
Abstract
Campylobacteriosis is one of the most common bacteria causing human gastroenteritis. Poultry is a major reservoir of Campylobacter spp. as well as the main source of transmission. Due to the increased occurrence of campylobacteriosis, poultry slaughterhouses are under pressure to deliver carcasses with low contamination. However, a few studies have been carried out to evaluate Campylobacter contamination of broiler carcasses in Brazilian slaughter lines. Therefore, in this study, we aimed at detecting and quantifying the thermotolerant Campylobacter spp. at different stages of the poultry slaughtering process. The samples were collected from 12 points in three slaughterhouses in southern Brazil, at an interval of 12 months, and were tested for Campylobacter spp. by conventional microbiological technique, the most probable number, and real-time PCR. A total of 432 samples were analyzed. The majority of strains belonged to Campylobacter jejuni (92%), and the flock positivity among the three techniques was similar in most cases. Campylobacter was detected in all slaughtering stages. Although contamination has remained similar (p > 0.05) throughout almost all the slaughter process, evisceration seemed to be an important source of contamination. Our results reinforce the idea that the final carcass quality after the slaughtering process is directly influenced by the level of contamination of the broiler flocks on arrival at the processing plant.
Introduction
Campylobacteriosis is one of the most common human bacterial gastroenteritis and it is caused by thermophilic Campylobacter spp. (CDC, 2021; EFSA and ECDC, 2021; Rodrigues et al., 2021; WHO, 2021). Poultry is considered as a major reservoir and source of transmission to humans (Humphrey et al., 2014), and campylobacteriosis is usually associated with the handling, preparation, and consumption of raw and undercooked chicken meat or cross-contamination by these products (Silva et al., 2011).
Campylobacter spp. can be recovered from poultry slaughterhouses, from both processing equipment and plant environments. After their introduction into the slaughter line, it can be spread to the poultry meat, despite good hygiene, especially after evisceration or from dirty surfaces (Cisco et al., 2017; Zhang et al., 2018). Carcass monitoring for Campylobacter during slaughter process may help the development of indirect measures, complementary of best practices that can reduce the contamination. However, a few studies have been carried out to evaluate Campylobacter contamination of broiler carcasses in Brazilian slaughter lines (Borges et al., 2020).
Campylobacter spp. are fastidious bacteria that require specific culture media and incubation conditions (Platts-Mills et al., 2021). Traditional techniques, such as isolation and most probable number (MPN), are time-consuming, cumbersome, and expensive (On, 2001). Therefore, in recent years, the molecular techniques have been improved and used as alternatives for such programs.
Real-time PCR (qPCR), for example, has been described as one of the most sensitive methods for the detection of Campylobacter spp. (Reis et al., 2018). However, the routine use of the qPCR is particularly difficult in developing countries, where it represents a major cost that cannot be supported by all laboratories.
In this study, we aimed at detecting and quantifying the thermotolerant Campylobacter spp. at different stages of the poultry slaughtering process in three establishments in southern Brazil by conventional microbiology, qPCR, and MPN. In addition, we also compared the three techniques evaluated.
Materials and Methods
Experimental design
All experiments in this study were conducted in accordance with Brazilian ordinance no. 210 (Brasil, 1998). This research was approved by the Committee on Ethics in the Use of Animals by the Universidade de Passo Fundo under approval number 034/2014.
Sample collection was performed via a cross-sectional observational study by using convenience sampling. The samples were collected at an interval of 12 months and were carried out in three slaughterhouses owned by three different companies (A, B, and C) located in the Rio Grande do Sul state in southern Brazil. All slaughterhouses were under the authority of the Federal Inspection Service. A total of four visits per slaughterhouse were conducted, totaling 12 visits and 12 different flocks were sampled. The slaughterhouse characteristics are described in Table 1.
General Characteristics of the Three Slaughterhouses Selected for This Study
All flocks were also tested for Salmonella spp. from cloacal swabs following the Brazilian government official methodology (Brasil, 1995), and all presented negative results. The water supply of each establishment was also tested for Campylobacter spp. to avoid external contamination. All samples presented negative results.
Sample selection
Twelve collection points were selected according to the sanitation standard operating procedures: cloacal swabs at the reception of the broilers at the slaughterhouses, swabs of transport crates (before and after the cleaning and disinfection processes), broiler carcasses through the slaughter process (before scalding, after scalding, after defeathering, after evisceration, after initial wash, after the final wash, and after the chiller), pre-chiller water, and chiller water. Sampling was always carried out during the first slaughter shift, and samples were collected in triplicate at each establishment.
