
Abstract
The current study investigated the emergence of multidrug-resistance (MDR), extended-spectrum beta-lactamase (ESBL)-producing Salmonella enterica serovar Heidelberg in broiler chickens and workers in poultry farms. A total of 33 S. Heidelberg isolates were recovered; 24 from the broiler cloacal swabs and 9 from the farm workers. All the S. Heidelberg isolates were tested for susceptibility to 11 antimicrobial agents and for the presence of resistance and virulence genes. MDR strains were found in 95.8% (23/24) and 88.8% (8/9) of the broiler and human isolates, respectively. Among the MDR strains, 66.6% of the broiler isolates and 55.5% of the human isolates were ESBL producing. The majority of broiler isolates showed resistance to ampicillin (100%) and ceftriaxone (91.6%), followed by ceftazidime and imipenem, (87.5%) and (75%). The resistance rate of the human isolates to those antibiotics were lower than the broiler isolates; ampicillin (88.8%), ceftriaxone (66.6%), ceftazidime (77.7%), and imipenem (66.6%). The resistance determinant genes found among the isolated strains was bla SHV-1, bla TEM-1, bla CMY-2, bla OXA-1, bla CMY-M2, bla PSE-1, and ampC. The most detected ESBL genes for broiler and human isolates were ampC (63.7%) and bla SHV-1 (56.6%), followed by bla CMY-M2 (48.5%), bla TEM-1 (39.4%), and bla OXA-1 (27.3%); whereas bla CMY-2 and bla PSE-1 were not detected. The finding of chromosomal and plasmid virulence genes revealed that the invA (100%), stn, sipC, and rck (72.8%), spvC (66.7%), ssr (63.6%), sopB (54.6%), and hilA and sipA (3.0%), while pefA and ssaR were absent. An elevated rate of MDR Salmonella Heidelberg in chickens is of potential great health risk. This signifies the role of the food of animal origin as a reservoir of MDR Salmonella that can affect the human health.
Introduction
S
In recent years, Salmonella enterica serovar Heidelberg (S. Heidelberg) has been found in poultry and poultry products in many countries all over the world (Alzwghaibi et al., 2018). In addition, the incidence of human infections by S. Heidelberg has increased by 25% from 1996 to 2005; even though there is an overall decrease in the number of cases of salmonellosis by 9% (Foley et al., 2008). Consumption of contaminated food materials is considered to be the main mode of transmission of S. Heidelberg to humans (CDC, 2006). There is a global concern about S. Heidelberg due to the emergence of multidrug-resistant (MDR) and virulent strains in animals (Hendriksen et al., 2011; Giuriatti et al., 2017). Surveillance reports have shown an increased prevalence of extended-spectrum β-lactams (ESBL)-resistant S. Heidelberg in food animals at slaughter, retail meat, and humans (Folster et al., 2012; CDC, 2017).
Drug resistance in pathogens has spread to various classes of antibiotics, including the β-lactams, which are widely used in treating bacterial infections in humans. The β-lactam comprises antibiotics such as penicillins, cephalosporins, monobactams, and carbapenems (Pitout et al., 2005). In Gram-negative pathogens, the β-lactams resistance is mainly attributed to a group of bacterial enzymes called β-lactamases. The prevalence and evolution of the β-lactamases seem to be directly linked to the clinical use of novel subclasses of β-lactams (Pitout et al., 2005). Indeed, in 1983, the extended-spectrum β-lactamases were first identified after the extended-spectrum oxyimino-cephalosporins were introduced to treat severe human infections (Bradford, 2001).
Bacteria producing those enzymes are resistant to the ESBL oxyimino-cephalosporins and monobactam antibiotics, but not cephamycins or carbapenems (Adesiji et al., 2014). Various forms of those drugs are used in veterinary medicine, thus contributing to the emergence of ESBL-producing pathogens (Abdellah et al., 2013; CDC, 2017).
