
Abstract
Melons are perishable fruit of high food safety risk, grown in contact with soil and soil-borne organisms. To assess whether food safety risk could be augmented by the presence of soil-borne fungi, this study investigated the relationship between Fusarium spp. that were isolated from the surface of melon and the foodborne pathogen Salmonella enterica. In four repeated trials, rind discs from cultivars, Arava, Athena, Dulce Nectar, Jaune de Canaries, and Sivan fruit, grown in the field and in high tunnels in Maryland were inoculated separately with Fusarium isolates, F. oxysporum, F. fujikuroi, F. armeniacum, and F. proliferatum, with no Fusarium inoculation serving as a control and incubated at 25°C. Salmonella Newport was inoculated onto melon discs 4 d post-Fusarium inoculation and recovered 24 h later. Melon cultivar impacted the retrieval of Salmonella Newport. In all four replicated experiments, one or more of the netted varieties, Arava, Athena, and Sivan, yielded higher Salmonella Newport counts than one or both smooth-rind melons, Jaune de Canaries and Dulce Nectar (p < 0.05). Fusarium inoculation did not have a marked impact on Salmonella retrieval. The average Salmonella count recovered was 5.0 log colony-forming unit (CFU)/mL for both Fusarium-inoculated and uninoculated melons. However, in one trial, Salmonella Newport counts recovered from F. fujikuroi-inoculated melons were higher than all other treatments (8.6 log CFU/mL; p < 0.001), due to high levels of Salmonella recovered from Jaune de Canaries compared with other experiments. The food safety risk of melon did not appear to be enhanced by postharvest colonization with saprophytic Fusarium spp. However, melons with netted rinds appeared to favor Salmonella colonization compared with smooth melons. Choice of melon cultivar may be an important consideration in reducing Salmonella colonization risk in areas where Salmonella may be endemic in the environment.
Introduction
The growing consumption of fresh fruit and vegetables in the United States has coincided with substantial reports of foodborne illnesses associated with fresh produce (Lynch et al., 2009; Callejón et al., 2015). Salmonella enterica is frequently responsible for these foodborne illness outbreaks associated with produce in the cucurbit family, many involving melons (CDC, 2002, 2008, 2011, 2012; Angelo et al., 2015). Salmonella can adhere to melon fruit (Ukuku and Fett, 2002; Richards and Beuchat, 2004), while extracts from cantaloupe pulp and rind provide nutrients that support Salmonella growth and biofilm formation on food processing surfaces (De Abrew Abeysundara et al., 2018).
On fruit, Salmonella may benefit from fungal saprophytes or plant pathogens known to cause degradation of plant cell wall polymers and nutrient leakage (Aruscavage et al., 2008; Simko et al., 2015). Surveys conducted on produce reported an increased likelihood of detecting Salmonella when plant tissue was damaged by soft-rot pathogens compared with healthy plant tissue (Meng et al., 2013). The altered microenvironment in infected plant lesions caused by phytopathogens can allow for coinfection with enteropathogens. A study conducted on supermarket produce showed that Salmonella incidence was higher in the presence of Erwinia carotovora, a phytopathogen that causes soft rot on plants (Wells and Butterfield, 1997).
Many fungal saprophytes also grow on mature melons, even those that are harvested without wounding at the optimum harvest date (Fernández-Trujillo et al., 2007). Up to 25% of melon near-isogenic lines displayed Fusarium rot, following 30 d of storage (Fernández-Trujillo et al., 2007). Fusarium infection on cucurbit fruit results in mycelial growth that penetrates plant tissue, causing disintegration of the melon rind and underlying tissues forming sunken lesions and migrating into the center of the fruit leading to water-soaked areas (Seebold, 2010).
Melon infection may occur in the field before or during harvest, and symptoms may develop in storage, resulting in both preharvest and postharvest losses. However, since infection often proceeds and manifests after purchase by consumers, melons with less severe infection are frequently handled and may even be consumed. Weakly pathogenic or nonpathogenic Fusarium may also cause fungal rot because they are both prevalent on farms and excellent saprophytes.
