
Abstract
This study investigated the inactivation mechanism of Aspergillus flavus conidia by high hydrostatic pressure (HHP). Activity counts, scanning electron microscopic (SEM) analysis, and sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) were used to study the effects of the HHP treatment on the morphology and protein composition of A. flavus spores. The results showed that that a 3-min-lasting 600 MPa treatment could completely abolish 107 colony-forming units/mL of live fungi. Furthermore, we also observed that lower spore viability corresponded to a higher Propidium Iodide absorption rate. The SEM images revealed that HHP disrupted the spore morphology and resulted in pore formation that led to the release of intracellular molecules, such as nucleic acids and proteins. The nucleic acid and protein concentration in the spore suspension increased in parallel with the increasing treatment pressure. The SDS-PAGE analysis showed that there were differences in the protein bands between the HHP-treated and untreated A. flavus spores, as the HHP treatment caused partial protein degradation and extracellular release. Therefore, the results of this study proved that high pressure could induce a morphological disruption in the internal and external cellular structures and degrade intracellular and extracellular proteins leading to an inactive state in A. flavus.
Introduction
The principle of high hydrostatic pressure (HHP) includes placing a sealed food packaging in a closed and insulated container, then applying an extremely high pressure of 100–600 MPa in a liquid (usually water) transmission medium to inactivate food pathogenic microorganisms at room temperature (Wang et al., 2016). In addition to the spore-forming microorganisms, high-pressure processing also has good pasteurization effects on most other microorganisms. However, as the physiological characteristics of the various microorganisms also vary, their pressure resistance might also be different. Microorganisms exhibit strong environmental adaptability and employ different mechanisms for self-protection against adverse environments (Huang et al., 2014a). Examples include domestication to adapt to the environment, conversion to a dormant state (spore), activating pressure resistance genes, or producing adaptive mutations. Therefore, pressure resistance in microorganisms is not fixed and is often affected by several internal and external factors, such as the type of microorganism, growth period, and environmental factors. In an environment suitable for growth, microorganisms would have better pressure resistance compared with suboptimal environmental conditions for microbial growth (Georget et al., 2015).
As the processing pressure increases, different effects could be observed in the microorganisms. According to previous studies, a 50 MPa pressure inhibited protein synthesis and reduced the number of ribosomes in the microorganisms, 100 MPa led to partial protein denaturation, 200 MPa disrupted the cell membrane and intracellular structure, and 300 MPa resulted in irreversible changes in the enzymes and proteins, and ruptured the cell membrane, causing a leakage of the intracellular substances and leading to subsequent microbial death (Syed et al., 2016). Therefore, the high-pressure-induced physical damage of the microorganisms could be observed through different analytical tools. Ma et al. (2019) applied scanning electron microscopy (SEM) and transmission electron microscopy (TEM) to observe the morphological changes in Escherichia coli O157:H7 under high-pressure processing. Their results revealed that significant intra- and extracellular damage occurred after exposure to 400 MPa for 5 min. Most cells shrunk when under pressure, while some became oval shaped or got crushed, and wrinkles or depressions appeared on their surface. Yang et al. (2012) used TEM to observe E. coli and Staphylococcus aureus under HPP. They showed that when the microbes were subjected to a 500 MPa pressure, the cells ruptured and the peptidoglycan layer degraded. At the same time, pulsed-gel electrophoresis showed that high-pressure processing resulted in the breaking of the DNA bonds in bacteria. In addition, protein electrophoresis showed that high-pressure processing led to protein degradation in Salmonella, and the expression level of some proteins decreased significantly with increasing pressure. The application of 300 MPa pressure led to the complete degradation of protein bands at a molecular weight of 48 kDa (Wang et al., 2013). The aforementioned study described that high pressure could cause significant damage in the external morphology and intracellular structures of bacteria and inhibit intracellular protein expression. These are the possible mechanisms by which high-pressure treatment could lead to cell death (Baptista et al., 2016).
Microbial contamination by A. flavus is a major problem in the case of agricultural raw materials. In particular, A. flavus proliferates rapidly at 30°C in a moist environment, leading to food material decay with subsequent production of aflatoxin, a toxic metabolite. Aflatoxin is extremely toxic and exhibits high hepatotoxicity, carcinogenicity, and immunosuppression. It is listed as a Group I carcinogen by the World Health Organization. Peanuts, maize, rice, wheat, beans, nuts, and Chinese herbs might be infected by aflatoxin-producing fungi (Bediako et al., 2019). This is especially true for processed peanut products, and aflatoxin-contaminated peanuts could no longer be consumed by humans, causing significant economic loss to the peanut industry. Aflatoxin contamination is a major problem in the global food and feed industry. Therefore, there is a growing interest among the researchers to identify effective treatment methods to maintain food quality and inactivate Aspergillus species, thus retaining food purity. Our previous study described that HHP could inhibit the growth of A. flavus in peanuts and indirectly decrease aflatoxin accumulation in peanuts (Huang et al., 2014b). However, the inactivation mechanisms of HHP on A. flavus conidia have not yet been fully elucidated. Therefore, the aim of this study was to observe the effects of HHP on the viable A. flavus counts, cell morphological disruption, and protein changes.
