
Abstract
Escherichia albertii is an emerging pathogen causing foodborne infections with diarrhea, abdominal pain, and fever. E. albertii has been isolated from various food sources, such as chicken and pork. Although many foodborne outbreaks of E. albertii have been reported, the causative food has not been identified. It is necessary to develop effective detection methods for E. albertii. Because enrichment procedure as the first step of food test is important for growing pathogens, this study aimed to develop a novel effective enrichment for E. albertii detection in food. In this study, we investigated the optimal concentration and combination of cefixime and tellurite for supplementing modified EC broth (mEC) to effectively isolate E. albertii from chicken meat. The results showed that mEC supplemented with 50 μg/L cefixime and 2.5 mg/L tellurite (CT-mEC) inhibited the growth of competitive bacteria in chicken meat but not that of E. albertii. Therefore, it was indicated that CT-mEC had strong potential to selectively grow E. albertii. In an E. albertii foodborne outbreak, CT-mEC was evaluated. E. albertii was successfully isolated from a food sample, a kind of salad, by enrichment with CT-mEC but not buffered peptone water and mEC. In this study, CT-mEC as a selective enrichment broth has been developed to detect E. albertii in chicken meat. It was demonstrated that the selective enrichment broth was effective for the efficient detection of E. albertii in food.
Introduction
Escherichia albertii is a foodborne pathogen causing diarrhea, abdominal pain, and fever in humans. E. albertii was originally recognized as an atypical strain of Hafnia alvei that expressed intimin. Isolates from children in Bangladesh with diarrhea, which were classified as H. alvei in 1991, were reclassified as E. albertii, based on their biochemical characteristics and genetic homology to Escherichia coli and Shigella flexneri. (Abbott et al., 2003; Huys et al., 2003). Recently, several foodborne outbreaks caused by E. albertii have been reported (Etoh et al., 2009; Nagaoka et al., 2016; Ooka, 2017; Tokoi et al., 2018). The prevalence of E. albertii among many species of wild birds was studied, and E. albertii has been found to have a broad host range among birds (Oaks et al., 2010; Oh et al., 2011). E. albertii has been isolated from various sources of meat, such as chicken, pork, mutton, and duck (Asoshima et al., 2015; Wang et al., 2016). It has also been isolated from water (Felföldi et al., 2010; Maheux et al., 2014; Takahashi et al., 2020). Causative foods, however, have not been identified in many foodborne outbreaks of E. albertii (Nagaoka et al., 2016; Tokoi et al., 2018).
This is because of insufficiently developed methods for detecting E. albertii, particularly inefficient selective media. On selective agar, deoxycholate hydrogen sulfide lactose agar (DHL) supplemented with xylose and rhamnose (RX-DHL), E. albertii has been effectively isolated from a chicken sample; this is because most of the E. albertii strains are unable to utilize lactose, rhamnose, sucrose, and xylose (Arai et al., 2021). The vigorous growth of bacteria from food samples often hampers the isolation of E. albertii on RX-DHL (Arai et al., 2021). Therefore, selective enrichment is necessary for further improvement of the isolation of E. albertii from food samples.
Buffered peptone water (BPW), as a nonselective medium, has been widely used for isolation of food-poisoning bacteria, including E. albertii (Maeda et al., 2015; Tokoi et al., 2018). On the other hand, Arai et al. (2021) used modified EC broth (mEC) or mEC supplemented with novobiocin (NmEC) as a selective medium for isolation of E. albertii. Therefore, mEC-based enrichment broths were used in this study.
To improve selectivity of broth, we focused cefixime and tellurite because supplement of cefixime and tellurite to tryptic soy broth (TSB) has been reported to improve detection of enterohemorrhagic E. coli (EHEC), which is a pathogen that belongs to the Escherichia genus (Zadik et al., 1993; Catarame et al., 2003). The optimal concentrations of cefixime and tellurite to promote the growth of E. albertii and inhibit competitive bacteria in food were investigated using pure culture strains and the pathogen-spiked culture of chicken meat. In addition, mEC supplemented with an optimal concentration of cefixime and tellurite was evaluated to detect E. albertii in food associated with a foodborne outbreak of the pathogen. This novel approach for the development of an E. albertii-specific selective enrichment will strengthen microbial investigation on E. albertii-contaminated food.
