
Abstract
The frequent occurrence of sequence-type 398 (ST398) livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) in pigs has become a major public health concern owing to the increased zoonotic potential of the pathogen. Recently, a novel oxazolidinone resistance gene, chloramphenicol–florfenicol resistant (cfr), conferring multiresistance phenotypes to phenicols, lincosamides, oxazolidinones, pleuromutilins, and streptogramin A (PhLOPSA), has been found among ST398 LA-MRSA strains isolated from pigs. In this study, we report the first in silico genome analysis of a linezolid-resistant ST398 LA-MRSA strain, designated PJFA-521M, recovered from a pig in Korea. Genomic analyses revealed that the presence of the cfr gene was responsible for the observed linezolid resistance in the PJFA-521M strain. Moreover, newer antimicrobial resistance genes, such as the dfrG, aadE, spw, lsa(E), lnu(B), and fexA genes, were found in the PJFA-521M strain. In addition to the genetic elements for antimicrobial resistance, the carriage of various virulence genes for adherence, invasion, and immunomodulation was identified in the genome, especially within several mobile genetic elements (MGEs). The presence of multiple antimicrobial resistance genes and virulence genes on MGEs in the genome of a linezolid-resistant ST398 LA-MRSA should raise awareness regarding the use of other antimicrobial agents in pig farms and may also provide selective pressure for the prevalence of the cfr gene and the associated multidrug-resistant phenotype.
Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) is one of the most important pathogens causing health care and community-associated (CA) infections with high mortality and morbidity rates (David et al., 2010). The recent emergence of a livestock-associated (LA) clone of MRSA with multilocus sequence-type (ST) 398 featuring a staphylococcal cassette chromosome mec type V (SCCmec V) has been a serious public health problem (He et al., 2018; Pirolo et al., 2019; Back et al., 2020). Although ST398 LA-MRSA has mainly been detected in pigs and pig farm environments, zoonotic transmission of ST398 LA-MRSA to humans through direct or indirect contact is increasing (Cuny et al., 2009; He et al., 2018; Pirolo et al., 2019). Furthermore, fatal cases of infections with human-adapted clones of ST398 CA-MRSA have been reported in China (He et al., 2018). Previous studies about virulence determinants of ST398 and closely related STs within clonal complex (CC) 398 revealed that CC398 LA-MRSA lacked several major staphylococcal virulence genes encoding immune evasion cluster (IEC), toxic shock syndrome toxin 1, exfoliative toxins, and Panton–Valentine leukocidin (PVL) (Price et al., 2012; Sahibzada et al., 2017). However, several uncommon antimicrobial resistance genes for S. aureus, such as phenicol exporter gene fexA, linezolid resistance gene cfr (chloramphenicol–florfenicol resistant), tetracycline resistance gene tet(L), and macrolide–lincosamide–streptogramin B (MLSB) resistance genes erm(T), have been identified in CC398 LA-MRSA strains (Kehrenberg et al., 2006; Kadlec et al., 2009; Gomez-Sanz et al., 2013).
The cfr gene encodes methyltransferase, which facilitates methylation of the adenosine residue at position 2503 in 23S rRNA. This modification results in a multiresistance phenotype, with resistance to
Materials and Methods
A linezolid-resistant ST398 LA-MRSA isolate
An ST398 SCCmec V LA-MRSA strain designated PJFA-521M was isolated from a nasal swab sample of a healthy finishing pig in Jeolla province, Korea in 2018 (Back et al., 2020). The PJFA-521M strain was identified using 16S rRNA sequencing (BIONICS, Seoul, Korea) and Vitek 2 system (bioMerieux, Durham, NC). Molecular characterization of the PJFA-521M revealed that this strain carried SCCmec V for methicillin resistance, belonging to multilocus sequence type 398 with an allelic profile of 3-35-19-2-20-26-39, spa type t18103, and agr type I. This strain showed resistance to ampicillin, cefoxitin, chloramphenicol, ciprofloxacin, clindamycin, erythromycin, gentamicin, quinupristin-dalfopristin, and tetracycline. The minimum inhibitory concentrations (MICs) of oxacillin and tetracycline were 8 and 64 μg/mL, respectively. The MIC to linezolid was determined using a standard E-test® (bioMerieux) on a Mueller–Hinton agar plate according to the manufacturer's instructions.