With regard to virulence, pathogenicity of Salmonella species has been related to major virulence factors. Most of the virulence related genes are located on a virulence-associated plasmid and chromosomal Salmonella Pathogenicity Islands (SPIs) (De Jong et al., 2012).
Resistance to ESBLs is clinically important, because they are commonly used for treatment of severe salmonellosis in humans (CDC, 2017). Hence, there is an emergent need for determining the extent of spread of ESBL-producing and virulent S. Heidelberg strains among food animals and humans in contact to develop strategies to limit salmonellosis infection. Therefore, the objective of the present study was to track MDR S. Heidelberg strains in broiler farms and poultry workers in Egypt, with subsequent investigation of antimicrobial resistance and virulence gene determinants.
Materials and Methods
Sampling and study population
Cloacal swabs from 300 broilers showing diarrhea were randomly collected from 6 major broiler production regions in Egypt: Alexandria, Cairo, Monofya, Behera, Sharqia, and Fayoum.
Stool samples were randomly collected from 60 poultry workers at the selected poultry farms, they were all males and their ages varied from 20 to 35 years. Duplicate Samples were taken from each case.
Salmonella isolation and phenotypic characterization
Isolation of Salmonella was carried out as recommended by FDA (Andrews et al., 1998). In brief, one ml from each swab sample was preenriched in 9 mL of buffered peptone water (Oxoid, Healthy Family Co., Cairo, Egypt) and incubated at 37°C for 18 h. This was followed by inoculation of 0.1 mL of the preenriched broth into 10 mL of Rappaport-Vassiliadis broth (Oxoid) and incubation at 42°C for 24 h. A loopful of the incubated enriched inoculum was streaked on xylose lysine desoxycholate agar (Oxoid) and incubated at 37°C for 24 h.
The presumptive colonies were subjected to biochemical test, including triple sugar iron, orthonitrophenyl galactosidase, urease test, and IMViC tests (Andrews et al., 1998).
Serotyping
The isolates were subjected to serogroup characterization using a slide agglutination test with polyvalent and monospecific antisera against somatic and flagella antigens. The serotyping was performed in the reference laboratory center of Animal Research Institute, Cairo, Egypt.
Antimicrobial susceptibility test
Antimicrobial susceptibility testing was carried out on S. Heidelberg (n = 33) isolates using the Kirby-Bauer disk diffusion method using Mueller-Hinton agar (Difco), according to the Clinical and Laboratory Standards Institute guidelines (CLSI, 2017). A total of 11 different antibiotic disks which belong to eight groups of antimicrobials were used to confirm resistance of the isolates to one or more agents. The disks included and their corresponding concentrations were as follows: ampicillin (10 μg), amoxicillin/clavulanic acid (10 μg), imipenem (10 μg), ciprofloxacin (5 μg), gentamicin (10 μg), streptomycin (10 μg), ceftazidime (30 μg), ceftriaxone (30 μg), tetracycline (30 μg), doxycycline (30 μg), and sulfa/trimethoprim (25 μg). The antimicrobial susceptibility of the S. Heidelberg isolates was measured and the results were interpreted in accordance with criteria provided by CLSI, 2017.
MDR Salmonella was defined as resistance to one or more agents in three or more classes or groups of the tested drugs (Johnson et al., 2010; Magiorakos et al., 2012).
ESBL detection by double-disk synergy test method
ESBL production in S. Heidelberg was identified by the double-disk synergy test. Mueller-Hinton agar was inoculated with standardized inoculum of S. Heidelberg (corresponding to 0.5 McFarland tube). The amoxicillin plus clavulanic acid (AMX/AC; 20/10 μg) disk was inserted in the plate center and four test disks of ceftazidime (CAZ 30 μg), ceftriaxone (CRO 30 μg), cefotaxime (CTX 30 μg), and aztreonam (ATM 30 μg) disks were placed at 20 mm distance from the amoxicillin clavulanic acid disk. The plates were incubated overnight at 37°C.