Materials and Methods
Preparation of the fungal suspension
Aspergillus flavus BCRC 30003 was purchased from the Food Industry Research and Development Institute, HsinChu, Taiwan. The fungus was cultured on potato dextrose agar at 25°C for 3 d until the surface was full of spores. The spores were then collected. The spores were added to sterile distilled water and serially diluted to prepare a spore suspension of 107 spores/mL. The suspension was stored at 4°C for subsequent experiments. The spore suspension (10 mL) was added to a sterile vacuum bag. After vacuum sealing, using a vacuum packaging machine (CVS3050; Artisan, Taiwan), the package was immediately processed for high-pressure treatment.
HHP treatment
The volume of the high-pressure device used in this study was 6 L (Baotou Kefa High-Pressure Technology Co. Ltd. Baotou city, Inner Mongolia Autonomous Region, China), and the maximal applied operating pressure was 600 MPa. Water was used as a transmission medium for high-pressure treatment and the pressure increase rate was ∼300 MPa/min. Deionized water was used as a transmission medium at an initial temperature of 10°C, then the operating temperature increased by 3°C/100 MPa. After the high-pressure treatment, the pressure was released within 10 s. The high-pressure treatment duration in this study did not include the time taken to increase or decrease pressure. The vacuum-sealed spore suspension was placed inside the high-pressure chamber and treated at 100, 200, 300, 400, 500, and 600 MPa pressure for 1, 3, and 5 min. The vacuum bags were immediately placed in an ice bath after the high-pressure treatment before subsequent experiments and analyses.
Enumeration of fungal counts
The high-pressure-treated spore suspensions were serially diluted with 0.1% buffer peptone water, and 0.1 mL of the diluted suspensions were plated on a 3M Petrifilm™ plate for yeasts and molds, then incubated at 25°C for 120 h. Plates with colony counts of 25–250 were used for further analysis, and the effects of high pressure on colony counts were recorded. The residual colony counts under different treatment conditions were compared. The results of the triplicate experiments were expressed as mean ± standard deviation (SD).
Scanning electron microscopy
The unprocessed and high-pressure-processed spore suspensions were centrifuged at 10,000 × g for 10 min and 0.1 M phosphate buffer (pH = 7.4) was used to wash the pellet twice. The pellet was then resuspended in 1 mL of 0.1 M phosphate buffer (pH = 7.4). After filtration (0.22 μm MF-Millipore, GSWP), the cell suspension was fixed on a membrane with 10 mL of 1.0% glutaraldehyde/0.1 M phosphate buffer (pH = 7.4). The cells were incubated with the fixing solution at 4°C overnight, and the membrane was transferred to a glass bottle and washed with buffer thrice (10 min per wash). Next, 1% osmium tetroxide (OsO4) was used for 1 h to fix the cells. The membrane was washed with the buffer twice (10 min per wash). The membrane was then treated with a series of 10 mL ethanol solutions (10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, and 100% ethanol) for 15 min each, for dehydration. The sample was first immersed in isopentyl acetate, in a carbon dioxide medium before critical point drying using a critical point dryer (Hitachi HCP-2; Hitachi High Technologies America, Inc., Dallas, TX). Then the sample was pasted to the SEM stage using the double-sided tape and gold foil was wrapped around it. A Hitachi S-4700 SEM (Hitachi High Technologies America, Inc.) was used to observe the samples at an operating voltage of 15 kV.
Nucleic acid and protein content analysis of the spore suspension
The unprocessed and high-pressure-processed spore suspensions were centrifuged at 5000 × g and 4°C for 10 min, and the supernatant was collected for nucleic acid and protein quantification in the spore suspension. We used a microplate reader (Spectrostar Nano; BMG Labtech) to measure the absorbance at 260 and 280 nm for the nucleic acid content. In addition, the protein concentration was also analyzed using a commercial Bio-Rad Protein Assay Kit: 20 μL of the protein sample was allowed to react with 1 mL of protein dye reagent (Bio-Rad) in dark for 15 min, then a microplate reader (Spectrostar Nano; BMG Labtech) was used to measure the absorbance at 595 nm. The protein content was then calculated based on a standard curve based on different bovine serum albumin concentrations.