A total of 36 samples were collected per flock, and at the end of the sampling period, a total of 432 samples were analyzed from the three establishments. After collection, all samples were transported to the laboratory in isothermal boxes and refrigerated with ice. The time taken between transportation to the laboratory and the beginning of laboratorial analysis was up to 2–4 h.
For the cloacal swabs, three pools per flock were collected, and each pool included 50 swabs. Each swab was used for two broilers, with a total of 300 sampled animals from each flock. The swabs were collected at the reception of the slaughterhouse and immediately inoculated into bottles containing 50 mL of Brucella broth (Oxoid, Basingstoke, UK).
Transport crates were randomly selected and identified with seals for control before washing. Commercial sponge sticks with neutralizing buffer (3M, Maplewood, MN, USA) were used throughout the entire crate extension both before and after the cleaning and disinfection process. The sponges were packed in sterile bags containing 50 mL of Brucella broth.
For water samples, 50 mL aliquots were collected and aseptically stored in sterile bottles.
The carcasses were randomly selected, stored in sterile plastic bags, and identified with seals. In the laboratory, each carcass was rinsed with 400 mL of buffered peptone water (Oxoid) and manually shaken in a vigorous way for 30 s (Sierra-Arguello et al., 2021). The contents were aseptically collected and transferred into sterile bottles.
Finally, aliquots of 1.5 mL were collected from each sample and frozen at −20°C for qPCR analysis.
Detection of Campylobacter spp. by conventional isolation
Detection of Campylobacter spp. by conventional isolation followed the methodology described by Sierra-Arguello et al. (2021). An aliquot (1 mL) of each sample (swabs, water, and carcasses) was homogenized in Bolton broth (9 mL) (Oxoid) supplemented with antimicrobials (cefoperazone 20 mg/L, vancomycin 20 mg/L, trimethoprim 20 mg/L, cycloheximide 50 mg/L; SR0183, Oxoid) and incubated under microaerophilic conditions by using a gas tank with a mixture (10% CO2, 5% O2, and 85% N2) for 48 h at 41.5°C.
After incubation, 100 μL of the suspension was spread onto an acetate membrane with 0.65 μm pores (Sartorius, Gottingen, Germany) and placed on the surface of a modified charcoal cefoperazone deoxycholate agar (mCCDA) (Oxoid) plate with selective supplements (cefoperazone, 32 mg/L; amphotericin B, 10 mg/L; SR0155, Oxoid). The plate was incubated under microaerophilic conditions for 48 h at 41.5°C.
Subsequently, the suspected colonies were seeded in blood agar plates (Oxoid) supplemented with 5% sterile defibrinated sheep blood (Laborclin, Curitiba, Brazil). Campylobacter spp. colonies were examined under phase-contrast microscopy for typical movement and morphology (Fonseca et al., 2006). Colonies were preliminarily characterized up to the genus level based on their ability to hydrolyze hippurate and indoxyl acetate, and on catalase reaction (Public Health England 2018). Campylobacter spp. (C. jejuni and C. coli) were determined by multiplex PCR (Perdoncini et al., 2015). The primer sequences, amplicon size, and controls are described in Supplementary Data.
MPN assay
The MPN method was used to quantify Campylobacter spp., as described by Silva et al. (2010), in triplicate, in Bolton supplemented with antimicrobials and in mCCDA. The counting was carried out according to the protocol proposed by Blodgett (2006), and the limit detection is 3 MPN/mL for three tube MPN series. The results were expressed as MPN/mL and then transformed into log10 MPN/mL.
Real-time PCR
qPCR was used to detect and to quantify Campylobacter spp., and this followed the methodology described by Borges et al. (2020). An aliquot (1.5 mL) of each sample was used for DNA extraction. DNA was extracted by using the PrepMan Ultra Sample Preparation Reagent (Qiagen, Manchester, UK). Amplification was performed with the Mericon Campylobacter triple Kit (Qiagen) in a Rotor-GeneQ real-time PCR system (Qiagen).
The reactions included a total volume of 20 μL, containing 10 μL of reconstituted mericon assay master mix and 10 μL of genomic DNA. Amplification was carried out at 95°C for 5 min, followed by 40 cycles at 95°C for 15 s, 60°C for 15 s, and 72°C for 10 s. According to the manufacturer's protocol, this kit can detect up to 10 copies of Campylobacter DNA in a reaction (Qiagen, 2015).
