
Abstract
The prevalence of Listeria monocytogenes in raw beef and in slaughterhouse environments was investigated from April 2019 to February 2020. Three hundred raw beef samples were purchased from 50 retailers and 10 restaurants (5 samples per source). One hundred and thirty-four samples from slaughterhouse environments were collected by swabbing (10 × 10 cm) the surfaces, gloves, splitting saw, and drains. L. monocytogenes was detected and identified according to the method described in ISO 11290-1, and confirmed by 16S rRNA sequencing. L. monocytogenes was detected in raw beef (2/300, 0.7%), gloves used in carcass splitting (6/21, 28.6%), the splitting saw (1/18, 5.6%), and the drain zone (1/15, 6.7%). All isolates were serotype 1/2a or 1/2c, based on screening using multiplex PCR-based serogrouping assay and serotyping kit for O-H antigens. Pulsed-field gel electrophoresis (PFGE) following ApaI digestion of eight PFGE pulsotypes and four PFGE groups were identified. Biofilm formation analysis using Crystal Violet staining revealed the highest biofilm formation in strain LM-16, followed by D190613. Although L. monocytogenes isolates were susceptible to most antimicrobials, some resistance to penicillin (8/15, 53.3%) and tetracycline (2/15, 13.3%) was observed. Through PFGE, G190426, G190829, and G200210 isolated from the same location in this study were genetically homologous similar to the LM-16 strain, previously isolated from beef carcass in 2006. These results suggest that LM-16 has been continuously present in biofilms in the slaughterhouse environments since 2006. Our study indicates that L. monocytogenes contamination in raw beef could consistently occur during beef processing in slaughterhouse environments through contact with gloves, splitting saws, and drains.
Introduction
L
Among the 13 serotypes of L. monocytogenes, serotypes 1/2a and 1/2b account for over 50% of the strains isolated from foods and food processing environments (Lukinmaa et al., 2003). A primary step in investigating cases of human listeriosis linked to contaminated foods is epidemiological subtyping, at the subspecies level, to identify and discriminate L. monocytogenes (Bae et al., 2014). Pulsed-field gel electrophoresis (PFGE) is one of these genotyping methods, and the exchange of bacterial PFGE patterns through PulseNet, a national network of public health and food regulatory agency laboratories coordinated by the U.S. Centers for Disease Control (51;
L. monocytogenes can persist in food processing environments by forming biofilms on food processing equipment, on workers' gloves, and in drains, which may disperse and contaminate the product (Carpentier and Cerf, 2011). Biofilm formation by L. monocytogenes varies among serotypes, lineage, origin of isolation, and intrinsic and extrinsic factors, such as the nutrient level and temperature (Kadam et al., 2013). L. monocytogenes in biofilms can cause sepsis, meningitis, and flu-like symptoms in pregnant women, fetuses, and neonates, through cross-contamination (Aureli et al., 1992).
Ampicillin or penicillin, combined with gentamicin, are the primary treatments for human listeriosis (Hof, 2003). However, an increasing number of strains are becoming resistant to most of the known antimicrobials, including those used for treating listeriosis over the past decade (Charpentier and Courvalin, 1999; Lyon et al., 2008). Resistance of L. monocytogenes isolated from raw beef and slaughterhouse environment has not been actively investigated in the most common sources of infection. The objective of this study was to determine the prevalence, characteristics, and antimicrobial sensitivity of L. monocytogenes, isolated from retail and slaughterhouse environments, and to trace the origins of L. monocytogenes in the production process.
Materials and Methods
Sample collection
Between April 2019 to February 2020, 300 raw beef samples were purchased from 50 retailers and 10 restaurants (5 samples per source), and 144 slaughterhouse environment samples were collected from surfaces (10 × 10 cm) by swabbing a beef carcasses before (n = 25) or after washing (n = 25), workers' gloves (n = 51), splitting saws (n = 18), and drains (n = 15) with the Pipette Swab Kit (3M, St. Paul, MN) in a slaughterhouse where over 1000 cattle and pigs are slaughtered per day in Kyeonggi-do, South Korea. All samples were individually wrapped, stored in an ice chest, and transported to the laboratory within 2 h for immediate processing.