Enhancement of the zone of inhibition of any one of the tested disks toward amoxicillin–clavulanic acid proposed the presence of the extended-spectrum beta-lactamases (Jarlier et al., 1988; Elhariri et al., 2017a).
Characterization of antimicrobial resistance and virulence determinants
Chromosomal and plasmid DNAs were extracted from 33 S. Heidelberg isolates by QIAamp DNA mini kit and plasmid Midi Kit (Qiagen, Hombrechtikon, Switzerland), respectively. The primers used in the current study for the detection of ESBLs resistance were bla SHV-1, bla TEM-1, bla CMY-2, bla OXA-1, bla CTX-M2, bla PSE-1, and ampC (Giuriatti et al., 2017). Besides 11 virulence genes (invA, stn, sipC, rck, spvC, sopB, ssr, hilA, sipA, pefA, and ssaR), they were also examined (Barilli et al., 2018).
The amplification cycles were performed using 12.5 μL master mix, 3 μL of DNA template, 0.25 μL of each primer in concentration (25 pmol), and nuclease free water up to 25 μL. Cycling conditions were done with a temperature profile of 94°C for 5 min, followed by 35 cycles of denaturation 94°C for 30 s, annealing temperature as in (Table 1) for 30 s, extension 72°C for 45 s, and final extension 72°C for 7 min. Uniplex primers used for the identification of genes are listed in Table 1.
Oligonucleotide Primers Encoding for Extended-Spectrum Beta-Lactamases-Encoding Genes & Virulence Determinants Genes
Phylogenetic analysis
All of the amplified invasion gene PCR products from the identified Salmonella isolates were partially sequenced, submitted to GenBank, and accessed the following numbers; KJ718885, KJ718887, KJ718874, KJ718875, KJ718876, KJ718877, KJ718878, KJ718880, KJ718881 KX524152, KX524153, KX524154, KX524155, KX524156, KX524157, KX524158, KX524159, KX524160, KX524161, KX524162, KX524163, KM282012, KM282013 KM282014, MF678530, MF678531, MF678532, MF678533, MF678534, MF678535, MF678536, MF678537 and MF678538. Sequence alignment was performed using Clustal W program (Altschul et al., 1997). Phylogenetic trees were constructed using Mega program version 7 (Tamura et al., 2013) with the neighbor-joining method to estimate the genetic relatedness between the chicken and human isolates that showed most common antibiotic resistance patterns.
Statistical analysis
The frequencies of antibiotic susceptibility and the presence of virulence genes were analyzed by Fisher's exact test and Chi-square test. p ≤ 0.05 was considered statistically significant. The test was performed with the GraphPad Prism 7 software.
Ethics statement
Protocols for the collection of samples as well as all methods were performed in accordance with the guidelines of the Institutional Animal Care and Use Committee (IACUC) of the Faculty of Veterinary Medicine, Cairo University, Egypt (VetCU1022019067). Oral consent was taken from all participating workers.
Results
Detection and identification of S. Heidelberg
The bacterial isolates were identified by phenotypical characterization and were confirmed by the presence of the invA gene, which is considered to be the common diagnostic target of Salmonella. We found that S. Heidelberg was the dominant serovar, representing 24/300 (8%) of the Salmonella-positive broiler isolates. As for the examined human stool samples 9/60 (15%), isolates were identified as S. Heidelberg.
Antimicrobial characterization and ESBL production
Of the 24 S. Heidelberg broiler isolates, 23 (95.8%) were MDR (Table 2). Similarly, eight of the nine (88.8%) S. Heidelberg human isolates were MDR. ESBL-producing strains were 66.6% and 55.5% for broilers and human, respectively. The antibiogram profiles of broiler strains showed the maximal resistance to ampicillin and ceftriaxone, 100% and 91.6%, respectively, followed by ceftazidime (87.5%) and imipenem (75%); whereas, the human isolates were less resistant to those antibiotics; ampicillin (88.8%), ceftriaxone (66.6%), ceftazidime (77.7%), and imipenem (66.6%). (Table 2) (Fig. 1).
