
Abstract
Owing to the emerging resistance to antimicrobials in Salmonella Kentucky isolates around the globe, the genomic comparison of all the registered multidrug-resistant Salmonella Kentucky isolates in Serbia (five from humans, one from turkey flock, and one from meat) was done. Most of the isolates were isolated from patients returning from Egypt or Tunisia or originated from imported turkey flock and turkey meat. The comparative analysis of resistance and virulence genes was done. All isolates belonged to sequence type-ST198 and were resistant to ciprofloxacin (Cip). The resistance to Cip was mediated by target mutations of the gyrA and parC genes, which encode topoisomerase I and II, respectively. Multidrug-resistant phenotype to aminoglycosides, β-lactam antibiotics, sulfonamides, and tetracyclines was detected in five isolates. However, none of the isolates was pan-resistant to antimicrobials. The number of single nucleotide polymorphisms between isolates varied from 8 to 43 and phylogenomics revealed the genetic proximity of the human isolate 10475/11 and the turkey meat isolate 5264/14, indicating a possible meat-to-human transfer. All isolates belonged to the main Salmonella Kentucky MDR lineage, carrying the Salmonella genomic island 1 (SGI1-K) subtype. The SGI1-K of Serbian isolates showed mosaicism attributed to rapid intraclonal evolution. Many virulence factors were detected in all the isolates, including SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, and C63PI. Although Salmonella Kentucky has rarely been isolated from humans, food, and animals in Serbia, further surveillance is needed to diminish the risk of the spreading of resistant clones and their meat-to-human transmission.
Introduction
Nontyphoidal Salmonella enterica subspecies enterica (NTS) is one of the two most important foodborne and zoonotic pathogens worldwide (Soltys et al, 2021). NTS serovars have been highlighted as a significant public health threat since they are considered reservoirs of antimicrobial drug resistance genes, whereas some isolates possess an increased virulence genotype (Aviv et al, 2014; Moreau et al, 2016).
The resistance to fluoroquinolones (FQ) such as ciprofloxacin (Cip) in Salmonella Kentucky raises major concerns (Le Hello et al, 2011). Cip-resistant Salmonella Kentucky isolates were first detected in French patients, travelers from Africa during 2002–2005 (Le Hello et al, 2011; Weill et al, 2006). Extended monitoring of human and nonhuman isolates from France, England and Wales, Denmark, Nigeria, and Morocco has revealed the existence of a single FQ-resistant clone, sequence type 198 (ST198) (Le Hello et al, 2011). Besides the resistance to FQ, Salmonella Kentucky may acquire resistance genes to extended-spectrum cephalosporins and carbapenems. Poultry was identified as a major reservoir of Cip-resistant Salmonella Kentucky clone due to the frequent use of FQ for the treatment of poultry flocks in developing countries (Le Hello et al, 2011).
Virulence factors of Salmonella Kentucky isolates are of great importance (Foley et al, 2013). As demonstrated experimentally, Salmonella Kentucky strains that acquired the ColV virulence plasmid have an increased virulence capacity for poultry (Johnson et al, 2010). It was also established that certain metabolic and nonmetabolic factors are involved in achieving fitness advantage and enhancing colonization of the Salmonella Kentucky strains in the gut (Cheng et al, 2015). Besides virulence genes, Salmonella Kentucky carries specific gene clusters encoding efflux pumps and resistance to copper, silver, and arsenic (Dhanani et al, 2015).
Salmonella Kentucky has been rarely isolated from human patients in Serbia. In poultry flocks this serovar was not detected (Jovčić et al, 2020), but from one turkey flock multidrug- and Cip-resistant Salmonella Kentucky was isolated for several years until 2015. Having in mind the potential of the global spread of Salmonella Kentucky, the whole genome analysis of isolates from patients, turkey flock, and turkey meat was done, to make the genetic comparison.
Materials and Methods
Bacterial strains and antimicrobial susceptibility testing
In this research seven Salmonella Kentucky isolates from Serbia were analyzed: five isolates from humans (546/06, 69227/09, 619/10, 10475/11, and K2011/4), one from a turkey flock (9039/12) and one from turkey meat (5264/14). Serotyping of all isolates was done according to White–Kauffmann–Le Minor scheme (Grimont and Weill, 2007) in the National Reference Laboratory for Salmonella of the Institute of Public Health of Serbia.