Strains
To compare the genetic characteristics of previously isolated strains of L. monocytogenes with those found in this study, we also tested strains isolated from beef carcass (n = 2), clinical samples (n = 2), and a type strain (n = 1). L. monocytogenes strains were kindly provided by the Animal and Plant Quarantine Agency (APQA: LM-16-5, LM-16), Korea Disease Control and Prevention Agency (NCCP 15743, NCCP 14714), and the American Type Culture Collection (ATCC 51776).
Isolation of L. monocytogenes
The presence of L. monocytogenes in food samples was detected by culturing, according to the methods described in ISO 11290-1 (ISO, 1996). The samples were placed in 225 mL Listeria enrichment broth (Oxoid, Ltd., Basingstoke, United Kingdom), homogenized in a BagMixer stomacher (Interscience, St Nom, France) for 2 min, and incubated at 30°C for 24 h. Aliquots (100 mL) of these primary enrichments were transferred to 10 mL of Fraser broth (secondary enrichment), inoculated on Oxford agar (Oxoid, Ltd.), and incubated at 37°C for 24 h. Listeria spp. colonies were selected and inoculated on Tryptone-Soy agar (TSA; Oxoid Ltd.) supplemented with 0.6% yeast extract (YE; Himedia Laboratories Pvt. Ltd., Mumbai, India) (TSA-YE), and incubated at 37°C ± 2°C for 24 h. The strains were identified through 16S rRNA sequencing, using the primers: 27F: 5′-AGA GTT TGA TCM TGG CTC AG-3′ and 1492R: 5′-GGT TAC CTT GTT AGG ACT T-3′. Each 16S rRNA sequence was subjected to BLAST (
DNA extraction
DNA was extracted using NucliSENS easyMAG (BioMérieux, Marcy l'Etoile, France) according to the manufacturer's instructions. Briefly, each colony was lysed in 1 mL lysis buffer by incubating at room temperature (∼26°C) for 30 min. The lysed sample was then transferred to easyMAG disposable sample strip wells containing 50 μL of magnetic silica and subjected to automatic magnetic bead isolation. DNA was then eluted in 100 μL of elution buffer.
L. monocytogenes serotyping
Multiplex PCR-based serogrouping assay using the serotyping kit for O-H antigens was employed for serotyping (Doumith et al., 2004). Each PCR was performed in a reaction volume of 20 μL, comprising FRENCHE PCR PreMix (iNtRON Biotechnology, Seongnam, Korea), 2 μL of template DNA, 1 μL of each primer, and 16 μL distilled water. In total, 5 μL of the amplified PCR product was analyzed by electrophoresis on a 1.5% agarose gel containing 50 μL of SafeView™ (Applied Biological Material, Inc., Richmond, Canada) per liter. PCR amplicons were examined under ultraviolet light using a Bio-Rad Molecular Imager® GelDoc™ XR (Bio-Rad Laboratories, Hercules, CA). Primer sequences are presented in the Supplementary Table S1. Serotyping was performed using the Listeria Antisera Set (Denka Seiken Co., Tokyo, Japan), according to the manufacturer's instructions. O (O-I/II, O-V/VI, O-I, OII, O-VI, OVII, O-VIII, and Y O-IX) and H (H-A, H-AB, H-C, and H-D) antigens were identified according to the serotyping scheme. Strains NCCP 15743, NCCP 14714, LM-16-5, and ATCC 19115 were the control strains for 1/2a, 1/2b, 1/2c, and 4b serotype, respectively.
Pulsed-field gel electrophoresis
PFGE was performed for the genetic characterization of 15 L. monocytogenes isolates, using the standard U.S. Centers for Disease Control and Prevention PulseNet protocol (
Biofilm formation assay
Biofilm formation assays were performed on polystyrene microplates by using a previously described method, with some modification (Stepanović et al., 2000). One colony from each L. monocytogenes isolate was inoculated in 9 mL of Brain Heart Infusion (BHI) broth (Oxoid, Ltd.) and incubated at 37°C for 24 h. Then, 100 μL of each culture (∼8.84 × 107 cells) was inoculated onto 96-well microtiter plates at 37°C for 24 h. After incubation, the microplates were carefully washed by pipetting with 200 μL of sterile phosphate-buffered saline (PBS; Oxoid, Ltd.). One hundred microliters of 0.1% Crystal Violet was added to each well of the microtiter plate and incubated for 15 min at room temperature, and the plates were then rinsed three times with PBS. The optical density (OD) of each well was measured at 595 nm using a Multiskan FC Microplate Reader (Thermo Fisher Scientific, Waltham, MA).