The standard curve was plotted against the reference strain of Campylobacter jejuni subsp. jejuni ATCC 29428. Colonies cultivated on mCCDA were inoculated in the Brucella broth and incubated at 42°C overnight under agitation at 4 × g. The optical density (OD 600 nm) was measured by using a nanospectrophotometer (Implen, Westlake Village, CA, USA).
Then, the material was centrifuged at 8,000 3 × g for 5 min. The supernatant was discarded, and the pellet was suspended in phosphate-buffered saline (PBS; Laborclin). The OD was adjusted to 1.0, and 10 × serial dilutions were made in PBS. Then, each dilution was plated on mCCDA and incubated at 42°C for 24 h to confirm the cell viability. DNA was extracted by using a Wizard Genomic DNA Purification kit (Promega, Madison, WI, USA).
From the extracted DNA, a 3 μL aliquot was separated into microtubes containing 27 μL of RNase-free water for serial dilutions (10−1 to 10−10). A total of 1 μL of each dilution was transferred to 10 microtubes in duplicate with 9 μL of ultrapure water and 10 μL of reaction mix. The negative control was prepared in a microtube containing 10 μL of reaction mix and 10 μL of ultra-pure water, whereas the positive control consisted of 10 μL of reaction mix and 10 μL of positive control DNA. The final results are expressed as colony-forming unit (CFU)/mL and then transformed into log10 CFU/mL.
Statistical analysis
All statistical analyses were performed by using the SPSS Statistics version 22.0 (IBM, Armonk, NY, USA), with a significance level of 5%. The t-test was used to compare the Campylobacter spp. quantification by MPN and qPCR. Fisher's exact test was used to compare the flock positivity among conventional isolation, MPN, and qPCR.
Spearman's rank-order correlation (rho) was used to measure the strength of the association among sources of isolation. The kappa index (Landis and Koch, 1977) was used to evaluate the concordance between conventional isolation and MPN in the detection of flock positivity.
Results
The results of Campylobacter spp. detection are presented in Table 2. Most of the strains (92%) detected by conventional isolation were characterized as C. jejuni. One strain (isolated from crates before cleaning and disinfection) was identified as C. coli. Five samples (cloacal swab, carcass after defeathering, carcass after final wash, carcass after chiller, and pre-chiller water) presented a combination of two species (C. jejuni and C. coli).
Qualitative Data on Campylobacter spp. in the Three Slaughterhouses According to the Source of Isolation
Different lowercase letters in the same line indicate significant differences (p < 0.05) in the detection of Campylobacter spp. by conventional microbiology, MPN, and qPCR in the analyzed flocks (n = 12) (Fisher's exact test; adjusted p-value = 0.0169). Different capital letters in the same column indicate significant differences (p < 0.05) in the detection of Campylobacter spp. among different sources of isolation (n = 12), within the same technique (Fisher's exact test; adjusted p-value = 0.0042).
MPN, most probable number; qPCR, real-time PCR.
Flock positivity among the three techniques was similar in most cases. Qualitative analysis showed significant differences (p = 0.0361) between MPN and qPCR for crates (before cleaning and disinfection), with a higher number of positive flocks detected by MPN than qPCR. For the other sources of isolation, no significant differences (p > 0.05) were found among the three techniques.
Thus, in this study, most of the results of MPN and qPCR may be considered equivalent. The comparison of flock positivity among sources of isolation was similar (p > 0.05) for all sources of MPN. Conventional isolation showed significant differences (p = 0.0033) between carcasses after evisceration and carcasses both before and after scalding. Finally, qPCR showed significant differences (p = 0.0028) between chiller water and cages before cleaning and disinfection processes, and between chiller water and carcass after scalding.
Quantification analysis indicated a significant difference (p < 0.05) in Campylobacter spp. counts between MPN and qPCR for carcasses after chiller, where the MPN value of detection was higher than that of qPCR (Table 3). Comparison of Campylobacter quantification among the sources of isolation was similar (p > 0.05) for all sources of qPCR.
Quantitative Data on Campylobacter spp. in the Three Slaughterhouses According to the Source of Isolation
Different lowercase letters in the same line indicate significant differences (p < 0.05) in the quantification of Campylobacter spp. by MPN and qPCR (t-test). Different capital letters in the same column indicate significant differences (p < 0.05) in the quantification of Campylobacter spp. among different sources of isolation (n = 12), within the same technique (t-test; adjusted p-value = 0.0042).