A list of antimicrobial agents, breakpoints for susceptibility tests, and profiles of MIC are given in Supplementary Tables S1 and S2.
Extraction of genomic DNA and whole-genome sequencing
Genomic DNA of the PJFA-521M strain was extracted using the TruSeq DNA PCR free Kit (Illumina SD). Assessment of the quality and quantity of the bacterial genomic DNA sample was performed using a NanoDrop 2000c spectrophotometer (Thermo Fisher Scientific, Wilmington). DNA sample with the absorbance (260/280 nm) ratio ∼2.0 was considered a good quality DNA and 1.0 μg of total DNA was used for the library construction (Huptas et al., 2016).
The whole-genome library of the PJFA-521M strain was sequenced using the Illumina HiSeq platform (150 bp paired-end reads) with 4,724,402 reads of 151 bases. The raw reads were trimmed out to remove low-quality and adaptor sequences using Trimmomatic (v0.36), resulting in 4,722,757 high-quality reads (Bolger et al., 2014). SPAdes (v3.13) was used for the generation of a de novo-assembled genome consisting of 54 contigs with 2,895,108 reads (Bankevich et al., 2012). The assembled contigs were then annotated with Prokka (v1.12) (Seemann, 2014).
Genomic annotation and analysis
Clusters of Orthologous Groups (COG) and SEED (
The multilocus sequence typing, SCCmec, and spa types were confirmed using the Center for Genomic Epidemiology (CGE) tools (
For the analysis of mobile genetic elements (MGEs), such as prophages and staphylococcal pathogenic island (SaPI) regions in the genome, the PHAge Search Tool Enhanced Release (PHASTER) (
Mechanisms of linezolid resistance
Linezolid resistance in staphylococci has been described as: (i) mutations in domain V of the 23S rRNA-binding site; (ii) mutations in the rplC, rplD, and/or rplV genes encoding 50S ribosomal proteins L3, L4, and L22; and (iii) acquisition of ribosomal methyltransferase gene, cfr (Stefani et al., 2010). Two novel genes (optrA and poxtA), which confer resistance to oxazolidinones, have also been reported in MRSA and enterococci (Antonelli et al., 2018; Ruiz-Ripa et al., 2019). All the determinants associated with linezolid resistance, including the presence of optrA and poxtA, were analyzed in the PJFA-521M strain.
Phylogenetic analysis
For phylogenetic analysis of the ST398 MRSA strains and closely related ST541 MRSA strains (CC398 MRSA strains), we downloaded raw sequence files of 89 strains by Price et al. (2012) from the Sequence Read Archive (
In addition, the profiles of antimicrobial resistance genes among CC398 strains were investigated using the National Center of Biotechnology Information's AMRFinderPlus, UniprotKB, and BLAST search (>90% coverage and identity).
Nucleotide sequence accession number
The draft genome of ST398 LA-MRSA strain PJFA-521M has been deposited in the GenBank database under the accession number SRKD00000000.
Results
Genomic features of the ST398 LA-MRSA strain PJFA-521M
The draft genome of ST398 MRSA strain PJFA-521M contains a circular chromosome with a length of 2,895,108 bp, coverage of 150.9 × , and GC content of 33%. A total of 2760 annotated genes possessed 2695 open-reading frames (ORFs), 57 tRNAs, and 7 rRNAs. Among 2695 ORFs, 2011 genes (74.6%) were predicted to encode functional proteins, whereas 684 genes (25.4%) were uncharacterized or hypothetical.