Antimicrobial susceptibility testing
The sensitivity of L. monocytogenes strains to 11 antimicrobials (10 μg ampicillin, 30 μg chloramphenicol, 5 μg ciprofloxacin, 15 μg erythromycin, 10 μg gentamicin, 30 μg kanamycin, 10 μg penicillin, 5 μg rifampicin, 30 μg tetracycline, 23.75/1.25 μg sulfamethoxazole–trimethoprim, and 30 μg vancomycin) (Millipore Sigma Co., St. Louis, MO) was assessed using the disk (Sigma Chemical Co., St. Louis, MO) diffusion method in accordance with the guidelines proposed by the Clinical and Laboratory Standard Institute (CLSI, 2015). Briefly, single colonies of L. monocytogenes were transferred to BHI broth (Oxoid, Ltd.) and incubated at 37°C for 24 h. Then, each suspension (0.5 McFarland) was spread onto the surface of Mueller Hinton Agar (Oxoid, Ltd.). The diameter of the inhibition zone around disks was measured after incubation at 37°C for 24 h. Staphylococcus aureus ATCC 25923 was a control strain.
Statistical analysis
GraphPad Prism 6.01 (GraphPad software, San Diego, CA) was used for data analysis. Biofilm formation was compared between slaughterhouse environment samples, using student's t-test and one-way analysis of variance (Tukey's method). Differences were significant at p < 0.05.
Results
Prevalence of L. monocytogenes in raw beef, and in slaughterhouse samples
In total, 10 strains of L. monocytogenes were isolated from the 300 samples of raw beef and 134 samples from the slaughterhouse environment (Table 1). The highest prevalence was observed on the gloves of carcass-splitting workers: G190426, G190522, G190613, G190829, G200210, and G200214 (6/21, 28.6%), followed by raw beef in retail markets: R190420-1 and R190420-2 (2/300, 0.7%), drains: D190613 (1/15, 6.7%), and splitting saws: S200214 (1/18, 5.6%). L. monocytogenes was not detected in raw beef samples from restaurants, beef carcasses before or after washing, or on gloves of workers involved in white and red offal processing.
Prevalence of Listeria monocytogenes in Raw Beef and Slaughterhouse Environment
Glove 1, 2, 3; white offal removal, red offal removal, and carcass splitting, respectively.
Glove 3 was detected in April, May, June, and August 2019, respectively, and two strains were detected in February 2020.
The strain was detected in February 2020.
The strain was detected in May 2019.
Two strains were detected in April 2019.
Phenotypic and genotypic diversity of L. monocytogenes isolates
The five L. monocytogenes isolates obtained from the KCDC, APQA, and ATCC, and the 10 isolated from raw beef and the slaughterhouse environments, belonged to four serotypes (1/2a, 1/2b, 1/2c, and 4b) (Fig. 1). The main serotype detected was 1/2a (10/15, 66.7%). Serotype 1/2c was detected in 4/15 (26.7%) of the strains, and serotype 1/2b and 4b were detected in 1/15 (6.7%) of strains, respectively. The isolates showed a PFGE banding pattern of A to H. They were further classified into groups 4 (group 1:A; group 2:C; group 3:D; group 4:F) based on their PFGE banding pattern, with over 90% similarity by ApaI-digestion. The dendrogram of PFGE profiles by ApaI-digestion divided them into groups according to banding pattern; PFGE pattern A: LM-16-5 (May, 2016) and D190613 (June, 2019); C: G190522 (May, 2019), G190613 (June, 2019), and S200214 (February, 2020); D: LM-16 (Unknown, 2006), G190426 (April, 2019), G190829 (August, 2019), and G200216 (February, 2020); and F: R190420-1 (April, 2019) and R190420-2 (April, 2019). All isolates that showed PFGE banding patterns 2 and 3 were classified as serotype 1/2a, and those with 1 and 4 were serotype 1/2c. Interestingly, isolates from the gloves of carcass-splitting workers exhibited similar PFGE pattern type, and were identical to the isolates from carcass splitting saw and slaughterhouse environment.
