-to-eat foods in day care centers and the results point out to the importance of additional studies to understand the distribution of different toxin genes among B. cereus s.l. isolates, enabling detailed risk assessment.
Footnotes
Acknowledgments
The authors thank Ann Van De Walle (ILVO, Technology and Food Science Unit, Melle, Belgium) for kindly providing technical support.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was partially supported by the Flanders Research Institute for Agriculture, Fisheries and Food (ILVO), Technology and Food Science Unit, Melle, Belgium. J.S.C. is a beneficiary received financial support for her doctoral studies from Colciencias (grant no. 647).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.