Materials and Methods
Bacterial strains and culture conditions
A total of 195 E. albertii strains, including EA12, EA24, and EA42, were isolated from the following sources: 105 strains from food and patients in foodborne outbreaks, 42 strains from foods and environment, 43 stains from animals, and 5 other strains. In addition, eight strains of Escherichia fergusonii (EF1-EF8), Morganella morganii-type strain (JCM1672T, Japan Collection of Microorganisms, Saitama, Japan), three strains of Proteus mirabilis (PM1-PM3), four strains of H. alvei (HA1-HA4), a strain of Enterobacter cloacae (EC1), E. coli type strain (ATCC11775T; American Type Culture Collection, Manassas, VA), a strain of EHEC (ESC425), and S. flexneri-type strain (ATCC24570T) were used.
Optimal mEC concentrations of cefixime and tellurite for selective growth of E. albertii
Three strains of E. albertii (EA12, EA24, and EA42) isolated from patients in Japan, a strain each of E. fergusonii (EF1), M. morganii (JCM1672T), P. mirabilis (PM1), and E. cloacae (EC1) were included. These bacterial strains were incubated at 37°C for 18 h in 10 mL of TSB (Oxoid, Hampshire, United Kingdom). The bacterial cultures were diluted with double-strength mEC (Nissui, Tokyo, Japan) to 106 colony-forming unit (CFU)/mL. The diluted cultures (50 μL) were dispensed into each well of 96-well plates. Cefixime (Fujisawa Pharmaceutical Co. Ltd, Osaka, Japan), potassium tellurite (Fujifilm Wako Pure Chemical Corporation, Osaka, Japan), and a mixture of the two suspended in 50 μL in distilled water were added to each well at varying final concentrations as shown in Supplementary Table S1. The plates were incubated at 42°C for 24 h. The optical density of each well was measured with Infinite 200 PRO F Plex (TECAN Group Ltd., Männedorf, Switzerland) at 590 nm. Single-strength mEC was used as the blank sample. The assay was performed in quadruplicates.
Optimal mEC concentration of cefixime and tellurite to isolate E. albertii from chicken-enriched culture
Twenty-five grams of chicken meat in each stomacher bag was homogenized with 225 mL of mEC for 1 min. Cefixime and tellurite were added to each 10 mL of the homogenates in the following three combinations: 12.5 μg/L + 0.625 mg/L, 25 μg/L + 1.25 mg/L, and 50 μg/L + 2.5 mg/L of cefixime + tellurite, respectively. Furthermore, each strain of E. albertii (EA12, EA24, and EA42) was inoculated at ∼1 CFU/mL and incubated at 42°C for 22 h. A 10 μL aliquot of the culture was streaked onto DHL supplemented with 1% (w/v)
Noncolored colonies and other colored colonies were counted as those corresponding to E. albertii and bacteria from chicken meat, respectively. DNA was extracted from three noncolored colonies on each agar plate by suspending in 100 μL of sterilized tris-EDTA buffer and heating at 100°C for 10 min, and used in real-time PCR specific for E. albertii (EA-rtPCR; Arai et al., pers. comm.). Chicken meat without inoculation with E. albertii was used as a negative control. The assay was performed in triplicates.
Growth of E. albertii and other bacterial strains in CT-mEC and CT-NmEC
A loopful of TSB culture of 195 E. albertii and 20 strains of other bacteria (Table 1) were inoculated into 10 mL of mEC and NmEC (Eiken Chemicals, Tokyo, Japan) supplemented with 50 μg/L cefixime and 2.5 mg/L tellurite (CT-mEC and CT-NmEC, respectively) and incubated at 42°C for 24 h. The turbidity of the broth was visually assessed to determine the growth of the strains.