Protein spectral analysis
The unprocessed and high-pressure-processed spore suspensions were centrifuged at 5000 × g and 4°C for 10 min, then spore pellet was collected and processed for protein extraction using the Minute™ Total Protein Extraction Kit for Microbes (Inventbiotech) according to the manufacturer's instructions. Briefly, 20 μL of denaturing buffer was added to 80 mg of pellet and mixed evenly, then 200 μL protein extraction buffer was added to the solution and mixed evenly. The solution was then vortexed vigorously for 10 s. Next, the solution was centrifuged at 5000 × g and 4°C for 3 min, then the protein-containing supernatant was collected for protein concentration analysis followed by an sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis. We analyzed the distribution of the A. flavus protein bands before and after high-pressure treatment. A 0.75-mm-thick discontinuous acrylamide gel was used, comprising of a 4% stacking and a 12% separating gel. The polymerized gel was placed in an electrophoresis tank and supplemented with running buffer (25 mM Tris, 192 mM glycine, and 0.1% SDS). The protein sample was mixed with 3 × SDS-PAGE Loading Buffer (6% SDS, 0.3% bromophenol blue, 30% glycerol, 150 mM Tris-HCl pH = 6.8, and 300 mM DTT) and heated at 100°C for 5 min before placing on ice. The samples were added to the wells in the gel and ran at a voltage of 70 V. After samples passed through the stacking gel (∼15 min), the voltage was adjusted to 140 V, and the electrophoresis duration to ∼45–50 min. The gel was stained with a staining solution (0.125% Coomassie Blue, 50% methanol, and 10% acetic acid) and destained with a destaining solution (methanol:acetic acid:water; 2:1:7) before the protein spectrum was observed.
Effects of HHP on cell membrane integrity
The method of Tian et al. (2012) was used as a reference, and Propidium Iodide (PI) to stain the nucleic acids. The PI staining was carried out to understand the effects of HHP on A. flavus spore membrane integrity. Briefly, 10 mL of fungal spore suspension (1 × 107 spores/mL) was vacuum packaged and exposed to different pressures (100–600 MPa) at 25°C for 5 min. The fungal spore suspension that did not undergo HHP was used as a control. Finally, the spore suspension was stained with 5 μM PI for 15 min, followed by centrifugation (10,000 × g) at 4°C, and washed with phosphate-buffered saline twice. The fluorescence absorbance was measured at an excitation wavelength of 490 nm and an emission wavelength of 635 nm using a microplate reader (Synergy H1; BioTek Instruments, Inc.). The absorbance at 635 nm was expressed as a percentage and the residual fungal count after the pressure treatment was used for correlation comparison.
Statistical analysis
The experimental results represent the mean of triplicate experiments. The data are presented as the mean ± SD, and they were analyzed using a statistical analysis system (SAS, Inc.), conducting one-way analysis of variance. The results were considered statistically significant at p < 0.05.
Results and Discussion
High-pressure treatment
After the A. flavus spores underwent varying degrees of high-pressure treatment, the differences in the combination of time and pressure resulted in significant viability differences. When 100 MPa pressure was applied, all the treatment durations decreased by only 1 log of fungal counts (Fig. 1). When we applied a pressure beyond 300 MPa, the colony counts in the various treatment groups showed a significant reduction. When treatment pressure was as high as 500 MPa and the duration was 1, 3, and 5 min, the fungal count decreased by 3.4, 4.8, and 6.3 log cycles, respectively, showing that the treatment at 500 MPa for 5 min was sufficient to lower the fungal count below the detection limit. When a 600 MPa pressure was applied, only the treatments lasting for at least 3 min could completely inhibit the A. flavus spore activity, resulting in fungal counts dropping below the detection limit, while 1 min of treatment resulted in 2.9 log of the residual fungal count. Thus, these results reveal that a pressure beyond 500 MPa and suitable treatment durations could effectively inactivate A. flavus. Similar results were reported by Pinto et al. (2020), who indicated that fungal ascospores under mild hydrostatic pressures are suitable to trigger the necessary germination pathways to surpass the dormancy, whereas the pressures over 500 to 600 MPa, wherein the ascospores are generally inactivated. Recently, many studies have pointed out that high-pressure pasteurization could prevent contamination by toxigenic fungi in food. Kalagatur et al. (2018) pointed out that treatment at 45°C and 550 MPa for 20 min could effectively inhibit the growth of Fusarium graminearum and reduce the levels of fungal toxins such as deoxynivalenol and zearalenone. Huang et al. (2014b) inoculated A. flavus on peanuts, carried out varying degrees of high-pressure processing, and found that pressures equal to or below 400 MPa do not significantly inhibit A. flavus, with the fungal counts returning to the levels before high-pressure pasteurization in 10 d. However, a pressure of 800 MPa resulted in the maintenance of A. flavus counts at the levels below the detection limit within a 30-day storage period, indirectly reducing aflatoxin accumulation in peanuts. Groot et al. (2019) applied high-pressure processing to reduce Penicillium counts in orange juice. Their results showed that a 400 MPa treatment for 5 min could reduce Penicillium counts in orange juice by 7 log. Penicillium bialowiezense spores were found to be the most sensitive to the HHP treatment, as a 1-min-lasting treatment at 400 MPa reduced the fungal count by 6 log. Chen et al. (2018) achieved the growth inhibition of ochratoxin-producing Aspergillus fresenii on coffee beans by treating them at 600 MPa for 5 min. In the 50-day storage experiment, A. fresenii started to grow on day 10 upon a 200 MPa-pressure treatment or below, while A. fresenii growth was inhibited in the coffee beans treated with a pressure of 600 MPa up to day 50.