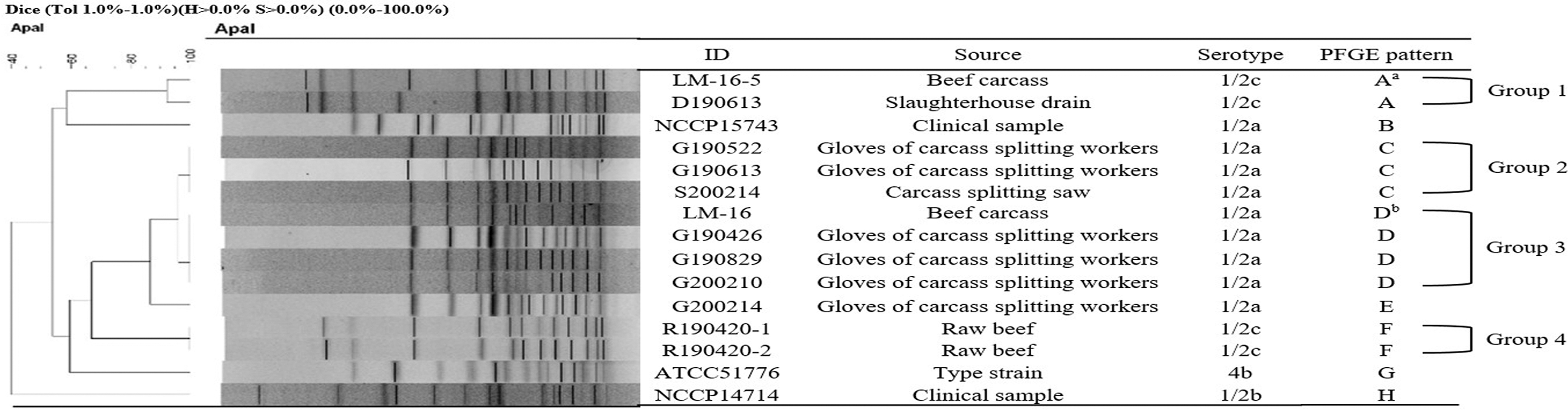
Analysis of PFGE patterns for Listeria monocytogenes isolated from raw meats, slaughterhouse environment, and reference strains. aIsolated in 2016 from the same slaughterhouse as from where L. monocytogenes corresponding to PFGE patterns A, C, D, and E were isolated. bIsolated in 2006. PFGE, pulsed-field gel electrophoresis.
Biofilm formation
The isolates formed biofilms, with an overall average absorbance of 0.29 (Fig. 2). Isolate LM-16 (beef carcass, isolated in 2006) had the highest ability to form biofilms, with an absorbance of 0.648, followed by D190613 (Drain, isolated in 2019), with an absorbance of 0.474 (p < 0.05). G190613 showed significantly lower biofilm formation ability than clinical strains (NCCP15743, NCCP14714); a type strain (ATCC 51776) showed no significant difference in all strains except D190613 and LM-16. Interestingly, biofilm formation ability was not significantly different in the raw beef and slaughterhouse environment isolates, except for D190613.
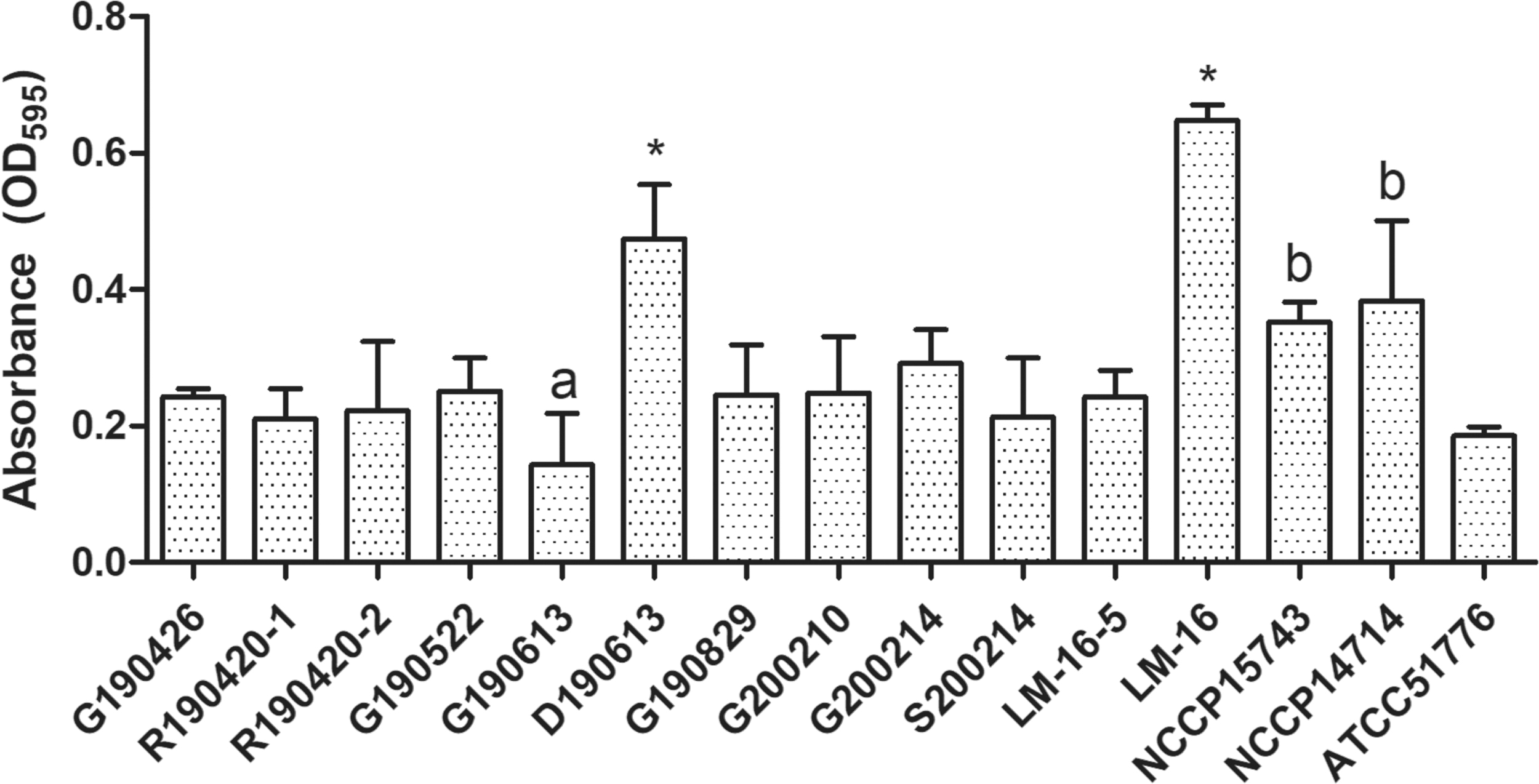
Biofilm formation of Listeria monocytogenes isolated from raw beef and slaughterhouse. R190420-1 and R190420-2: from raw beef; G190426, G190522, G190613, G190829, G200210, and G200214: from gloves of carcass-splitting workers; S200214: from carcass-splitting saw; D190613: from drain; LM-16-5 and LM-16: from beef carcass; NCCP15743 and NCCP14714: from clinical specimen; ATCC51776: a type strain. a,bThe different lowercase letters indicate significant differences at p < 