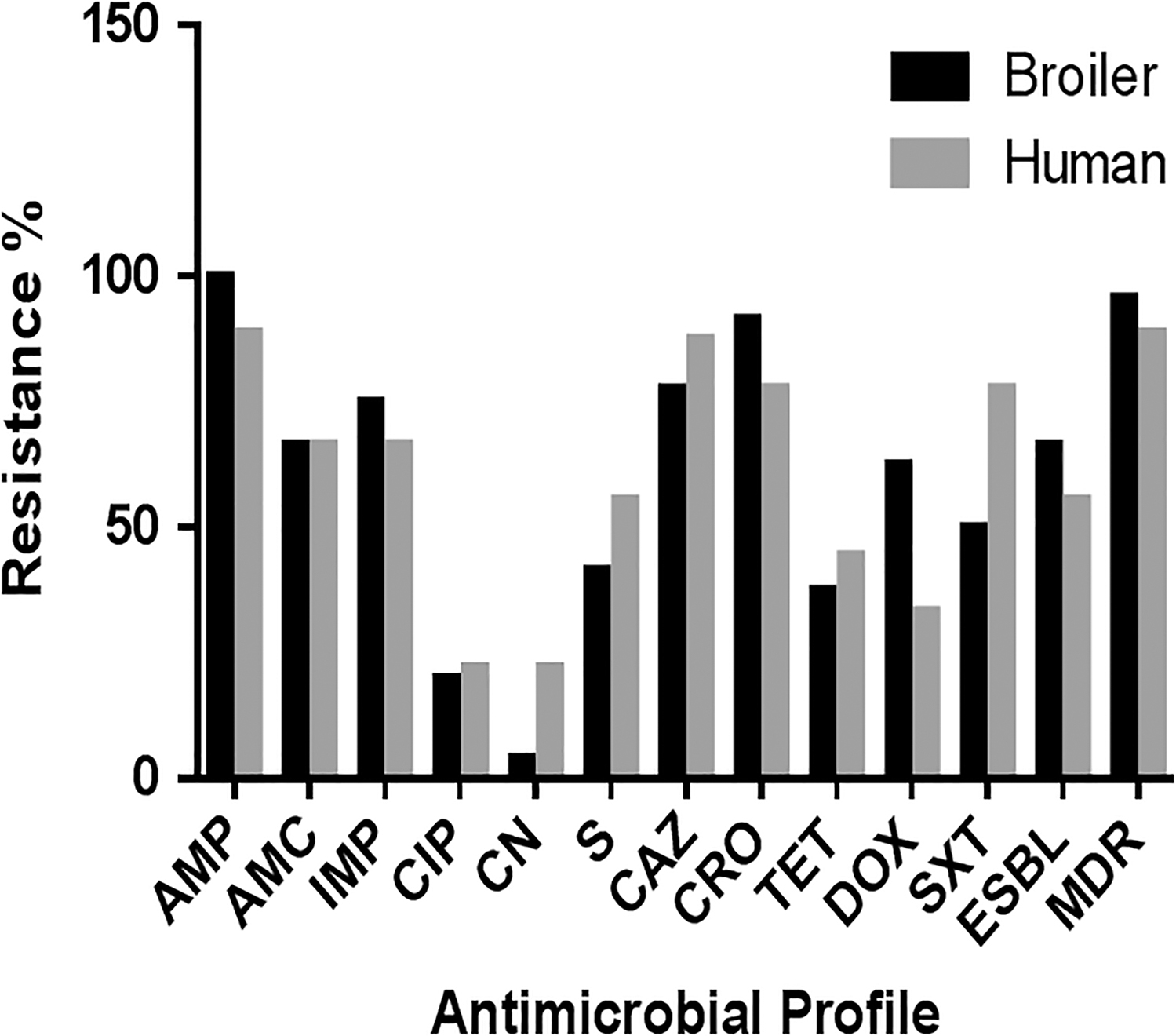
Antibiotic resistance pattern of different S. Heidelberg isolates from broiler chicken & human.