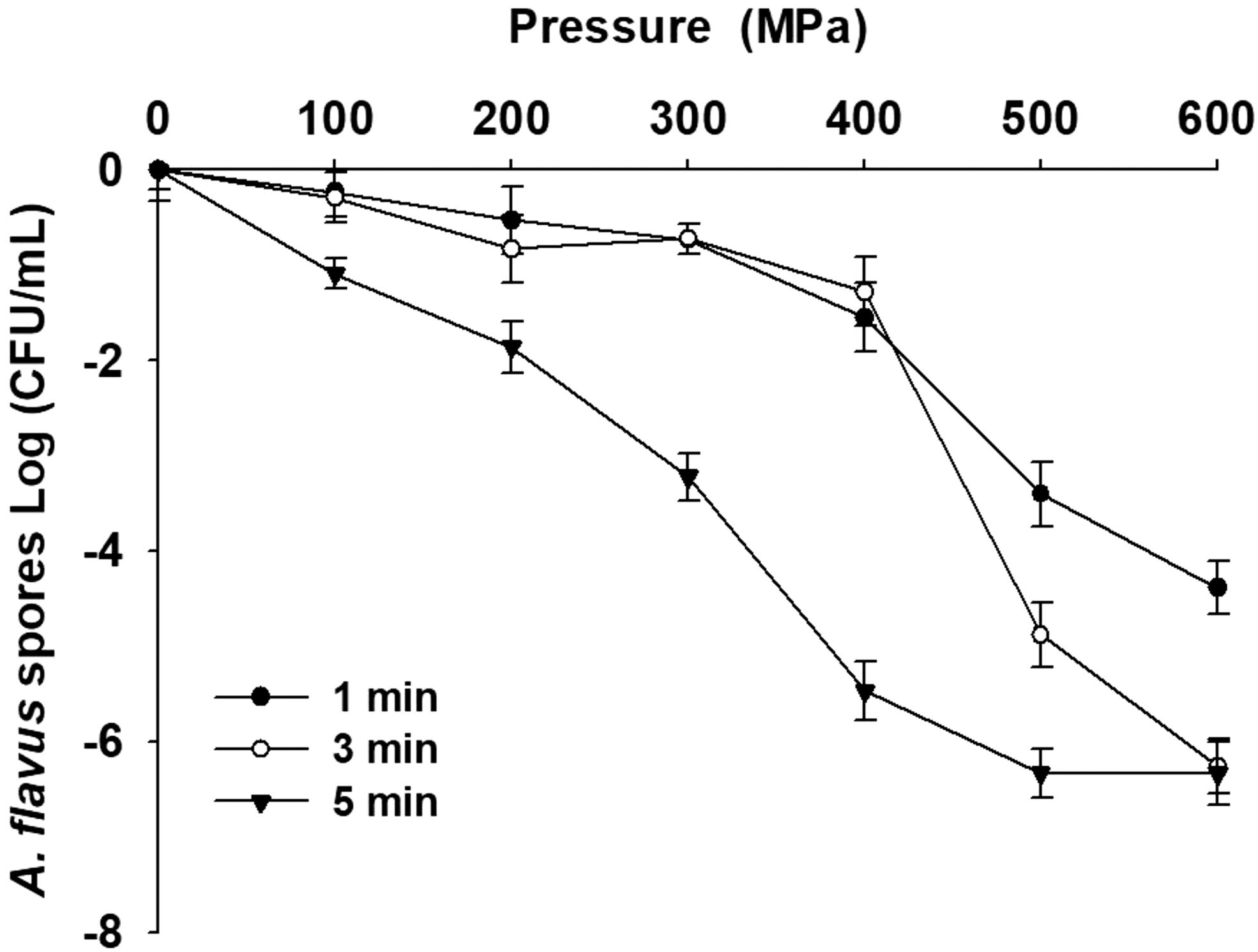
Aspergillus flavus loss of viability after 1, 3, or 5-min-lasting treatments at different pressures. The plotted values are the mean of three replicate trials ± standard deviation.