Both Fusarium and Salmonella are endemic in the agricultural environment of the Eastern Shore of Maryland, a region known for cucurbit production. Our current study evaluated the impact of four Fusarium strains of the species F. oxysporum, F. fujikuroi, F. armeniacum, and F. proliferatum, isolated from melons grown in this region, on the fate of Salmonella Newport inoculated onto netted, honeydew, and Canary melons.
Materials and Methods
Microbial material
Fusarium spp. isolates for these experiments were obtained from a field survey on muskmelon grown in an organic field at the University of Maryland Lower Eastern Shore Research and Education Center (LESREC) during the summer 2015 and in a farmer's field in Delaware in summer 2016. Nine Fusarium isolates were isolated from melons with visible white or pinkish mycelia and identified based on translation elongation factors 1α and 2; Tef1—5′-ATGGGTAAGGAGGGACAAGAC-3′ and Tef2—5′-GGAAGTACCAGTGATCATGTT-3′ (O'Donnell et al., 1998), and RNA polymerase II second largest subunit (RPB2) (5′-GGGGWGAYCAGAAGAAGGC-3′) (O'Donnell et al., 2007) and RPB2-RP1 (5′-GGNGTCATGCARATCATNGC-3′) (LeBlanc et al., 2015). Four isolates identified as F. armeniacum, F2015007; F. oxysporum, F2015002; F. fujikuroi, F2015003; and F. proliferatum, F2016V016A (Table 1) were selected for the experiments, the latter used only in the fourth trial.
Fusarium spp. Isolates (F2015002, F2015003, F2015007) Isolated from Sivan F1 Certified Organic Melon Fruit During the Summer Season of 2015 and the Isolate F2016V016A from Ariel Fruit Obtained from a Conventional and Commercial Production Field in Laurel, Delaware During the Summer of 2016
All the four isolates used were identified using the Fusarium-ID database and NCBI Nucleotide database.
Pathogenicity test of Fusarium isolates
Pathogenicity was tested on store-bought honeydew melons, surface sterilized with 1.1% NaOCl, at the stem and blossom ends and in the center of the fruit, as previously described (Champaco et al., 1993). Fusarium oxysporum f.sp. niveum F063-2 (Zhou and Everts, 2007) and water were used as controls. Lesion depths of F2015003 and F2016V016A at the stem were 12.6 and 11.3 cm, respectively, less than, and the lesion at the blossom end caused by F2015003 was 22.6 cm wider than, the lesions caused by the nonpathogenic F. oxysporum control isolate. However, all other measurements were not significantly different from the control. These isolates were likely not pathogenic but saprophytic on the fruit surface.
The strain of Salmonella Newport used was a Virginia irrigation pond isolate identical to a tomato outbreak strain traced back to the same farm (Greene et al., 2008). The isolate had been previously adapted for rifampicin resistance.
Melon production
Five Cucumis melo L. cultivars were grown at LESREC in the field, in summer 2016 or in a high tunnel, a hoop house protected structure that allows extension of the growing season, in spring/early summer 2017; or in a high tunnel at the University of Maryland Wye Research and Education Center (WYEREC, Queenstown, MD) in fall 2016. Seed for cultivars representing different melon types: Arava (Cucumis melo var. reticulatus, Galia), Athena (var. reticulatus, muskmelon), Dulce Nectar (var. inodorus, honeydew), Jaune de Canaries (var. inodorus, Canary), and Sivan (var. cantalupensis, Charentais) were purchased from Johnson Seed Company (Salisbury, MD) and Eden Brothers (Fruitland, MD) in 2016 and 2017.