Antimicrobial Resistance Profile of S. Heidelberg Broiler & Human Isolates
R, Resistant; I, Intermediate; S, Sensitive; +, isolate is identified as ESBL.
Molecular detection of ESBL-encoding genes
The frequencies of ESBL-encoding genes had significantly higher p < 0.05 among broiler and human strains in general.
The antibiotic resistance genes of the isolated strains were bla SHV-1, bla TEM-1, bla CMY-2, bla OXA-1, bla CMY-M2, bla PSE-1, and ampC. The most detected ESBL gene for broiler and human isolates was ampC (63.7%), the second most prevalent gene was bla SHV-1 (56.6%), followed by bla CMY-M2 (48.5%), bla TEM-1 (39.4%), and bla OXA-1 (27.3%); whereas bla CMY and bla PSE-1 genes were not detected (Table 3).
Frequency of Extended-Spectrum Beta-Lactamases-Encoding & Virulence Genes to S. Heidelberg Strains Isolated from Broiler and Associated Poultry Workers
p < 0.05, Chi-square. Broiler versus Human S. Heidelberg isolates.
Virulence determinants
Regarding the detection of virulence determinants, there was a significant difference for the majority of the genes studied between broiler and human strains (Table 3). The detection of chromosomal and plasmid virulence genes revealed that all of the strains tested harbored invA (100%), stn, sipC and rck (72.8%), spvC (66.7%), ssr (63.6%), and sopB (54.6%). In contrary, only 3% of the strains carried hilA and sipA genes, while pefA and ssaR genes were negative (Table 3).
Based on combinations of virulence genes, 24 virulence profiles were observed among the tested strains. The most common virulence profile was VP6 and VP10, which was represented in three (9.0%), whereas VP1, VP5, and VP7 were estimated as two (6.0%), while VP2, VP3, VP4, and VP8 to VP24 were the least prevalent ones (Table 4).
Virulence Genes Profile of S. Heidelberg Strains Isolated from Broilers and Poultry Workers
+, presence of the gene; −, absence of the gene.
Phylogenetic and antimicrobial resistance patterns
Partial sequencing of S. Heidelberg strains based on invA gene, followed by phylogenetic analysis (Fig. 2) illustrated bunching of the isolates into three distinct clusters (A, B & C). The clusters were further subdivided into several clades that most probably had considerable sequence diversity. The cluster (A) contains the broiler strains with the most prevalent antibiotic resistance patterns to AMP, CRO, CAZ, IMP, AMC, and S and/or TET. The cluster (B) includes mainly the human strains with variable resistance patterns to two to more than five antibiotic groups. In contrast to clusters (A) and (B), cluster (C) is a part of a small monophyletic group, which encompasses both broiler and human strains without restricted antibiotic resistance pattern which is too related to other groups. Furthermore, there was no particular antimicrobial resistance pattern found to be specific to broiler or human strains. The three major monophyletic clusters, varied in the distance relatedness, the broiler, and human strains (Clade C), act as the major root and ancestor for broiler clade (A) and human clade (B).