Scanning electron microscopy
Electron microscopes are the most direct tools for observing the morphological and structural changes in the microorganisms. In this experiment, we compared the physical damage on A. flavus spores before and after high-pressure treatment through electron microscopic observations. Figure 2 shows the scanning electron micrograph of A. flavus spore morphology with or without the HHP treatment. From Figure 2a, it could be seen that A. flavus spores in the untreated group appeared to be spherical and were densely attached to the hyphae. Figure 2b shows the spores after a 600 MPa HHP, where some spores look irregular, collapse, and are distorted after the compression with pores appearing on their surface. In Figure 2c, no pores were observed on the 15,000-times-magnified spores, appearing as saturated spheres. Figure 2d shows that the HHP-treated spores were compressed with an appearance of hollow pores on their surface. The contents of the spores leaked out through these pores while the spores still appeared spherical. From the above-described results, it was clear that the HHP treatment led to physical damage of the A. flavus spores but did not rupture them. Instead, pores appeared on the spore surface, causing an internal content leak. Similar results were reported by Rozali et al. (2017), who evaluated the impact of high pressure (600 MPa for 5 min) on the ultrastructure of Neosartorya fischeri ascospores, the SEM picture revealed that high-pressure treatment tore the membranes apart, with evident leakage of inner cell content. Dasan et al. (2017) also used SEM to observe the morphological changes in A. flavus and A. parasiticus spores before and after nonthermal plasma treatment, and found that similar to the HHP-treated A. flavus spores, nonthermal plasma treatment also resulted in pore formation on the A. flavus spores, causing an intracellular content leak. However, nonthermal plasma caused the spores to detach from the hyphae. As the high-pressure treatment was carried out within a sealed container and was transmitted uniformly by water to the spores, there was no significant change in the appearance with changes observed only in the microstructure in our present study. Therefore, the spores were still attached to the hyphae after the HHP. This observation was similar to the results of previous studies showing that HHP treatment resulted in a cell wall and cell membrane damage, leading to disrupted cell permeability, resulting in morphological and structural changes. The ascospores of Talaromyces macrosporus were observed by SEM after treatment at 600 MPa for 10 s and 60 min, respectively, and the results showed that pressure treatments resulted in indentations/bending of the cell wall ornaments, which were more noticeable for prolonged holding times. Prolonged holding times (60 min) also resulted in quicker formation of hyphae than 10 s, with this thought to be related to the damages induced by longer pressure exposure (Dijksterhuis and Teunissen, 2004). Reyns et al. (2003) revealed that high-pressure treatment induced a collapse of ascospores of Talaromyces macrosporus, by sensitized cell wall lytic enzymes and loss of spore wall barrier properties.

Scanning electron micrographs of Aspergillus flavus spores.
Analysis of nucleic acid and protein content in A. flavus spore suspension
Figure 3 shows the nucleic acid and protein content released by the A. flavus spores into the buffer after varying degrees of HHP. In the control group, the absorbance at 260 nm was greater than at 280 nm for the supernatant of A. flavus spore suspension, and no proteins could be detected after the Bradford assay. The 100 MPa treatment resulted in a significant absorbance increase both at 260 and 280 nm. Moreover, small amounts of protein (22.7 μg/mL) could be detected in the supernatant. As the extent of the treatment pressure increased, the absorbance values at 260 and 280 nm also increased significantly. The maximum absorbance at 260 nm (1.72) was recorded after treatment at 600 MPa for 5 min, while a maximum absorbance at 280 nm occurred in the 500 MPa treatment group (1.24). We also observed a similar trend to the absorbance at 280 nm during the Bradford protein assay to quantify the protein concentration in the supernatants of the A. flavus spore suspension, that is, increasing protein content in the supernatant with increasing pressure. Following the 400 MPa treatment, the protein concentration was 117.2 μg/mL. Although there was a slight increase in the protein concentration when pressure increased to 600 MPa, no significant difference could be observed compared with the 400 MPa-treated samples. This result was similar to those of previous studies. Usually, the cell membrane damage increases with increasing pressure, and lethality is also higher, resulting in the secretion of multiple intracellular components. HHP would also affect the function of the genetic material in the organisms. Studies have shown that high pressure would inhibit the enzyme activity during DNA replication, transcription, and the condensation of the genetic material, causing the DNA to interact with nucleic acid enzymes and in the process, the DNA would be cleaved, causing chromosomal DNA degradation. Microorganisms possess regulatory genes to adapt to the changing environment. When microorganisms are in an environment that is not suitable for their growth, they would increase their resistance against the adverse environments by intracellular accumulation of large amounts of heat shock proteins, a classical form of genetic control that could strengthen bacterial resistance facing several types of environmental stress. Yang et al. (2012) employed pulsed-field gel electrophoresis to observe the effects of high-pressure processing on E. coli and Shigella dysenteriae DNA. The results from that study showed that high pressure resulted in the DNA bond breaking. Interestingly, Mohamed et al. (2012) found different results. Listeria monocytogenes underwent high pressure, gamma radiation, and pulsed electric field treatment. Their DNA fragment analysis showed that only gamma radiation resulted in DNA lesions, whereas the high pressure and pulsed electric fields did not affect the DNA.