Seeds were sown in the greenhouse at LESREC before being hardened-off outside to prepare for planting, treated with Admire insecticide (ADMIRE® PRO Systemic Protectant, Bayer Crop Science LP, NC; 0.58 mL/L of water—29.6 mL/A) to prevent insect pest damage, then transplanted in the field, with spacings of 61 cm within and 183 cm between rows. Melon seedlings were started 3 weeks apart for two experiments in 2016. Melon cultivars, arranged in a randomized complete block design with 4 replicates for a total of 20 plots (3.0 × 1.8 m), were grown on raised bed/rows covered with 1.25 mm black plastic over a single line of a 20 cm-spaced emitter drip tape in a one-pass operation in the field. In the high tunnel, raised beds were covered in landscape fabric.
Fertigation was done using 1.1 kg/567.8 L of water of nitrogen (N), phosphorus (P), and potassium (K) (NPK ratio: 20:20:20) and applied using a water wheel transplanter. Melon foliar diseases and pests were managed with recommended fungicides, chlorothalonil (Bravo Weatherstik 6L Syngenta, Wilmington, DE), quinoxyfen (Quintec 2.08L; Dow Agro Sciences), and triflumizole (Procure 480SC; Arysta LifeScience). All fungicide applications were stopped 2 weeks (LESREC) and 4 weeks (WYEREC) before harvest. Fruit were harvested at the 3/4 slip stage (indicating the degree of detachment of stem from fruit) and kept at 4°C until processing.
Salmonella Newport and Fusarium spp. inoculum preparation
The Salmonella Newport strain was revived from frozen stock by growing at 37°C in Tryptic Soy Agar (TSA; BD, Franklin Lakes, NJ) amended with rifampicin (rif) (50 μg/mL) (Sigma-Aldrich, USA). Colonies were used to make Salmonella Newport suspensions in sterile 0.1% Peptone Water (PW) to OD600 = 0.5 equivalent to ∼109 colony-forming unit (CFU)/mL and diluted to ∼104 CFU/mL for melon rind disc inoculation. The culture was plated onto Tryptone Soy Broth + rif and incubated overnight for quantification.
Frozen Fusarium spp. isolates were streaked onto fresh Potato Dextrose Agar (PDA; Fisher Scientific, Fair Lawn, NJ) and incubated at 25°C for 10 d. To produce inoculum, conidial suspensions were prepared by transferring a fragment of sporulating mycelium to a sterile 10-mL test tube containing 0.1% PDA and vortexed to release conidia by filtering through sterile four-layered cheese cloth (ULINE, Pleasant Prairie, WI) and rinsed with sterile water into a beaker. The conidial concentration (spores/mL) was determined with a Hemocytometer (Fisher Scientific/Hausser Scientific Partnership 1475). Concentrations were adjusted to ca. 6 log10 spores/mL by preparing serial dilutions in sterile water containing 0.1% PDA.
Preparation of melon discs, inoculation of Fusarium spp. and Salmonella Newport, and Salmonella Newport enumeration
The experiments were conducted as a factorial design with five cultivar treatments and eight inoculation treatments for a total of forty experimental units. Melon fruit was washed with sterile water and a soft brush and transferred to a 1.1% household bleach (NaOCl) solution for 5 min for surface disinfestation, followed by rinsing in sterile water. After air drying, a biscuit cutter was used to cut eight melon rind discs (surface area = 158.5 cm2) per melon to be distributed uniformly to each of the eight treatments.
The inoculation treatments were inoculated with one of four Fusarium spp. plus Salmonella Newport. Experimental controls received no Fusarium (water only) plus Salmonella Newport. Treatments were replicated six times in the first, third, and fourth experimental runs and three times in the second run. Fruit inoculated with the four Fusarium spp. but not Salmonella Newport were evaluated as controls by reisolating Fusarium from the rind for inoculation on PDA and observation under 10 × magnification to confirm presence of Fusarium conidia. An additional control received no Fusarium or Salmonella inoculum to ensure no background Salmonella was present on melons.