CFU, colony-forming unit; MPN, most probable number; qPCR, real-time PCR; SD, standard deviation.
For MPN, significant differences (p < 0.05) were found between carcasses after evisceration and carcasses before and after scalding (p = 0.0094 and p = 0.0061, respectively). Although contamination has remained similar (p > 0.05) throughout almost the entire slaughter process, Figure 1 shows the oscillation in Campylobacter spp. quantification over the slaughtering stages.
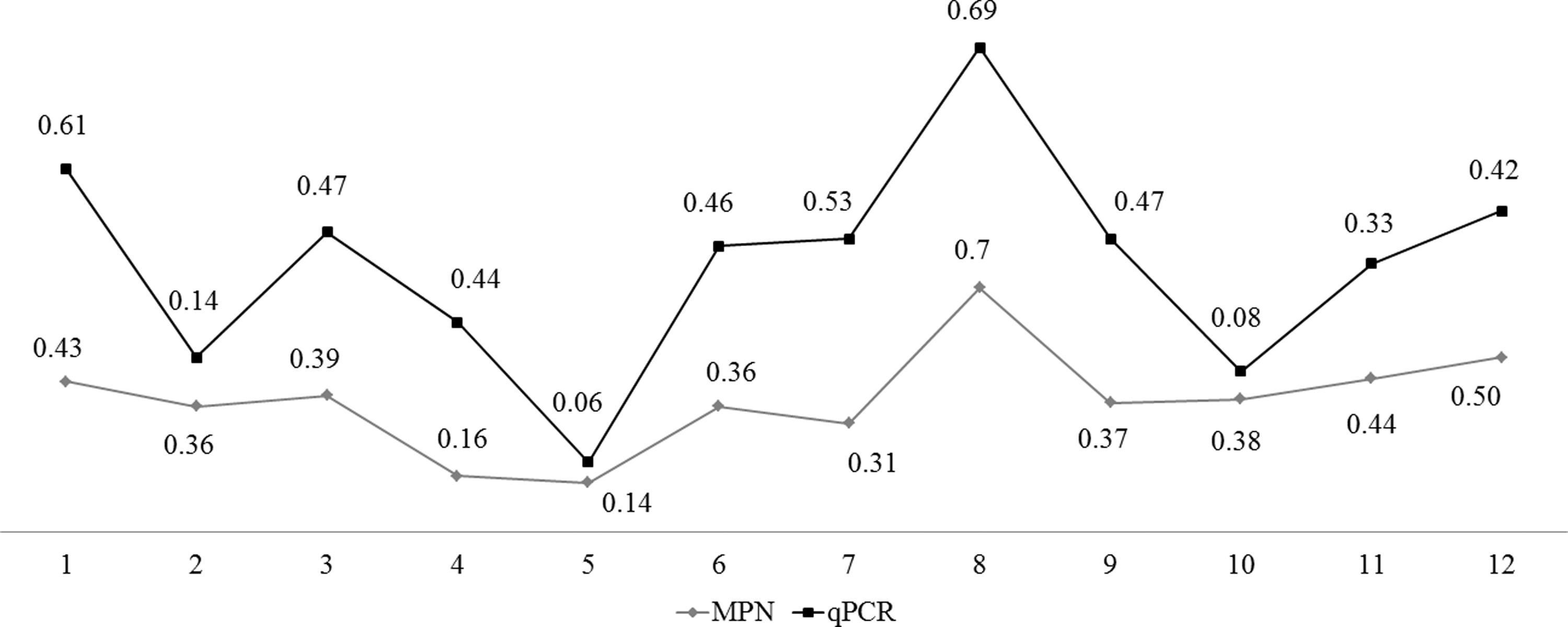
Oscillation in Campylobacter spp. quantification over the slaughtering stages, according to qPCR and MPN. Collection points: cloacal swab (1), cages before cleaning and disinfection process (2), cages after cleaning and disinfection process (3), carcass before scalding (4), carcass after scalding (5), carcass after defeathering (6), carcass after initial wash (7), carcass after evisceration (8), carcass after final wash (9), carcass after chiller (10), pre-chiller water (11), and chiller water (12). MPN, most probable number; qPCR, real-time PCR.