0.05 using one-way ANOVA (Tukey's method). *p < 0.05 (ANOVA; Tukey's method) indicates significant differences compared with strains isolated from raw beef and slaughterhouse environment samples and collected strain samples. ANOVA, analysis of variance.
Antimicrobial susceptibility of L. monocytogenes
Antimicrobial susceptibility/resistance of the 15 isolates of L. monocytogenes, to ampicillin, chloramphenicol, ciprofloxacin. and other antimicrobials are shown in Table 2. Antimicrobial agents against which resistance was indicated were penicillin (8/15, 53.3%), and tetracycline (2/15, 13.3%). Interestingly, antimicrobial resistance was not observed in isolates from raw beef, but antimicrobial resistance was observed in most strains isolated from the slaughterhouse environment, except for G190613, LM16–5. None of the strains showed multiple drug resistance.
Analysis of Antimicrobial Susceptibility of Listeria monocytogenes Isolates Using the Disc Diffusion Test
AMP, ampicillin; C, chloramphenicol; CIP, ciprofloxacin; CN, gentamicin; E, erythromycin; K, kanamycin; P, penicillin; R, resistant; RD, rifampicin; S, susceptible; SXT, sulfamethoxazole–trimethoprim; TE, tetracycline; VA, vancomycin.
Discussion
In Korea, the level of pathogenic microbial contamination of raw beef was low, both at the point of slaughter and of consumption (Park et al., 2002). Our previous study showed that the presence of L. monocytogenes in raw beef in Korea depended on processing temperature, processing area, and market type (Kang et al., 2018).