Melon discs were placed in Whirlpak bags (Fisher Scientific) for inoculation with a 100 μL (20 μL in premarked 0.5 cm diameter circles) Fusarium spp. inoculum (6 log spores/mL) or water as a control. A shallow wound was made on the discs using a sterile pipette and the Fusarium inoculum applied to the wound. Disks were incubated at 25°C under 12-h light/12-h darkness for 4 d, followed by surface inoculation with 104 CFU/mL Salmonella Newport (20 μL per circle, a total of 100 μL total per disc) and incubated at 25°C for 24 h under the same conditions. Incubation conditions were selected to achieve a final 5-d incubation for Fusarium and 24 h incubation for Salmonella as previously determined for microbial establishment (Ukuku et al., 2006; Sui et al., 2014). Disks were washed in 0.1% PW in the same bag by hand rubbing for 2 min then wash used to prepared serial dilutions to be plated onto TSA + rif and cycloheximide (50 μg/mL; Sigma-Aldrich) for bacterial enumeration after incubation at 37°C for 24 h.
Data analysis
Differences in levels of recovered Salmonella Newport coinoculated with Fusarium on five melon cultivars were statistically assessed in JMP Pro 14.10.0. using a mixed model with response being bacterial counts and model effects being Fusarium treatment, melon cultivar and Fusarium treatment × cultivar and Tukey–Kramer honestly significant difference (HSD) was used to compare the differences among means when the overall effect was significant. CFU/mL were log transformed before analysis and α-level was set at 0.05.
Results
Impact of melon cultivar on Salmonella Newport
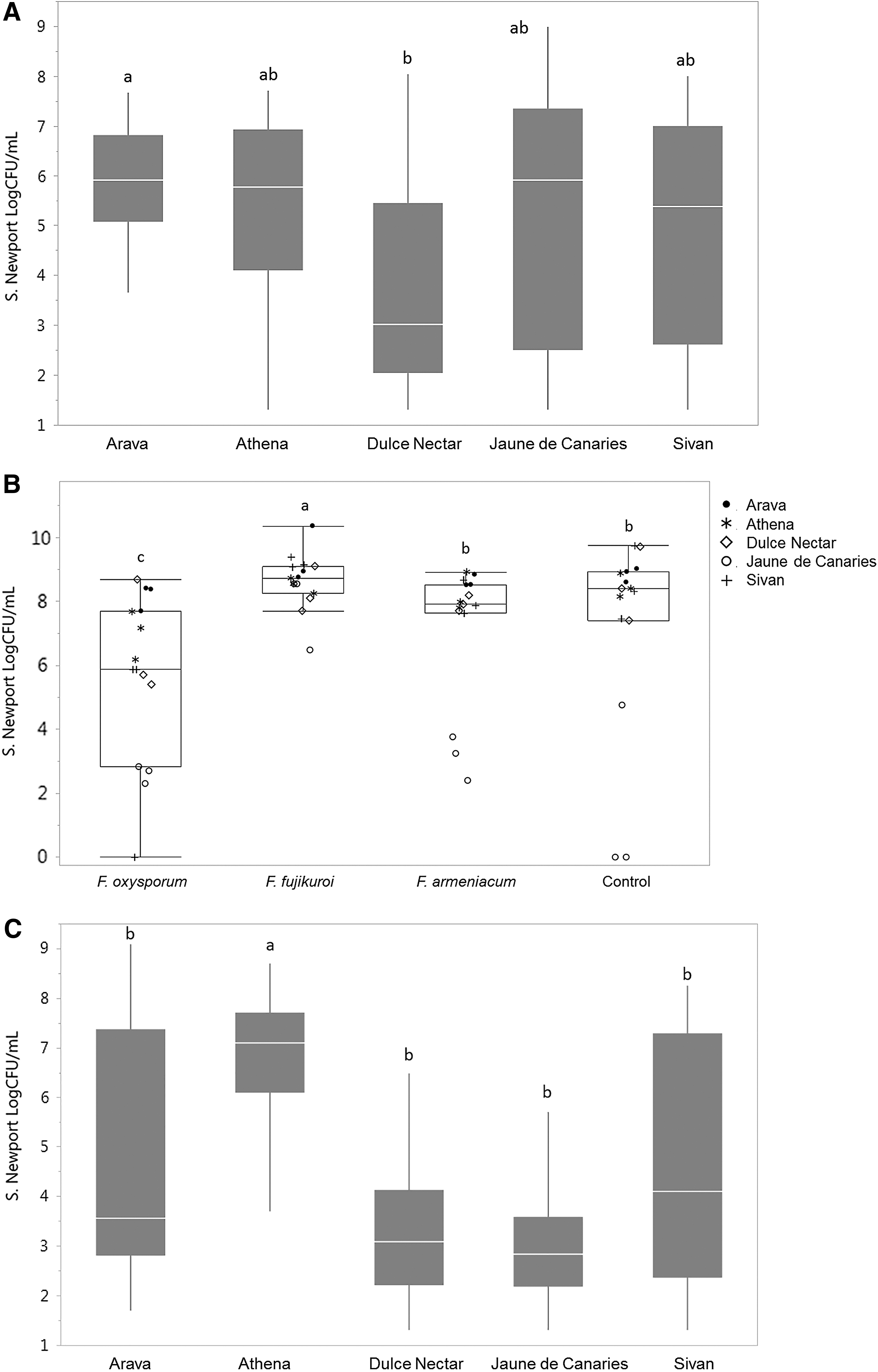
Melon cultivar consistently impacted the levels of Salmonella Newport recovered from melons. Growth of Salmonella Newport was detected on several melon samples and on all samples of Athena, Arava, and Sivan (i.e., retrieved counts were higher than original inoculum). Overall, Athena, Arava, and Sivan (mean counts of 5.9, 5.8, and 5.3 log CFU/mL, respectively) were more favorable to Salmonella Newport colonization than Jaune de Canaries and Dulce Nectar (mean counts of 4.1 and 3.9 log