The functional categorizations of COG and SEED are given in Figure 1. In the COG categorization, the five major COG categories were as follows: 211 ORFs of amino acid transport and metabolism (7.8%, category E); 161 of inorganic ion transport and metabolism (6%, category P); 150 of translation, ribosomal structure, and biogenesis (5.6%, category J); 143 of replication, recombination, and repair (5.3%, category L); and 140 of carbohydrate transport and metabolism (5.2%, category G). Likewise, ORFs belonging to amino acids and derivatives (273), carbohydrates (178), protein metabolism (160), cofactors, vitamins, prosthetic groups, pigments (117), and DNA metabolism (91) were mainly distributed within the SEED subsystem categorization. In addition, 68 ORFs and 25 ORFs belonged to the virulence, disease-defense category and MGEs, respectively. The functional characterization of genes in PJFA-521M revealed that metabolic functions comprise 32% of the S. aureus genome.
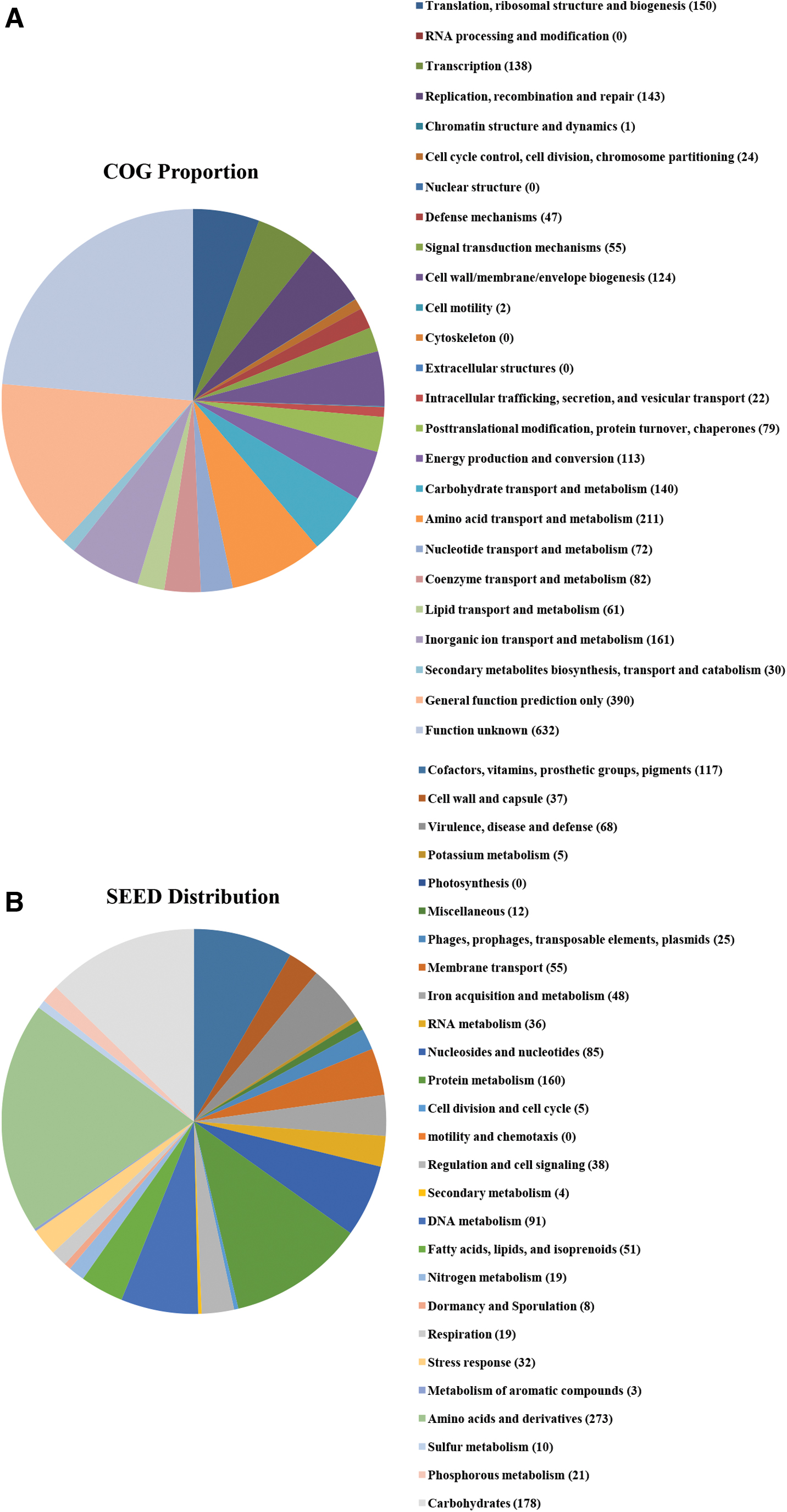
Analysis of functional categories in the genome of strain PJFA-521M based on the
Virulence-associated genes
The LA-MRSA stain PJFA-521M carried a series of virulence-associated factors in the genome and the catalog of virulence genes are given in Table 1. Well-recognized virulent determinants of S. aureus including protein A (spa), hemolysins, staphylococcal enterotoxin, and exotoxins, were also present in PJFA-521M. A variety of genes encoding adhesion-associated proteins were dispersed at several loci. In particular, three innate immune modulator genes, scn-1, scn-2, and scn-3, and a bi-component pore-forming leukocidin gene, lukGH, were identified.
Virulence Genes of PJFA-521M