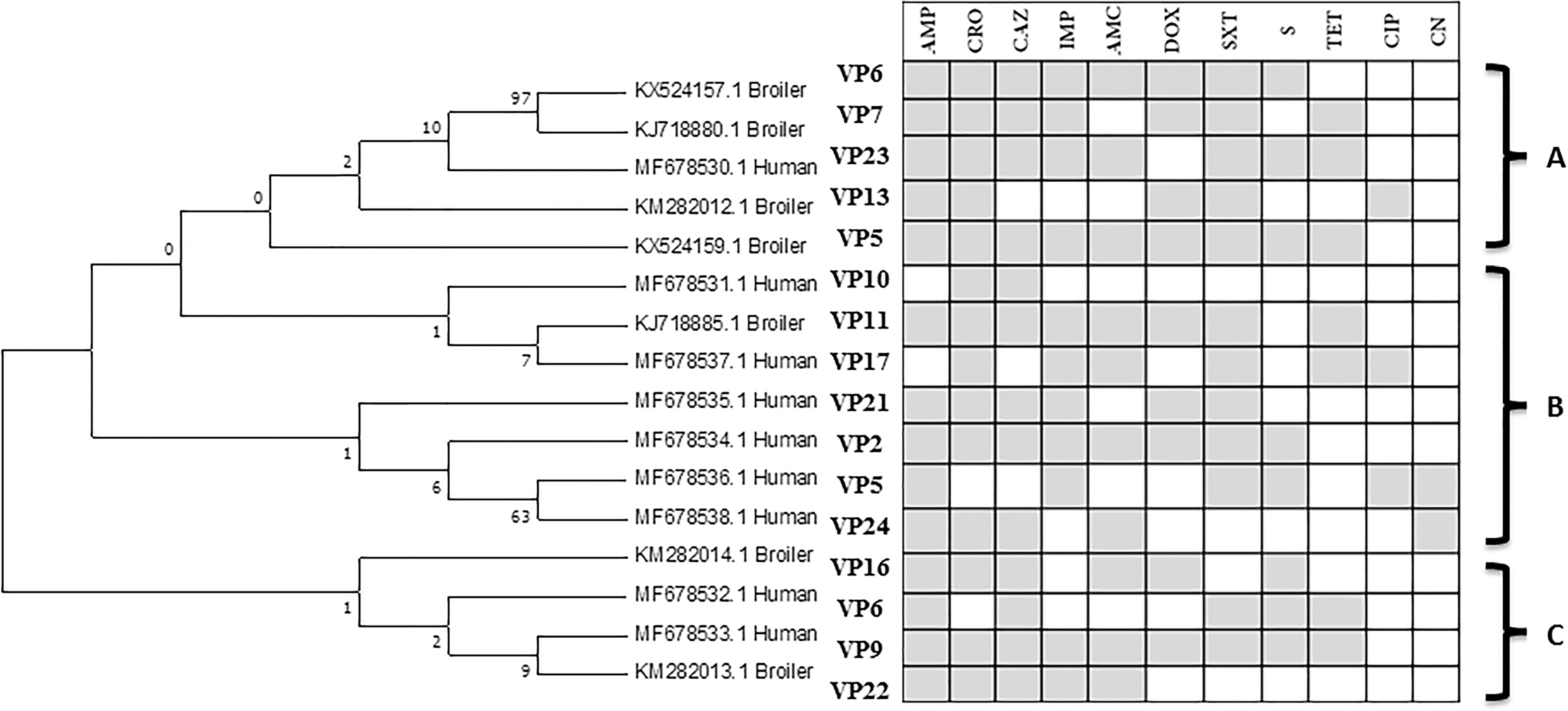
Phylogeny and AMR profile of S. Heidelberg isolates from this study. The phylogeny tree was constructed using the neighbor-joining method. The bootstrap consensus tree inferred from 1000 replicates. The percentage of replicate trees, in which the associated taxa clustered together in the bootstrap test (1000 replicates) is shown next to the branches. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. VP, virulence profile.
Discussion
Salmonellae are widely distributed in the environment, and can be found in livestock, wild animals, birds, and pet animals. Nowadays, there is an increasing prevalence of the organism in the global food chain, which results in the spread of diarrheal diseases in humans and animals. In addition, Salmonella have a tremendous ability to carry and spread resistance genes, posing a significant threat to the public health as well as economic problems (Pegues et al., 2005). Acquired antibiotic resistance in nontyphoidal Salmonella is a global challenge, particularly the MDR strains, which have been related to increased risk of severe infection, with limited choices of treatment and subsequent death (Nair et al., 2018).