CFU/mL, respectively). The cultivar effect varied slightly among experiments, but this trend was consistent. Arava supported significantly higher counts than Dulce Nectar on the first set of field-grown melons (5.8 and 3.7 log CFU/mL, respectively; p < 0.01; Table 2; Fig. 1A). In the second set of field-grown melons, Jaune de Canaries yielded the lowest Salmonella, with a count at 3.8 log CFU/mL, which was significantly different from all other cultivars (p < 0.001; Table 2; Fig. 1B).
Population levels of Salmonella Newport retrieved from melon rinds of five different cultivars inoculated with Fusarium oxysporum, Fusarium fujikuroi, Fusarium armeniacum, or uninoculated (water control).
Mixed Model Results for the Four Experimental Runs
Field-grown melons (1) early summer, (2) late summer, 2016 and high tunnel-grown melons (1) late fall 2016, (2) late spring, 2017 showing impact of coinoculation with Fusarium spp., melon cultivar and interaction between the two factors on Salmonella Newport population levels retrieved from melon rinds (Fusarium spp.: n = 30 for field 1, high tunnel 1 and 2; n = 15 for field 2; Cultivar n = 24 for field 1, high tunnel 1 and 2; n = 12 for field 2). Asterisks denote significance at α = 0.05.
df, degrees of freedom; Prob, probability.
Dulce Nectar on the other hand was comparable to Arava, Sivan, and Athena at 7.8 log CFU/mL. In the experiment with the first set of high tunnel-grown melons, Salmonella Newport counts from Athena (6.6 log CFU/mL) were significantly higher than all other cultivars, with a ≥3 log CFU/mL difference between this cultivar and Jaune de Canaries and Dulce Nectar (3.3 and 3.6 log CFU/mL, respectively; p < 0.001; Fig. 1C). In the experiment with the second group of high tunnel-grown melons, counts from Arava, Sivan, and Athena significantly exceeded (by 1.4–2.5 log CFU/mL) counts from Jaune de Canaries and Dulce Nectar (p < 0.05; Fig. 2A).

Box plots displaying population levels of Salmonella Newport retrieved from melon rind of five different cultivars inoculated with Fusarium fujikuroi, Fusarium oxysporum, Fusarium proliferatum, or uninoculated (water control).