As with the antimicrobial resistance genes, MGEs have an important role in the acquisition of new traits of pathogenicity in MRSA. Many virulence genes promoting immune modulations for host adaption were inserted in the various MGEs in the PJFA-521M strain. Five SaPIs (SaPI-S0385, SaPI1, SaPI2, SaPI5, and SaPIbov4), two genomic island regions (νSa3 and νSaα), and four prophage regions were identified in the PJFA-521M genome. Two hemolysin-encoding genes, scn and vwb, were integrated into the SaPI-S0385 region. A set of staphylococcal superantigen-like toxins and lipopeptide-like genes (lpl) were also located within the νSaα island in contig 14.
Antimicrobial resistance genes
The ST398 LA-MRSA strain PJFA-521M had linezolid MIC value of 12 μg/mL, which indicated linezolid resistance (MIC ≥8 μg/mL as a linezolid-resistant S. aureus). In our previous study, the pig-associated ST398 LA-MRSA strain, PJFA-521M, exhibited resistance to multiple classes of antimicrobial agents and zinc chloride (Back et al., 2020). Although the polymerase chain reaction-based detection method used in a previous study identified the mecA gene for β-lactams resistance, as well as the tet(M) and tet(L) genes for tetracycline resistance, in silico analyses revealed the presence of additional multiple resistance genes; four aminoglycoside resistance genes in contigs 31, 46, and 47; a trimethoprim resistance dfrG gene located in contig 33; and the MLS resistance erm(T), lsa(E), and lnu(B) genes in contigs 31 and 34 (Table 2). In accordance with a previous study (Kaya et al., 2018), SCCmecFinder uncovered the presence of SCCmec V (5C2) carrying mecA and ccr genes in contig 5. Of note, the cfr gene involved in PhLOPSA resistance was identified in contig 44 and the fexA gene conferring phenicol resistance was detected in contig 42. As given in Table 2, two operons involved in copper resistance, copAZ and copB-mco, were integrated in contig 4. Cadmium and zinc resistance genes were located in contigs 2, 9, and 27, respectively.