Peaks of contamination were observed at reception of broilers (cloacal swabs), cages after cleaning and the disinfection process, carcass after evisceration, and water from chiller tank. The peaks indicate possible “point of attention” to the establishments.
For the MPN, Spearman's rank-order correlation showed a high and positive correlation (p < 0.05) for (1) cloacal swab (carcasses after initial wash, after evisceration, after final wash, and after chiller), (2) crates before cleaning and disinfection processes (carcasses after initial wash, after final wash, and after chiller, pre-chiller water, chiller water), (3) crates after cleaning and disinfection processes (carcasses before and after scalding, chiller water), (4) carcass before scalding (carcass after scalding), (5) carcass after initial wash (carcass after final wash), carcass after evisceration (carcass after final wash), (6) carcass after evisceration (carcasses after final wash and after chiller), (7) carcass after final wash (carcass after chiller), (8) pre-chiller water (chiller water, carcass after chiller), and (9) chiller water (carcass after chiller).
However, for qPCR, only two correlations were observed: (1) crates before cleaning and disinfection processes (carcass after initial wash) and (2) chiller water (carcass after chiller). The complete results of Spearman's rank-order correlation are shown in the Supplementary Data.
To measure the agreement between conventional microbiology and MPN results, a kappa test was applied. The results showed almost perfect agreement for chiller water (κ = 0.833; p = 0.003, concordance 91.7%), and substantial agreement for pre-chiller water (κ = 0.676, p = 0.013, concordance 100%), carcass after scalding (κ = 0.750, p = 0.007, concordance 100%), and carcass after the initial wash (κ = 0.667, p = 0.014, concordance 100%). Other sources of isolation presented fair or moderate agreement.
The positive flocks according to the establishment of isolation are listed in Table 4. Statistical analysis was not conducted in this case because of the lower number of flocks per establishment. However, it is possible to note that contamination of carcasses after evisceration was a critical point for all establishments, whereas carcasses before and after scalding seemed to be less important.
Campylobacter spp. Detection According to Establishments (A, B, and C)
Total number of positive flocks per establishment.
Discussion
The data on Campylobacter infections in Brazil are still insufficient. The underdiagnosis and underreporting of campylobacteriosis are mainly associated with the difficulty of isolation and the expensive supplies that are necessary for bacterial cultivation. In addition, there is no internal monitoring program adopted by Brazilian poultry companies or government authorities, which probably justifies the lack of national data (Borges et al., 2020).
C. jejuni was the most frequent species (92%) detected by conventional microbiology. A combination of C. jejuni and C. coli was also detected. This result was expected, since C. jejuni is the most common species isolated from poultry meat (Skarp et al., 2016).
The three techniques tested in this study present differences in the requirements of cost-effectiveness, ease of execution, and execution time. However, flock positivity was similar among the three techniques, and there were significant differences between MPN and qPCR only for crates before cleaning and disinfection. Similarly, quantification analysis showed significant differences between MPN and qPCR for carcasses after the chiller.
In both cases, qPCR presented lower results. This result was not expected, since qPCR is one of the most powerful technologies in molecular biology and presents a more effective detection of microorganisms in food (Reis et al., 2018). Previous studies have shown that qPCR detection was higher than detection by conventional microbiology and by MPN (Borges et al., 2019).
A possible reason for the results found in this study is the selected kit. The mericon Campylobacter triple kit qPCR detection limit found by other research was higher than that reported by the manufacturer (Kingsbury and Soboleva, 2020). Other authors have also found lower detection by qPCR when using this kit (Borges et al., 2020; Kingsbury and Soboleva, 2020). Thus, this kit does not seem to be an appropriate alternative to the culture method in this study.
The lower detection by qPCR can be attributed to the sample type, which may present components that are potentially inhibitory to PCR reactions, such as biological materials (feces, blood, and fat) and active ingredients used in the cleaning and disinfection process, including chlorine, of slaughter plants and equipment, even with the use of a DNA extraction kit that allows the reduction of contaminants (Schrader et al., 2012; Borges et al., 2020). It is important to note that qPCR sensitivity seemed to be higher in samples from chiller water and carcasses after chilling because of the smaller concentration of inhibitors.
Ceca, colon, and cloaca are the primary sites of colonization, and infected broilers carry a very high Campylobacter load in their gastrointestinal tract (Perez-Arnedo and Gonzalez-Fandos, 2019). Thus, positivity in cloacal swabs indicates that the poultry arrives contaminated at the slaughterhouse and the infection occurred on the farm (Borges et al., 2020). In this study, 2 of the 12 flocks arrived negative and remained negative during the slaughtering process. Two flocks arrived negative, but they presented at least one further stage positive for Campylobacter spp. Eight flocks were positive during the entire process.