Antimicrobial susceptibility testing was done and evaluated applying disk diffusion method as recommended by the Clinical Laboratory Standards Institute (CLSI, supplement M100, 2018). In addition, the determination of minimal inhibitory concentration (MIC) was performed by the in-house broth microdilution method. The results of the MIC testing were evaluated according to the ECDC protocol (2016) and CLSI M100 document (2018), based on which the isolates were marked as resistant or susceptible. The Escherichia coli ATCC 25922 reference strain was inoculated on each plate and used for the quality control. The types of antimicrobial disks with antimicrobial concentrations and the results of MIC analysis are listed in Supplementary Table S1.
Whole genome sequencing and genome analysis
Total genomic DNAs of seven Salmonella Kentucky strains from Serbia were sequenced using Illumina HiSeq 2500 platform by MicrobesNG service (MicrobesNG, IMI-School of Biosciences, University of Birmingham, Birmingham, United Kingdom). The genome assembly was done using De Bruijn Graph methods and the contigs shorter than 200 bp were not taken for the analysis (Peng et al, 2012). Raw reads were assembled to scaffolds with Burrows-Wheeler Aligner (Li and Durbin, 2010). Gene annotation and prediction of the open reading frames of the whole genome sequence were conducted by NCBI Prokaryotic Genome Annotation Pipeline service.
Genomic analyses of the sequenced genomes of Salmonella Kentucky were performed using services available at Centre for Genomic Epidemiology (
PHASTER platform was used for identification and annotation of prophage sequences within analyzed genomes (Arndt et al, 2016).
Draft genome sequences of the seven Salmonella Kentucky isolates have been deposited at the NCBI GenBank database under accession numbers JAHQJE000000000-JAHQJI000000000, JAHQRB000000000, JAHQRC000000000, and JAHPZY000000000. Truncated transposon Tn1721 was detected by PCR as presented in the Supplementary Table S1.
Salmonella genomic island reconstruction and phylogenomic analysis
Phylogenomic analyses were performed with the CSI Phylogeny 1.4 (single nucleotide polymorphism [SNP] detection and construction of a phylogenetic tree) software. Salmonella Kentucky ST198 isolate 201001922 was used as a reference strain.
Mobile Element Finder v1.0.3 (Johansson et al, 2021) was used to fuse the Salmonella genomic island (SGI) segments in each genome. Assemblies were queried with the Basic Alignment Search Tool-BLAST to identify which contigs contained SGI backbone and AMR genes. In addition, assemblies were queried for IS26. Contigs containing flanking IS26 sequence with SGI genes or AMR genes were hypothesized to be part of the SGI. The results were used in conjunction with the reference Salmonella genomic island 1 (SGI1-K) reference sequence (accession number AY463797) to determine which contigs could be joined together.
Results
Molecular identification of Salmonella Kentucky isolates
In silico analyses of the seven sequenced Salmonella Kentucky genomes revealed that they share an almost uniform genome size of 4.8 Mb and GC content of 52% on average (Supplementary Table S2). According to the SeqSero software, all isolates were confirmed as Salmonella Kentucky with 8:i:z6 antigenic profile, and MLST analysis resulted in the identification of all isolates as sequence type ST198.
Phylogenomics
The percentage of reference genome that was covered by all isolates was 97.06% and 4715831 positions were found in all the analyzed genomes with the reference genome size of 4858671 bp. The number of SNPs between the Serbian ST198 isolates varied from 8 to 43 (Fig. 1). Phylogenetic tree derived from the SNPs grouped 10475/11 (human) and 5264/14 (turkey meat) distinguishing those from others (Fig. 2).

Heat map indicating a number of single nucleotide polymorphisms among genomes of Serbian Salmonella Kentucky ST198 isolates derived using CSIPhylogeny 1.4 platform. SNP, single nucleotide polymorphism.

Phylogenomics of Serbian Salmonella Kentucky ST198 isolates.
Salmonella genomic islands
The presence of any SGI backbone genes was taken as evidence of SGI integration in the analyzed Salmonella Kentucky ST198 isolates from Serbia. All genomes belonged to the main Salmonella Kentucky MDR lineage, carrying the SGI1-K subtype. Some of the isolates had large deletions of the SGI backbone similar to large deletions spanning from S015 to S025 (human isolate K2011/14), from intI1 to tnpA1133 (human isolate 619/10) or from S025 to tnpA1133 (human isolate 69227/09), but still retained the core of the multidrug-resistance region between the trmE and yidY genes (Fig. 3).