Antimicrobial Resistance Genes of PJFA-521M
Most of the antimicrobial resistance genes identified in PJFA-521M were flanked by insertion sequences (ISs) and transposons; IS257 carried a gene cluster of aadE, spw, lsa(E), and lnu(B) in contig 31; putative IS and bacteriophage sequences contained a novel trimethoprim resistance gene (dfrG) and blaZ gene in contig 33; a putative ISSau10 contained tet(L) and erm(T) in contig 34; fexA and cfr genes in contig 42 and 44 were flanked by putative transposons Tn558 and Tn552, respectively.
cfr-mediated linezolid resistance
The PJFA-521M strain carried the cfr gene for the observed linezolid resistance. However, the optrA and poxtA genes were not detected in the whole-genome sequence analysis. Moreover, no mutations in domain V of the 23S rRNA and the ribosomal proteins L3, L4, and L22 were identified in PJFA-521M, indicating that the linezolid resistance in PJFA-521M was caused by acquisition of the cfr gene.
BLAST analyses revealed that the contig 44 containing cfr gene has 100% sequence similarity to the 39,243 bp-sized plasmid p14-01514 of linezolid-resistant S. epidermidis (NZ-KX520649) (Wessels et al., 2018). Moreover, the cfr and fexA genes were assembled into one segment consisting of Tn552 and Tn558, which showed >99.9% sequence similarity to the plasmid p14-01514 (Supplementary Fig. S2A, B).
Phylogenetic lineage and antibiogram of CC398 isolates
A phylogenetic tree of 90 different CC398 S. aureus strains including PJFA-521M is given in Figure 2. PJFA-521M was most closely related to four of the ST398 LA-MRSA strains (UB08116, 12152-5, 44523-1, and 30116 strains) isolated in European countries. All the CC398 MRSA and methicillin-susceptible Staphylococcus aureus (MSSA) strains were resistant to tetracycline and carried one or more tetracycline resistance genes. Of note, all the CC398 S. aureus strains had the tet(38) gene except one pig-originated MRSA strain (AV6) (Fig. 2). However, only 18% (16/90) of the CC398 MRSA strains carried the dfrG gene, which was identified in PJFA521M for trimethoprim resistance. The frequencies of two phenicol resistance genes, cat and fexA, among the 90 CC398 MRSA strains were 4% and 6%, respectively (data not shown). None of the strains carried genes associated with mupirocin and fusidic acid resistance. Of importance, only 2/90 CC398 S. aureus strains (PJFA-521M and 09/01691/3 strains) were positive for the cfr gene. The genetic environments of the cfr gene in PJFA-521M and 09/01691/3 were identical (Supplementary Fig. S2B).

Phylogenetic relationship and antibiogram of CC398 strains. Color images are available online.
Discussion
Although CC398 LA-MRSA has mainly colonized pigs, human infections with CC398 LA-MRSA through direct or indirect contact with various livestock animals have been increasingly reported in recent years (Cuny et al., 2009; He et al., 2018; Pirolo et al., 2019). Genomic analyses of the CC398 lineage of LA-MRSA revealed that gain or loss of certain genetic components was frequent during its transmission between animal and human hosts (Price et al., 2012).
The absence of pvl genes and ϕSa3 prophage containing IEC in porcine-derived CC398 strains supports the notion that these genes are associated with immunomodulation in humans but not in nonhuman hosts (Price et al., 2012). Similarly, the PJFA-521M strain was also negative for lukS/F genes encoding PVL. However, novel leukocidins designated lukG and lukH (also known as lukAB) were detected in the genome of PJFA-521M. LukGH displaying cytotoxic potency in human phagocytes was initially described in the USA300 CA-MRSA TCH1516 strain and subsequently identified in a ST398 LA-MRSA isolate (Schijffelen et al., 2010; Trstenjak et al., 2019). The sequence analyses of LukGH identified in the PJFA-521M strain resulted in 99% and 89% amino acid identity to those of the ST398 LA-MRSA S0385 and USA300 CA-MRSA TCH1516 (CP000730) strains, respectively. Based on previous studies, LukGH displays a marked host-species specificity, and variations in LukH may greatly impact the pathogenesis of MRSA during its transmission between pig and human hosts (Ventura et al., 2010; Badarau et al., 2015; Trstenjak et al., 2019).