In this study, the prevalence of L. monocytogenes was higher in the slaughterhouse environment (8/134; 6.0%) than that in raw beef (2/300; 0.7%). However, some studies have reported that raw beef would lack the nutrients needed for the growth of L. monocytogenes, and that the low pH (5.5–5.7) and storage conditions of 4°C or lower would inhibit the growth of L. monocytogenes immediately after slaughter (Shelef et al., 1989; Grau et al., 1990).
Long-term survival of L. monocytogenes in raw meat and in the environment imposes a higher risk of bacterial transfer to food for consumption, and therefore a higher risk of human exposure to the pathogen (Lambertz et al., 2013). Since the isolates from raw beef and slaughterhouse environment samples coincided temporally and geographically, in this study, PFGE clarified whether the sample types isolated were potential sources of contamination. Some isolates from the gloves of carcass-splitting workers in the slaughterhouse had the same PFGE type over a few months (Fig. 2). Interestingly, comparison strain LM-16, isolated from the beef carcass in 2006, was 100% genetically identical to strains on the gloves of workers (G190426, G190829, and G200210). In addition, Miettinen et al. (2001) found that isolates from the slaughterhouse environment (skin-removing machine) and from retailed raw beef 2 years later harbored the same PFGE type. It is possible that at least some of these strains are adapted to long-term survival in a food processing environment (Leong et al., 2014).
L.monocytogenes can attach and produce biofilms, a key factor in its persistence in food processing facilities; once established, it can act as a permanent source of environmental contamination and dispersion, leading to cross-contamination (Bonaventura et al., 2008; Bonsaglia et al., 2014). L. monocytogenes biofilms have been isolated from the surfaces of food processing equipment and working environment (Miettinen et al., 1999; Lunden et al., 2000; Tompkin, 2002). Lin et al. (2006) showed that L. monocytogenes could be transferred from the cutter to meat. In our study, L. monocytogenes was isolated from splitting saws, drains, and gloves. D190613 and LM-16 showed significantly higher ability to form biofilms than other strains (p < 0.05). There was no significant difference in biofilm-forming ability between raw beef isolates and the slaughterhouse environment isolates, except in the drain isolate D190613. Serotypes 1/2c and 4b have a higher ability to form biofilms on stainless steel, and biofilm formation and adhesion may be strain specific (Norwood and Gilmour, 1999; Kramarenko et al., 2013; Weiler et al., 2013). Chen et al. (2016) reported that persistent detection of L. monocytogenes during sampling periods may be attributed to the sanitizing methods used in the slaughterhouse that failed to remove contamination, probably because of improper sanitary procedures.
A survey of Korean livestock farms revealed penicillin to be the most used antimicrobial from 2009 to 2018 (APQA, 2019). Oh et al. (2018) reported that over 93.3% of L. monocytogenes isolates from slaughterhouse environments in Korea were resistant to penicillin. Similarly, L. monocytogenes isolates from slaughterhouse environment samples were frequently resistant to penicillin, probably because of using penicillin in veterinary practices to control infections in animals.
Conclusion
Our study indicates that contamination of L. monocytogenes in beef could consistently occur during beef processing in slaughterhouse environments through items, including gloves, splitting saws, and drains. PFGE showed that strain LM-16, isolated from the beef carcass in 2006, was genetically homologous to G190426, G190829, and G200210 isolated in this study. These results suggest that LM-16 has been present continuously in biofilms in the same slaughterhouse since 2006. Given the small sample and isolate size, our correlations of sample source, biofilm formation ability, and serotype may not be conclusive. Frequent monitoring and additional studies are needed on the epidemiology and antimicrobial resistance of L. monocytogenes strains in raw beef and slaughterhouse environments.
Footnotes
Author Contribution Statement
Y.-S.J.: Conceptualization, formal analysis, investigation, methodology, technique, writing review and editing; J.-S.M.: Technique, methodology, review and editing; H.J.K.: Technique, methodology; D.B.: Review and editing; K.-H.S.: Review and editing, supervision.
Acknowledgments
The authors would like to thank Hye-Young Youn, Hyeon-Jin Kim, Kwang-Young Song, and Yeonjin Kim for assistance with the technique.
Disclosure Statement
No competing financial interests exit.
Funding Information
This article was supported by Konkuk University Researcher Fund in 2019.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.