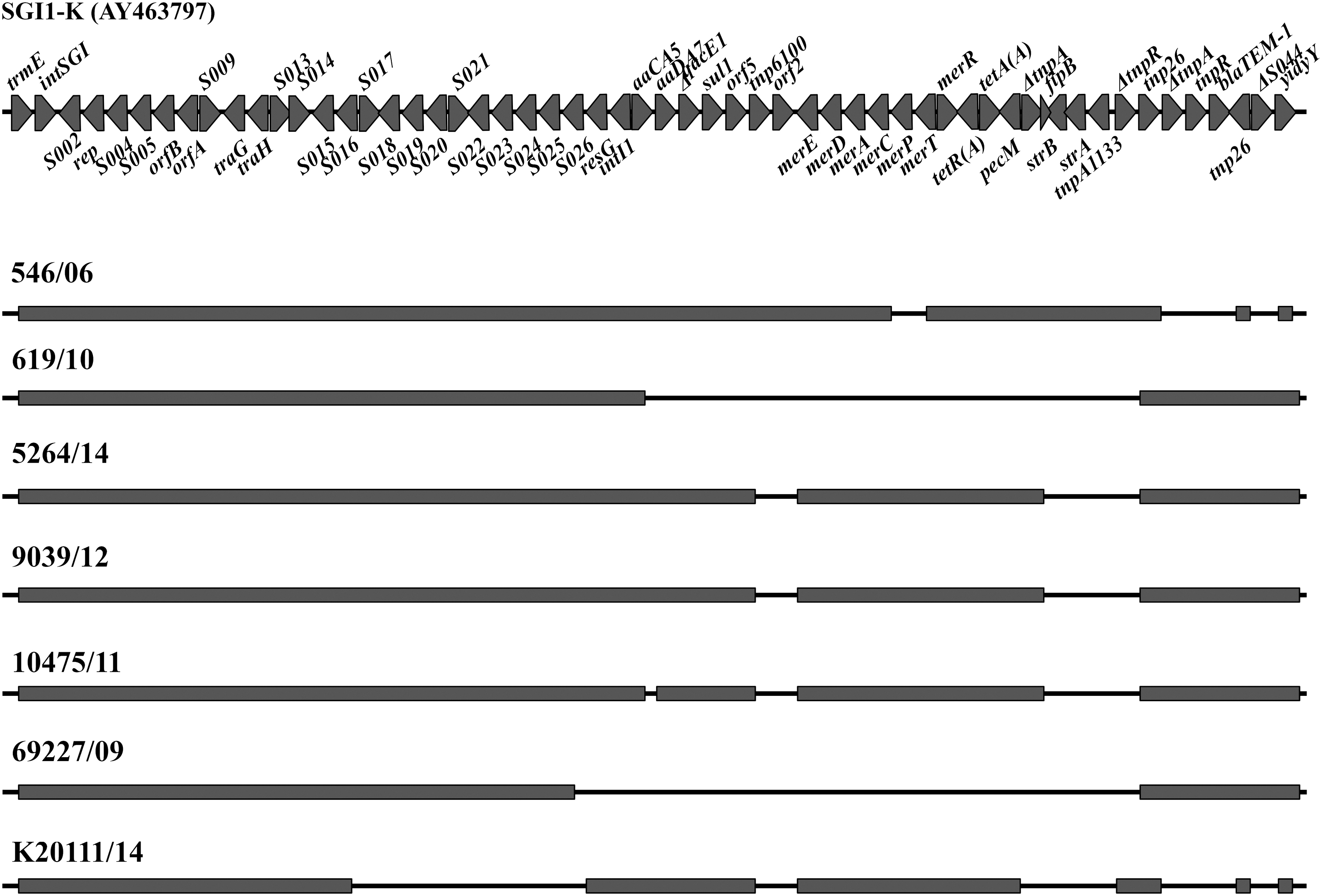
SGI variations in Serbian Salmonella Kentucky ST198 isolates. The sequence of SGI-K (GenBank accession number AY463797) is shown as a reference. Black lines indicate the deletion regions corresponding to the reference. SGI, Salmonella genomic island.
Resistome of Salmonella Kentucky ST198 isolates from Serbia
Five Salmonella Kentucky isolates were multidrug resistant: two of them (human isolates 546/06 and K2011/14) to four and three isolates (10475/11-human, 9039/12-turkey flock and 5264/14 turkey meat) to five antimicrobials of different classes. Resistance to aminopenicillin and Cip were detected in two human isolates (69227/09 and 619/10). Five isolates were resistant to aminopenicillin, gentamicin, streptomycin, sulfonamides, and tetracyclines, whereas all of the seven isolates were resistant to Cip (Supplementary Table S1). The ResFinder confirmed the presence of the genetic determinants conferring resistance to FQ, aminoglycosides, β-lactams, sulfonamides, and tetracyclines (Tables 1 and 2).
The Mutations on Topoisomerase Genes in Salmonella Kentucky ST198 Isolates
All isolates belong to the clade that emerged in Egypt from the year 2000 and spread to Europe and Asia.