A homolog of staphylococcal pathogenicity island (SaPI-S0385) harboring scn and vwb was also identified during the PJFA-521M genome analysis (Schijffelen et al., 2010). Staphylococcal complement inhibitor (SCIN), a product of the scn gene, interferes with complement activation by blocking C3 convertases. The vwb homolog in the PJFA-521M genome encodes von Willebrand factor-binding protein (vWbp), which binds to the vWf protein. It has been shown that both human and porcine serums displayed similar levels of coagulation rates after exposure to vWbp (Bjerketorp et al., 2004). Although a complete understanding of the host range for scn and vwb homologs on SaPI-S0385 is not available, the presence of these genes may broaden host specificity during transmission between human and animal hosts.
In addition to immunomodulators, various genes encoding adhesion factors were identified in the genome of the PJFA-521M strain. A number of cell wall-anchored proteins and secreted proteins appear to be involved in adherence to host cells during staphylococcal colonization (Tulinski et al., 2014). In addition to their binding to host cell molecules, such as fibrin and fibrinogen, ClfA, ClfB, FnBPA, FnBPB, SdrC, SdrD, and SdrE, the role of enhanced staphylococcal immune escape or host cell invasion was reported (Tulinski et al., 2014). In a previous report (Schijffelen et al., 2010), it was demonstrated that the ST398 LA-MRSA strain S0385 carried several unique MGEs containing genes involved in host-cell adhesion. The presence of MGEs containing scn, vwb, and tet(M) was identified only in animal-derived ST398 MRSA strains (Schijffelen et al., 2010). Similarly, the PJFA-521M strain harbored various unique MGEs, especially a homolog of vwb-containing MGEs, in ST398 LA-MRSA S0385 strain. SCIN encoded by scn identified in SaPI-S0385 and ϕ3, which inhibits the complement activation by interfering with C3b deposition, resulted in a reduction in pathogen killing of human neutrophils (Rooijakkers et al., 2005). Proteins encoded by vwb (vWbp), which binds to vWf, play multifunctional roles in the coagulation activity with host prothrombin in plasma (Viana et al., 2010; Thomer et al., 2013). Moreover, surface-bound protein A of S. aureus also increases the adherence ability and damage of host cells by interacting with vWf (Viela et al., 2019). These results suggest that the carriage of scn and vwb genes may help evade host innate immunity and facilitate adaptation of ST398 LA-MRSA in human host.
The carriage of multiple antimicrobial resistance genes responsible for multidrug-resistant phenotypes has recently been reported in ST398 LA-MRSA strains (Sarrou et al., 2016; Fessler et al., 2018). The tetracycline resistance phenotype along with the resistance genes have frequently been found in CC398 LA-MRSA strains (Price et al., 2012; Eom et al., 2019). The presence of the chromosomal tet(M) gene has been considered a robust genetic marker for CC398 LA-MRSA strains (Price et al., 2012). In addition, dual carriage of tet(M) and plasmid-borne tet(K) or tet(L) genes was observed in many LA-MRSA strains, which displayed higher MIC values for tetracycline than the strains carrying a single gene (Kadlec et al., 2010; Back et al., 2020). In this study, the presence of both tet(M) and tet(L) in the PJFA-521M strain has also been confirmed, which may explain the high MIC (64 μg/mL) and minimum bactericidal concentration values (128 μg/mL) to tetracycline.