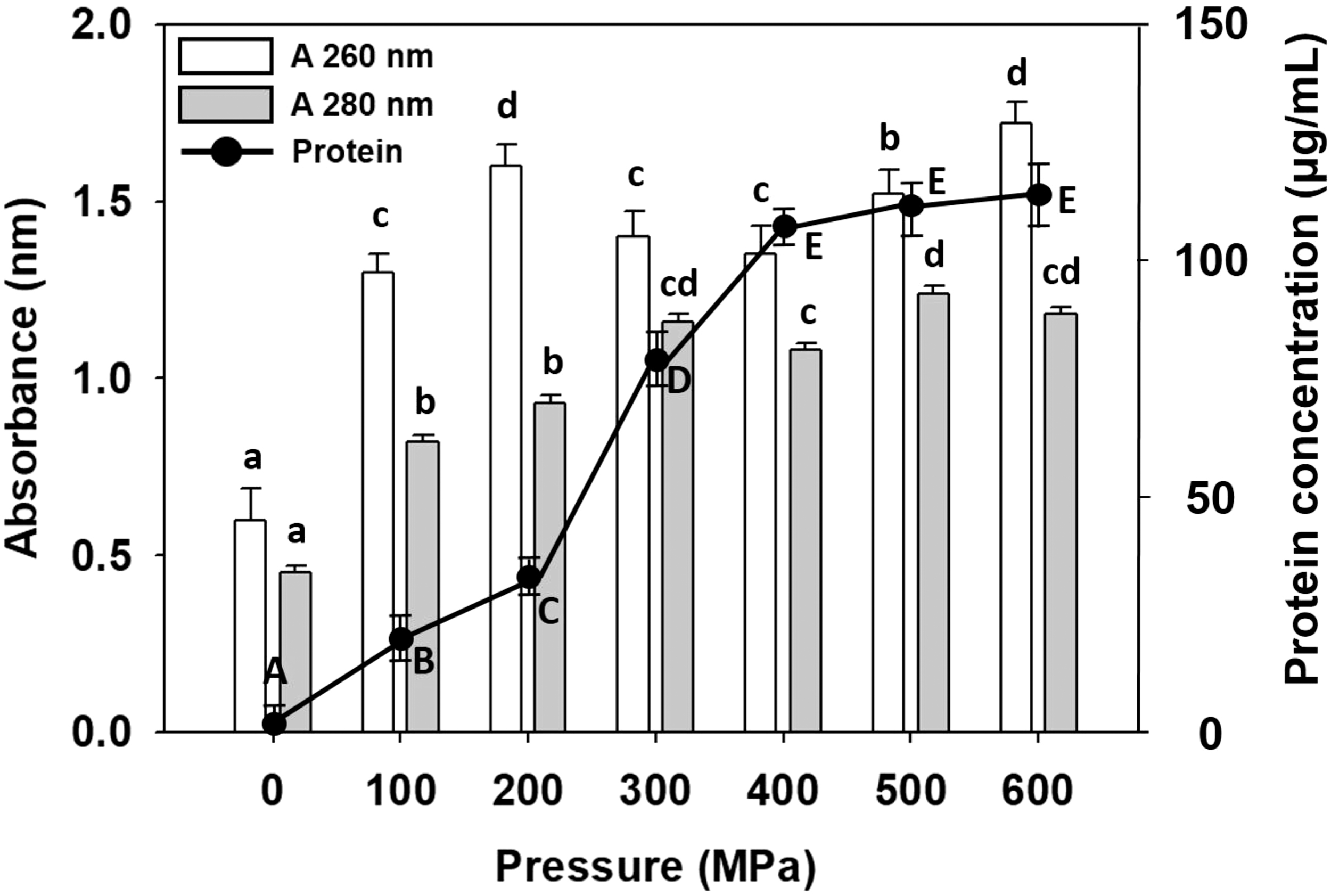
Nucleic acid (A 260 nm) and protein (A 280 nm and Bradford assay) concentrations in the culture supernatants of pressure-treated Aspergillus flavus. The values are represented as the mean of three independent experiments. Data denoted with the different letter combination in the same measurement are significantly different (p > 0.05).