Salmonella infection in chickens is recognized as an important contributor to human salmonellosis. In Egypt, a variety of poultry organs have been reported as important sources of salmonellosis infection with a percentage range of 3 up to 17% (Salem et al., 2017).
The current study revealed the recovery of S. Heidelberg from broiler cloacal swabs (9%) and from stool samples of poultry workers (15%) collected from six Egyptian governorates. The isolates were confirmed to be salmonellae by presence of invA gene, which is considered as an international standard procedure for detection of Salmonella (Malorny et al., 2003). The invA gene is positioned on Salmonella pathogenicity island 1(SPI-1) and is crucial for full virulence of Salmonella. The occurrence of antimicrobial resistant-Salmonella spp. isolated from foods of animal origin, especially poultry, has increased globally (Bacci et al., 2012). Antimicrobial-resistant S. Heidelberg strains have been found throughout the food production continuum (Nayak et al., 2004).
Presently, the prevalence and MDR patterns of Salmonella isolated from the broiler and human samples are high (Table 2). Most of the examined broiler isolates were MDR 95.8% (23/24), while human MDR isolates were 88.8% (8/9). The ESBL-producing strains were 66.6% for broiler and 55.5% for human. On the contrary, the CDC reported that from January 2015, outbreaks with S. Heidelberg were reported from 15 states in the United States, till 2018 to reach 35% (CDC, 2017).
The antibiogram profiles of the broiler strains showed the maximum resistance to ampicillin and ceftriaxone, 100% and 91.6%, respectively, followed by ceftazidime and imipenem, 87.5% and 75%, whereas human isolates were less resistant to these antibiotics, ampicillin (88.8%), ceftriaxone (66.6%), ceftazidime (77.7), and imipenem (66.6%) (Table 2).
The worldwide records of resistance in Salmonella spp. isolated from the poultry production chain are variable (Yan et al., 2010). It is worth mentioning that the present data go confirming on that after the introduction of antimicrobial agents into human and veterinary medicine as well as agriculture for growth promoting, the percentage of resistance has been steadily increasing in the isolated and investigated Salmonella strains in many regions all-over the world (Teuber, 1999). In the past, the antimicrobials recommended for susceptible Salmonella included the ampicillin, chloramphenicol, and trimethoprim-sulfamethoxazole. However, resistance to these antimicrobials has developed in many regions (McDermott, 2006). Furthermore, (Linto et al., 2018) found that among isolates of human origin, 30% percentage (47/154) contained at least one antibiotic resistance gene.
The S. Heidelberg strains encountered 100% resistance percentage against ampicillin, and also, it is noteworthy that many reports have mentioned that many Salmonella strains are resistant to multiple antimicrobials, including ampicillin (Karczmarczyk et al., 2010).
In addition, 80% and 96% of the strains tested were sensitive to ciprofloxacin and gentamicin, respectively, which represents an important finding because fluoroquinolones are the drugs of first choice in the treatment of invasive Salmonella infection (ETACAR, 1999).
The uncontrolled use of antibiotic in veterinary medicine resulted in emergence of MDR- ESBL-producing Salmonella spp., which might be due to the use of antibiotics mainly cephalosporin and ceftiofur, which are permitted in the veterinary practice, as well as extending the hydrolysis spectrum directed toward third-generation cephems and aztreonam (Shiraki et al., 2004). In the present study, the prevalence of ESBL-producing strains was 66.6% for broilers and 55.5% for humans. The Animals have the potential to act as reservoirs for a number of zoonotic infections, including those caused by pathogenic ESBL-producing bacteria, which might be transmitted to humans through direct contact or via the food chain (EFSA, 2011; Clemente et al., 2015; Elhariri et al., 2017b). The ESBL-producing and AmpC β-lactamase-producing Enterobacteriaceae have emerged globally (Ewers et al., 2012).