Grouping melons by rind type; netted (Athena, Arava, and Sivan), honeydew (Dulce Nectar), and Canary (Jaune de Canaries), revealed a significant difference between netted and smooth rind types. The average Salmonella count enumerated from netted types was 5.7 log CFU/mL, compared with 4.1 and 3.9 log CFU/mL counted on honeydew and Canary melons, respectively (p < 0.001).
Impact of Fusarium spp. on Salmonella Newport on melon
Twenty-four hours after Salmonella inoculation, Salmonella Newport was recovered from all melon rinds assayed (N = 420), except for one Sivan sample inoculated with F. oxysporum and two control Jaune de Canaries samples. Salmonella was not recovered from no Salmonella control melons, that is, inoculated with Fusarium only or water only (data not shown). Overall, no significant difference was detected in Salmonella counts recovered from melons inoculated with Fusarium and uninoculated (no Fusarium) melons.
The average Salmonella counts recovered were 5.0 log CFU/mL for both melons inoculated with Fusarium and Fusarium-uninoculated melons. When each run was assessed separately, to account for any difference in melon ripeness among harvests, Fusarium inoculation significantly affected Salmonella Newport in the second experiment with field-grown melons (field 2) and the second experiment with high tunnel-grown melons (high tunnel 2; Table 2).
In field 2 melons, Salmonella Newport counts recovered from F. oxysporum-treated melons were significantly lower (5.7 log CFU/mL) compared with melons with F. armeniacum and water control (both at 7.2 log CFU/mL; p < 0.01), whereas Salmonella Newport counts on F. fujikuroi-treated melons were significantly higher than on all other treatments (8.6 log CFU/mL; p < 0.001; Fig. 1B). The increase was attributed to high levels of Salmonella recovered from Jaune de Canaries melons infected with F. fujikuroi, which was anomalous for this cultivar and attributed to a more advanced ripeness stage.
By contrast, in the second experiment with high tunnel-grown melons, F. fujikuroi-inoculated melons yielded 0.9–1.1 log lower Salmonella counts than the water control (p < 0.05) and other Fusarium treatments (not significant) (Fig. 2B).
Interaction between Fusarium spp. and Salmonella
Overall, no interaction was detected between Fusarium spp. and cultivar or melon type (netted vs. smooth types). Assessing melon harvests separately, however, uncovered a significant interaction in the second field experiment (p < 0.001; Table 2). This was attributed to the cultivar Jaune de Canaries which, as explained above, when inoculated with F. oxysporum and F. armeniacum, or left uninoculated, yielded significantly lower Salmonella counts than other cultivars (p < 0.001), in contrast to inoculation with F. fujikuroi. (Fig. 1B). This effect was not reproduced in other experiments. Similarly, an interaction was also seen with Sivan melons inoculated with F. oxysporum, where significantly lower Salmonella counts were recovered (p < 0.05) than from other Sivan treatments and other cultivars, excluding the Jaune de Canaries treatments that were also significantly lower (Fig. 1B).
Discussion
In this study, the significant role of melon type on the ability of Salmonella to colonize melon surfaces was clearly demonstrated. The enhanced recovery of Salmonella Newport from cultivars with netted rinds, compared with the smooth honeydew and Canary types assessed, was consistent and reproducible. Surface roughness of exocarps enhances bacterial attachment to fruit surfaces (Wang et al., 2009; Fernandes et al., 2014). The extremely rough surface provided by the melon net of Athena, Arava, and Sivan fruit compared with the much smoother honeydew and Canary cultivars used could explain this difference. The increased surface area itself could also contribute to augmented bacterial numbers.