Protein analysis
Figure 4 shows the effects of varying degrees of pressure on the total cell protein content in A. flavus spores. Proteins were extracted from the spores after the HHP treatment and analyzed using SDS-PAGE. On the SDS-PAGE electropherogram, we could observe four major protein bands in the control A. flavus spore total protein samples with molecular weights ranging 35–110 kDa. In the electrophoresis gel, the quantity and brightness of the protein bands changed depending on the varying degrees of pressure treatment. In the 100–400 MPa HHP group, the protein bands were still similar to those of the control, although they became fainter. However, the 35, 48, and 110 kDa bands became significantly fainter in the 500 MPa treatment group, and the brightness was significantly lower compared with the bands from the untreated spore samples, showing that HHP affected fungal morphology, causing some proteins to undergo denaturation or hydrolysis. When the treatment pressure was 600 MPa, the most significant difference was that the 35, 48, 75, 100, and 110 kDa bands, that were present in the control, disappeared in the 600 MPa treatment group. These results showed that the hydrogen and covalent bonds of certain proteins in the spore structure were broken after HHP, the intact protein bonds were destroyed, and some proteins were released. These results were consistent with the results in Figure 4. In Figure 4, the 500 to 600 MPa treatment groups released the most protein into the culture medium, showing that the released protein amounts increased with the increasing pressure. Taken together, these results collectively suggest that the proteins that disappeared in the protein gel presented in Figure 4 due to the pressure treatment were released into the culture medium. Similar results were also observed in the study published by Phuvasate and Su (2015). They found that high-pressure treatment disrupted the structure and protein composition of Vibrio parahaemolyticus cells, and the two protein band intensities at 150 and 250 kDa molecular weights decreased with increasing pressure. In addition, as the processing pressure increased, the intensity of some protein bands between 75 and 100 kDa decreased. These results showed that high-pressure processing damaged the cell membrane and changed the protein spectrum of V. parahaemolyticus cells, resulting in cell inactivation. The level of inactivation was directly proportional to the processing pressure. Huang et al. (2015) reported that after L. monocytogenes cells were treated with a pressure of 450 MPa for 5 min, the major protein bands (25, 40, and 45 kDa) disappeared compared with the control group. This might be one of the most important mechanisms by which high pressure causes microbial inactivation.

Sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis of the Aspergillus flavus spore membrane proteins after high-pressure treatment (100–600 MPa). Color images are available online.
Effects of HHP on cell membrane integrity
The evaluation of the PI-absorbing HHP-treated A. flavus spore proportions was carried out to understand the correlation between the level of inactivation and PI absorption rate. The HHP causes spore inactivation through cell membrane disruption, while PI is known to penetrate the damaged cell membranes and stain only the inactivated or damaged spores and emit a resulting bright red fluorescence. Figure 5 shows that the fluorescence intensity increased and the viability decreased with increasing treatment pressures. The residual number of 100 MPa and 200 MPa-treated spores was greater than 70% with only a minority of spores being eliminated, and the PI absorbance was lower compared with the 400 MPa and abovementioned pressure treatment groups, but was still significantly higher than the control group. When the pressure was 500 MPa, the residual fungal count was lower than the detection limit. Therefore, when PI reached its maximum absorption rate, the residual fungal count was similar to the 600 MPa treatment group and the PI absorbance did not significantly increase. The degree of PI absorption in the spores and the viability under different treatment pressures inversely correlated (i.e., the greater the absorption the lower the viability). This shows that high pressure could inactivate most of the spores. Kalagatur et al. (2018) treated maize with 100, 200, 300, and 380 MPa pressure and carried out PI staining. The effects of the different pressure treatments on the disruption of fungal spore morphology in maize were observed and the results showed that the proportion of PI-absorbing spores increased simultaneously with the increasing processing pressure. This showed that high pressure disrupted spore morphology and structure, increasing the opportunity for PI binding to the nucleic acids, leading to increasing stained spore numbers. We could conclude from the PI measurements that the HHP treatment disrupted the cell membrane to inactivate the fungal spores, which might be key to the HHP effector mechanism of fungicidal activity. This is the first study to show that HHP could effectively inactivate the spores of A. flavus and we could thus conclude that HHP could be used as an effective technology for fungal spore inactivation.