In all the examined Salmonella isolates from the investigated samples, the most detected ESBL genes were generally expressed phenotypically. This highlights the involvement of a specific resistance mechanism as estimated from the high existence of AmpC (63.7%), which is in accordance with the finding that this gene was more frequently present in Salmonella spp. isolated from chicken meat (Giuriatti et al., 2017). The second most prevalent gene bla SHV-1 (56.6%), followed by bla CMY-M2 (48.5%), bla TEM-1 (39.4%), and bla OXA-1 (27.3%), whereas bla CMY and bla PSE-1 genes were not detected. The other detected genes have a major role in resistance to third generation cephalosporins. Overall, there was a degree of correlation between the antibiogram phenotypic resistance pattern and the presence of resistance genes.
This antimicrobial resistance percent may result from extensive use or misuse of β-lactam group of antibiotics, which have been used for therapeutics of infectious diseases in animal feeds (Elhariri et al., 2017a). Our study has focused on the phenomenon of antibiotic resistance in S. Heidelberg isolates, which is concerned with the correlation between a resistance phenotype, common antibiotic resistance patterns, and the presence of the related genes, which is partially displayed in some cases. Therefore, although it is not always correlated with the resistance phenotype, the absence of a resistance gene does not necessarily indicate that S. Heidelberg is sensitive strain to the related antibiotic group of tested antibiotic gene.
The phylogenetic analysis of MDR strains of S. Heidelberg revealed low genetic diversity between isolates of different locality with different antibiotic patterns (Fig. 2). In general, human and avian MDR S. Heidelberg isolates cluster together, displaying a variable degree of diversity. For the three monophyletic groups, there was very low genetic diversity within the groups.
To estimate the potential virulence factors that control and enhance S. Heidelberg to cause infection in broilers and humans working in contact with chickens, PCR was used to investigate 11 virulence genes: invA, stn, sipC, rck, spvC, sopB, ssr, hilA, sipA, pefA, and ssaR. The majority of the examined broilers and human strains were positive to invA gene. The detected virulence genes were randomly distributed without constant profile even for human or broiler strains.
This is consistent with Retamal et al., 2015, who found that the most prevalent virulence genes found in S. Enteritidis strains isolated from poultry were invA (100%), pefA (93%), and spvC (93%) genes. Moreover, Han et al., 2013 evaluated putative virulence genes among S. Enteritidis strains from human and poultry production facilities and found that sopB, spvC, and invA genes (100%) are the most detected virulence genes. To our knowledge, there is a lack of the studies that have examined the prevalence of those genes in S. Heidelberg in Egypt.
Conclusion
The high prevalence of MDR-resistant Salmonella spp. in chickens and poultry workers in Egypt points to a great health risk in this part of the world, which has resulted in influential consequences in human health due to the high inclination for acquiring severe infections and productivity loss. All major resistance determinants, including those that confer resistance extended spectrum β-lactams, have been identified in various S. Heidelberg isolates. In addition, the current study suggests a role of animal-based food as a source of MDR-Salmonella demanding veterinary supervision on antibiotic use for therapy purposes in poultry. We suggest that substitutes such as probiotics and prebiotics to be tested against drug-resistant pathogens as intervention strategies.
To increase the current data concerning the epidemiological spreading of MDR strains between poultry and man, our study provides valued information correlated to circulation of MRD-resistant S. Heidelberg. This underscores the need for continuing surveillance of food-borne zoonotic pathogens along the food chain. It has become progressively clear that antibiotic resistance will remain a significant barrier to challenge in the nearby future.
Authors' Contributions
Mahmoud Elhariri and Salah Selim designed the study and contributed to the experimental work, data analysis, as well as the preparation of the article. Rehab Elhelw, Dalia Hamza, Mai Ibrahim, and Eman Hamza performed the experimental work. All authors read and approved the final article.
Ethics Approval and Consent to Participate
Oral consents were taken from workers before taking the stool samples.
Availability of Data and Material
All data generated or analyzed during this study are included in this published article.
Footnotes
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This study was carried out as a research activity without any funding or financial support.