Bacterial surface attachment to fruit however is not only dependent on surface roughness, but also surface hydrophobicity and electric charge of both fruit and bacterial surfaces (Ukuku and Fett, 2002, 2006; Fernandes et al., 2014). In addition, among foodborne pathogens, Salmonella may be particularly adapted to adhere to fruit, exhibiting the highest surface hydrophobicity and attachment to cantaloupe (Ukuku and Fett, 2002). However, Salmonella Typhimurium did not exhibit differences in attachment to tomato, considered to be hydrophobic compared with hydrophilic mango surfaces in a study that quantitatively measured the free energy of interaction to assess hydrophobicity (Fernandes et al., 2014). Therefore, even if differences exist in the surface hydrophobicity of various melon fruit, a characteristic we did not measure, the microbiological traits facilitating adhesion of Salmonella Newport could supersede any repulsion conferred by fruit hydrophobicity.
Fernández-Trujillo et al. (2007), observed that skin netting was correlated to cracking and increased fruit decay. This likely also leads to nutrient leakage, which could favor Salmonella Newport growth. Regardless of mechanism, fissures and irregularities in the fruit have been associated not only with enhanced enteropathogen attachment, but also reduced efficacy of aqueous sanitizer treatments (Bastos et al., 2005; Wang et al., 2009; Fernandes et al., 2014).
Findings from this study demonstrated that Salmonella Newport can grow in the presence of Fusarium spp. present on melon fruit, but the fungal saprophytes did not affect survival or growth of Salmonella Newport. In almost all cases, the water (no Fusarium) controls yielded equivalent population levels of Salmonella to Fusarium-inoculated samples, regardless of Fusarium spp. Noncooperative relationships between fungi or bacteria and human pathogens have been reported. Although coinoculation of Salmonella with Erwinia tracheiphila on cantaloupe rind increased the incidence of Salmonella-positive samples, cohabitation with the phytopathogen did not benefit Salmonella, which declined over time (Gautam et al., 2014). Coinoculation of fungal pathogens in the genera Alternaria, Cladosporium, Geotrichum, Epicoccum, and Penicillium with Salmonella Poona onto intact cantaloupe rind also had no effect on the enteropathogen, although previous wounding provided some benefit to Salmonella growth at 4°C but not 20°C for all fungi, except Pencillium expansum (Richards and Beuchat, 2005).
In this study, the Fusarium spp. isolates used were collected from the same region where melons were grown and all species were determined to be saprophytic on honeydew melon. This scenario would approach the kind of organism that may be present on store-bought melons as severely infected fruit would not be marketable. The saprophytic or necrotrophic nature of these Fusarium isolates could explain the minimal influence on Salmonella Newport we observed.
Although we generally saw no effect of Fusarium spp. on melons, in one experimental trial using field-grown melons we retrieved higher levels of Salmonella from Canary melons inoculated with F. fujikuroi. This result was inconsistent with all other data collected and is attributed to the late-season crop (September) and the more advanced ripeness of this cultivar at the time of sampling. It is possible that the softer rind and higher sugar levels present on riper fruit could have facilitated Salmonella growth on a cultivar that in general exhibited the lowest propensity for Salmonella proliferation.
Conclusion
The food safety risk of melon did not appear to be enhanced by postharvest coinoculation with Fusarium spp. Choice of melon cultivar, however, may be an effective strategy in reducing Salmonella colonization risk in areas where Salmonella may be endemic in the environment and the inherent risk is deemed higher (e.g., areas of hot, humid climates or where Salmonella is detected in surface waters or the environment, etc.). Assessing risk of any fruit- and vegetable-growing area should be part of a farm food safety plan; production of smooth rind melons such as honeydew and Canary types over netted types should be considered as a mitigating strategy in higher risk areas.
Footnotes
Acknowledgments
The authors sincerely thank Yacintha Johnson and Anthony LaBarck for laboratory assistance, and David Armentrout and Michael Newell for field management.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the U.S. Department of Agriculture's National Institute of Food and Agriculture (USDA-NIFA) through the Organic Transitions Program, grant 2014-51106-22090. The sponsor played no role in the study design and implementation. Any opinions, findings, and conclusions expressed in this material are those of the authors and do not necessarily reflect the views of the USDA-NIFA.