The Resistance Genes in Serbian Salmonella Kentucky Isolates ST198, According to ResFinder Online Platform
Truncated transposon Tn1721 was detected with primers for Tn1721, the right arm and the tetR gene. In all the isolates following Salmonella pathogenicity islands were identified: C63PI, SPI1, SPI2, SPI3, SPI4, SPI5, and SPI9.
The most abundant mutations conferring resistance to FQ were those in the gyrA gene, resulting in Ser83Phe and Asp87Tyr changes (5/7 isolates), and in the parC gene, resulting in Ser80Ile and Thr57Ser changes (7/7 isolates) (Table 1). CARD analysis confirmed ResFinder results and broadened the spectrum of discovered resistome genes in the analyzed isolates, with the exception of the aph(3′)-Id gene in the human isolate 546/06 and aac(3)-Id and aad7 genes in the isolate 5264/14 from turkey meat that failed to be detected by ResFinder, but were detected by CARD (Supplementary Table S3).
Thus, five isolates from this collection (546/06, 10475/11, 9039/12, 5264/14, and K2011/14) had aac(3)-Id and aadA7 gene cassette, including aac(6′)-Iaa gene. Also, five isolates carried bla TEM-1, sul1, and tet(A) gene (Table 2). In addition to the number of antimicrobial efflux pumps, genes from families of RND (resistance-nodulation-division: bae, sdiA, marA, golS, rsmA, adeF, etc.), MATE (multidrug and toxic compound extrusion, mdtK), MFS [major facilitator superfamily: kpnF, kpnE, mdfA, emrB, emrR, tet(A)], and ABC (ATP-binding cassette, msbA) mutations in the glpT gene (E448K) conferring resistance to fosfomycin and in tuf gene (EF-Tu, R234F) conferring resistance to pulvomycin were found using CARD (Supplementary Table S4).
Virulence determinants in Salmonella Kentucky isolates from Serbia
The following genes encoding virulence factors have been found in Salmonella Kentucky isolates from Serbia using VFDB: the capsule, fimbrial adherence determinants, macrophage inducible genes, genes involved in Mg2+ transport, nonfimbrial adherence determinants, PhoPQ two-component regulatory system, SPI-1 and SPI-2 secretion systems, including genes encoding corresponding translocated effectors and serum resistance. In addition, stress adaptation genes coding for toxin, adherence, antiphagocytosis, and autotransporter virulence factors were found (Supplementary Table S4). All isolates were characterized by the presence of the Salmonella pathogenic islands SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, and SP-I9, C63PI (centisome 63 pathogenicity island).
Prophages and plasmids present in genomes of Salmonella Kentucky isolates from Serbia
Intact, complete prophage sequences were found in all the isolates except in K2011/14. The dominant prophage PHAGE_Salmon_vB_SemP_Emek_NC_018275 was found in five out of seven isolates. Prophage PHAGE_Salmon_118970_sal4_NC_030919 was found only in isolate 546/06 (Supplementary Table S2).
Plasmids were detected in isolates 619/10 and K2011/14. Isolate 619/10 carried Inc1-I(Alpha) plasmid that was 100% identical with the template (length 142/142 bp) from Salmonella Typhimurium plasmid R64 (GenBank accession No. AP005147). Isolate K2011/14 carried Col (pHAD28) plasmid (NODE_13_length_4223_cov_147.097412) that shared identity with 97.48% of the template (length 119/131) from S. Hadar plasmid pHAD28 (GenBank accession No. KU674895).
Discussion
In this study, it was established that high-level resistance to Cip was induced by multiple mutations on gyrA and parC genes in all isolates. It could be assumed that they do not have a dramatic impact on bacterial fitness because Salmonella Kentucky ST198 has been a well-established global clone for decades (Hawkey et al, 2019; Le Hello et al, 2013; Soltys et al, 2021). In this study, we detected the mutation Thr57Ser in the parC gene as well, which is a naturally occurring compensatory mutation in Salmonella that change its binding affinity for the quinolone (Eves et al, 2004). Resistance to aminoglycoside, β-lactam antimicrobials, sulfonamides, and tetracyclines was also prominent in most of the isolates and some of the resistance genes were detected in SGI1-K.
Chromosomal integration of the Salmonella genomic island SGI1-K had been dated in the mid-1990s, just before the increase of resistance to Cip was noted in Salmonella Kentucky strains. Independent and quick acquisition of SGI-K in several bacterial isolates in Egypt may have risen in farms practicing integrated aquaculture (Le Hello et al, 2011). The diversity of SGI1-K among Serbian Salmonella Kentucky isolates is in concordance with previously reported results and further state on rapid intraclone evolution (Chen et al, 2021). It is proposed that these differences may rise due to IS26 mediated gain or loss of the genes within the SGI1-K (Chen et al, 2021; Doublet et al, 2008; Hall, 2010).