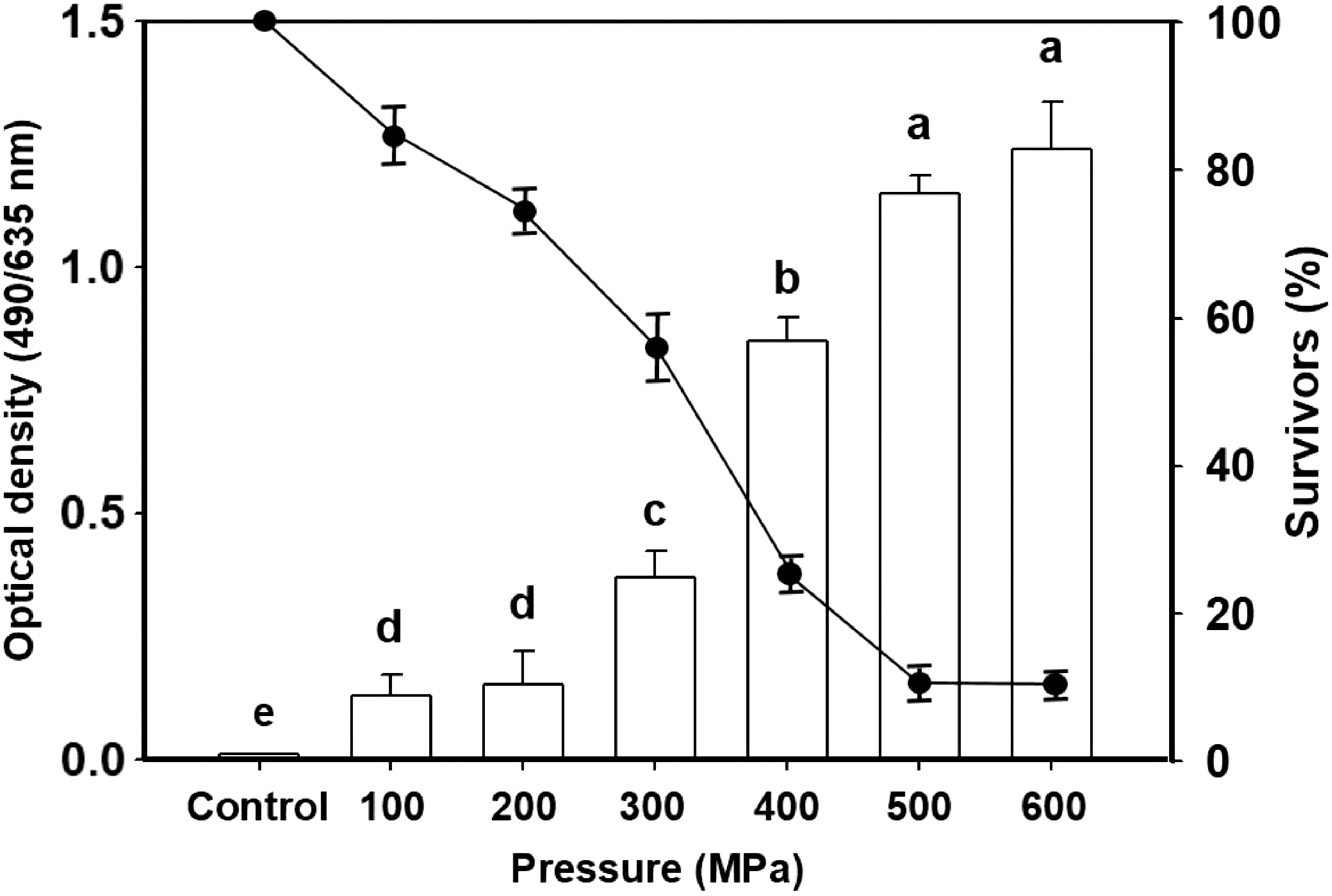
Relationship between the uptake of Propidium Iodide (□) and the survivors (%) (●) during the Aspergillus flavus spore pressure treatment.
Conclusion
High-pressure treatment is widely used to eliminate food pathogens. However, most studies examined the effects of high pressure on bacterial morphology while only a few studies examined the fungicidal mechanisms of high pressure. A. flavus is a common food pathogen that produces aflatoxin, resulting in significant agricultural crop loss. To the best of our knowledge, our study is the first to document the correlation between HHP-induced A. flavus spore inactivation and cell damage with changes in the cellular protein components. This study revealed the tolerable pressure treatment range for the A. flavus spores; lower and mild pressure treatment (100 to 400 MPa) caused little or no inactivation. Higher pressure at 500 to 600 MPa combined with a suitable duration could completely inhibit A. flavus spore activity. The pressure treatment resulted in mechanical damage to the spores, causing collapsed morphology and in turn affecting the intra- and extracellular spore morphology. When the cells were inactivated or damaged, PI absorption increased, while the intracellular components (such as nucleic acids and proteins) were simultaneously released. We hope that the results of this study could help to better understand the major mechanisms of A. flavus spore inactivation under HHP conditions, which could potentially contribute to the future application of high-pressure processing in pasteurization of foodstuffs that are at risk of A. flavus contamination.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This research work was supported by the Ministry of Science and Technology, MOST 107-2221-E-150 -053 -MY2, Taiwan, Republic of China.