Recently, the detection of rather uncommon resistance genes has been reported in CC398 LA-MRSA isolates (Kadlec et al., 2010; Gomez-Sanz et al., 2013). The MLSB resistance gene, erm(T), the trimethoprim resistance gene dfrG, and the tetracycline resistance gene tet(L) were identified in the genome of the PJFA-521M strain (Supplementary Fig. S1). The novel gene cluster of erm(T), dfrK and tet(L) was initially found on the plasmid pKKS25 of a ST398 LA-MRSA isolate from a sow (Kadlec et al., 2010). Instead of drfK, the PJFA-521M strain carried dfrG in contig 33, which showed 100% nucleotide sequence similarity to that of the CM.S2 MRSA strain (GenBank Accession No. IMCJ1454) (Sekiguchi et al., 2005). Although occurrence of dfrG and dfrK genes has been rare in animals, the presence of dfrK and tet(L) gene cluster may affect emergence of trimethoprim resistance genes under the selective pressure of tetracycline in pig farms (Back et al., 2020). As presented in Supplementary Figure S1, the genetic composition of contig 31 containing aadE, spw, lsa(E), and lnu(B) genes in the PJFA-521M strain showed 99% nucleotide sequence identity to the chromosomal integration region of MSSA C2828 strain (GenBank Accession No. JQ861959) (Lozano et al., 2012). The novel aminoglycoside and lincosamide resistance gene clusters in a multiresistance plasmid have been previously described in animal isolates (Wendlandt et al., 2013; Sarrou et al., 2016). Collectively, these data indicate that colocalization of multiple resistance genes conferring resistance to various classes of antimicrobial agents may enable their co-selection and persistence even in the absence of a direct selective pressure in pig farms.
Linezolid has been exclusively used for clinical practice as a last-resort antimicrobial for the treatment of MRSA and Gram-positive bacterial infections (Choo et al., 2016; Wessels et al., 2018). Although it is not approved for use in veterinary medicine, the occurrence of linezolid-resistant S. aureus and staphylococci in food-producing animals has caused significant public health concerns, especially owing to the cfr-mediated multiresistance PhLOPSA phenotype (Cuny et al., 2017; Li et al., 2018). In the PJFA-521M strain, the cfr gene was identified within 3216-bp contig 44, which exhibited 100% nucleotide sequence identity with the plasmid p14-01514 of S. epidermidis (NZ_KX520649) (Wessels et al., 2018). Moreover, identification of the florfenicol–chloramphenicol exporter gene fexA in contig 42 suggests that fexA and cfr are colocalized on a plasmid of the PJFA-521M strain (Supplementary Fig. S1). Co-selection of cfr and fexA in MRSA and other staphylococci has been linked to the widespread use of chloramphenicol and florfenicol in swine farms (Kehrenberg et al., 2006; Shen et al., 2013; Zhao et al., 2016; Sun et al., 2018) and coexistence of cfr and fexA with additional resistance gene such as aacA-aphD, tet(L), dfrK, erm(A), erm(B), or lsa(B) may allow the cfr and fexA genes to be co-selected under nonphenicol use in farms (Kehrenberg et al., 2007; Shen et al., 2013). Besides the cfr gene, a novel oxazolidinone resistance gene, optrA, has been reported in staphylococci from pigs (Fan et al., 2016, 2017). In this study, whole-genome sequence analysis confirmed the absence of the optrA gene in the PJFA-521M strain. However, large-scale investigation of the surveillance of the cfr and optrA genes in staphylococci and enterococci of livestock origin is needed to assess the prevalence of these genes in the farm environment. To the best of our knowledge, this is the first report of whole-genome sequence analysis of an ST398 LA-MRSA strain isolated from a pig in Korea.
Conclusions
Our results demonstrate that: (i) the linezolid-resistant ST398 LA-MRSA strain isolated from a pig carries the cfr gene for the PhLOPSA resistance phenotype; (ii) in addition to cfr for linezolid resistance, the tet(L), erm(T), dfrG, aadE, spw, lsa(E), lnu(B), and fexA genes were identified in PJFA-521M, which may help co-select the strain under exposure to various antimicrobial agents in the pig farm environment; and 3) the PJFA-521M strain harbored various unique MGEs, some of which contain host-specific virulence factors for human and animal host infections.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was carried out by the support of Research of Korea Centers for Disease Control and Prevention (Project No. 2020ER540500).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.