In all analyzed Salmonella Kentucky isolates we have found chromosomally located genes aac(6′)-Iaa and aac(3)-Id, which encode aminoglycoside acetyltransferase enzymes. These enzymes have a role in modifying the 3 and 6′-amino group of aminoglycoside antibiotics (Shaw et al, 1993). The aadA7 gene cassette was determined in five isolates (546/06, 10475/11, 9039/12, 5264/14, and K2011/14). It was established, for the first time, that aac(3)-Id and aadA7 gene cassettes are inserted into the SGI1-H and confer resistance to gentamicin, streptomycin, spectinomycin, and sisomicin in the serovar Newport (Doublet et al, 2004).
Only the isolate 546/06 had aph (3″)-Ib and aph(6)-Id resistance gene that was identified in the SGI1-K backbone. In addition, two human isolates (546/06 and K2011/14) did not have the bla TEM-1B β-lactamase resistance gene in the SGI1-K due to the loss of genetic material. In five isolates (546/06, 10475/11, 9039/12, 5264/14, and K2011/14) the tetracycline-resistance gene tetA was detected by PCR on transposon Tn1721, truncated at the left arm. This transposon was frequently found in Serbian Salmonella Infantis isolates from poultry, collected during routine monitoring in 2014 (Todorović et al, 2015) and the tet(A) gene was a common resistance gene in clinical E. coli isolates from cattle and pigs (Todorović et al, 2018).
All isolates in this research harbored the SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, and C63PI Salmonella pathogenicity islands. It is postulated that SPI was acquired from plasmids or phages by the horizontal transfer mechanism (Marcus et al, 2000). Except for the SPI-9, all other SPIs from Salmonella Kentucky were also found in Salmonella Infantis isolates from poultry flocks in Serbia (Jovčić et al, 2020). The SPI-9 has been characterized as an ancient genomic island previously found in different Salmonella enterica serovars and has become stable in the host genome of Salmonella (Velásquez et al, 2016).
The distribution and abundance of virulence genes within genomes of Serbian Salmonella Kentucky isolates are uniform except for isolate number 619/10, which has, in addition, the type IV pili virulence factors. The most dominant prophage present in the Serbian Salmonella Kentucky isolates is vB_SemP_Emek (NC_018275) prophage. The closest homologues of this prophage are Salmonella Dublin SD3246, Salmonella Dublin CT_02021853 and Salmonella phage 22 of Podoviridae. Although vB_SemP_Emek belongs to P22-like virus genus it lacks homologues of P22 O1 serovar conversion genes, integrase and exicionase (Ho et al, 2012).
Interestingly, an isolate from a human patient (10475/11) and imported turkey meat (5264/14) differed in the eight SNPs. This close relationship between the isolates was further confirmed by the phylogenetic tree. Genetic relatedness between human and food isolates has been identified in Salmonella Kentucky strains in the United States by Haley et al (2019). In addition, we found that isolates from an outpatient (10475/11), turkey meat (5264/14), and turkey poults (9039/12) share 8, 11, and 13 polymorphisms and are characterized by the resistomes that resemble closely to those of the isolates from travelers outside the United States, which belong to the clade 198.2 (Haley et al, 2016, 2019).
Conclusion
This study presents the first whole-genome analysis of Salmonella Kentucky isolates in Serbia. All the isolates belonged to the sequence type ST198 and showed high resistance to Cip. From the global epidemiological point of view, the close genetic link detected between the isolate from turkey meat and a human patient is troublesome. The weak part of the research is that only multidrug-resistant isolates were included in the genetic analysis. Therefore, common clones, regardless of their resistance phenotypes, need to be identified in the future. Even though Salmonella Kentucky has rarely been isolated from humans in Serbia, it may change, since travel and trade still involve a significant risk in spreading resistant clones around the globe.
Footnotes
Authors' Contributions
Conceptualization, methodology, writing, formal analysis, and funding acquisition by B.J., M.K., and M.V. Investigation by B.J. M.K., D.T., M.M., and D.V. Writing, review, and editing by B.J., M.V., and M.K. All authors have read and agreed to the published version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Ministry of Education, Science and Technological Development of the Republic of Serbia by the Contract of implementation and financing of scientific research work Contract Nos. 451-03-68/2022-14/200031, 451-03-68/2022-14/200042, and 451-03-68/2022-14/200178.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.