Crates can be a reservoir for microorganisms, including Campylobacter spp., and thus represent a risk of infection for susceptible broilers (Slader et al., 2002). Because they are reused among flocks from different farms, crates can be considered a potential source of contamination for negative flocks (Hastings et al., 2011). Thus, cleaning and disinfection processes are crucial to avoid cross-contamination among flocks and transportation trucks.
In this study, eight flocks (58.3%) presented contamination by conventional microbiology in crates before cleaning and disinfection processes, but only three of them were negative after this process. The other five remained contaminated, indicating an ineffective process. Even worse, one flock presented a negative crate before the cleaning and disinfection process, and after the process, the same crate became positive for Campylobacter spp. Our results indicate that, in most cases, the cleaning and disinfection processes of crates in poultry slaughterhouses were not effective and may contribute to carcasses contamination during the slaughter process.
Carcasses at all stages of processing were contaminated with Campylobacter spp. Although contamination did not present a statistically significant difference throughout the slaughter process, Figure 1 has shown that some steps may cause oscillation in Campylobacter quantification. On the one hand, some processes, such as scalding and cooling (pre-chiller and chiller), seemed to have an effect on the reduction of contamination.
The internal controls and monitoring programs may allow a significant reduction in contamination. An example is the control temperature and pH of the scalding process, because these factors play an important role in minimizing contamination (Duffy et al., 2014). On the other hand, processes such as defeathering and evisceration may be responsible for an increase in contamination in slaughterhouses. In fact, according to MPN results, evisceration presented a higher contamination than carcasses both before and after scalding, for example.
The amount of Campylobacter detected after the evisceration process could indicate cross-contamination with gastrointestinal content. This is especially important when considering the Spearman's correlation analysis, which indicates that flocks with higher contamination in cloacal swabs are more likely to present greater contamination after processes, such as evisceration.
Evisceration is the main route of cross-contamination within slaughterhouses, as the rupture of viscera can release a large number of Campylobacter cells onto the carcass (Hue et al., 2010). However, it is noteworthy that the showers for the final wash after evisceration helped to reduce the bacterial load in the following stages, but the remaining bacteria may have contaminated the pre-chiller and chiller water (Jimenez et al., 2002).
When we analyzed each establishment individually, it was observed that Campylobacter spp. were not detected in carcasses before scalding in establishments A and C, and in carcasses after scalding in establishment A. The cleaning and disinfection process of crates seemed to be less effective in establishment B than in A and C, as the number of contaminated crates increased after the process.
However, all establishments presented problems in their processes, as some crates remained contaminated after the cleaning and disinfection processes. Although the same type of process (sprinkle) was used at all establishments, the sanitizers used were different. Both compounds are effective in reducing the bacterial contamination, but their efficacy is reduced in the presence of organic matter (Banach et al., 2020). The establishments also presented great differences in their pre-chiller and chiller tanks (pH, temperature, and chlorination).
However, these characteristics did not seem to influence the Campylobacter contamination in this study. Further analyses are needed to compare the influence of the characteristics of establishments on bacterial contamination.
Conclusion
The results of our study indicate that conventional microbiology, MPN, and qPCR present similar results and can be used to monitor the presence of Campylobacter spp. in slaughterhouses. Our results reinforce the idea that the final carcass quality after the slaughtering process is directly influenced by the level of contamination of the broiler flocks on arrival at the processing plant.
Authors' Contributions
G.P., M.J.P.G., C.T.P.S., H.L.S.M., and V.P.N. conceived and designed the experiments. G.P., Y.M.S.A., L.M.L., L.R.S., and L.B.R. performed the experiments. K.A.B. and T.Q.F. analyzed the data. G.P., K.A.B., and T.Q.F. wrote and prepared the article. All the authors critically reviewed and approved the article.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Coordination for the Improvement of Higher Education Personnel (CAPES) and the Brazilian National Council for Scientific and Technological Development (CNPq) through the concession of a scholarship to Gustavo Perdonci and Yuli Melisa Sierra Arguello. CNPq also provided financial support through the concession of research grants numbers 578124/2008-4, 303086/2013-0, and 476092/2013-2.
Supplementary Material
Supplementary Data
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.