Growth of Bacterial Strains Used in This Study in CT-mEC and CT-NmEC
Two of E. fergusonii strains grow both in CT-mEC and CT-NmEC showed robust growth in CT-NmEC compared with CT-mEC.
CT-mEC, mEC supplemented with 50 μg/L cefixime and 2.5 mg/L tellurite; CT-NmEC, mEC supplemented with 25 μg/L novobiocin and 50 μg/L cefixime, and 2.5 mg/L tellurite; mEC, modified EC broth.
Growth of E. albertii cocultured with E. coli in chicken-enriched culture
Twenty-five grams of chicken meat in each stomacher bag was homogenized with 225 mL of CT-mEC for 1 min. E. albertii (EA12, EA24, or EA42) was inoculated into each 10 mL of the homogenates with/without E. coli-type strain or EHEC at ∼1 CFU/mL each, and incubated at 42°C for 22 h. By the same procedure described above, RX-DHL inoculated with 10−5 dilution of cultures was incubated, and growing colonies were counted and confirmed as E. albertii.
E. albertii isolation from food associated with a foodborne outbreak
Eleven samples (Table 2), including 10 foods (No. 1–10) and a mixture of the 10 foods (No. 11) from an E. albertii foodborne outbreak in Otsu, in 2020 (118 cases, 0 death), were incubated at 37°C for 22 h in 9 × BPW (Oxoid). DNA was extracted from the cultures using the alkali extraction method (Hara-Kudo et al., 2016) and used for EA-rtPCR. For a second enrichment, each 0.1 mL culture positive for EA-rtPCR was inoculated into 0.9 mL of mEC, NmEC, CT-mEC, and CT-NmEC, and incubated at 42°C for 22 h. In this study, the CT-mEC and CT-NmEC were prepared with mEC, NmEC, and a mixture supplement of cefixime and tellurite (CT-supplement; Oxoid). DNA was extracted from the second cultures and used for EA-rtPCR. Ten microliters of the second culture in mEC and CT-mEC, each was streaked on RX-DHL and incubated at 37°C for 18 h. Noncolored colonies grown on RX-DHL were confirmed as E. albertii by EA-rtPCR.
Detection of Escherichia albertii in Food Samples Associated with a Foodborne Outbreak by Real-Time Polymerase Chain Reaction
BPW, buffered peptone water; Ct, cycle threshold; CT-mEC, mEC supplemented with 50 μg/L cefixime and 2.5 mg/L tellurite; CT-NmEC, mEC supplemented with 25 μg/L novobiocin and 50 μg/L cefixime, and 2.5 mg/L tellurite; mEC, modified EC broth; NT, not tested; NmEC, mEC supplemented with 25 μg/L novobiocin.
Statistical analyses
To compare the optimal concentrations of cefixime and tellurite to selectively grow E. albertii in mEC, the mean OD590 values of each bacterial culture in mEC supplemented with cefixime and tellurite against that without the reagents were statistically analyzed using the Shirley–Williams test (p < 0.05).
To compare the optimal concentrations of cefixime and tellurite to isolate E. albertii from chicken-enriched culture in mEC, the mean values of the bacterial count of each bacterial culture in mEC supplemented with cefixime and tellurite against that without the reagents were statistically analyzed by Dunnett's multiple comparison test (p < 0.05).
To compare the growth of E. albertii and competitive bacteria in chicken-enriched cultures cocultured with/without E. coli, the mean values of the bacterial count in culture with E. coli against that without the bacterium were statistically analyzed by Dunnett's multiple comparison test (p < 0.05).
Results
Optimal mEC concentration of cefixime and tellurite to inhibit the growth of bacteria, except E. albertii, from food
For this analysis, three E. albertii strains (EA12, EA24, EA42), a strain each of E. fergusonii (EF1), M. morganii (JCM1672T), P. mirabilis (PM1), and E. cloacae (EC1) were included and all seven strains used in this study grew well in mEC without cefixime and tellurite (Figs. 1–3). Therefore, the OD590 values in mEC supplemented with various concentrations of cefixime and tellurite were compared with those in mEC with cefixime and tellurite for each strain.
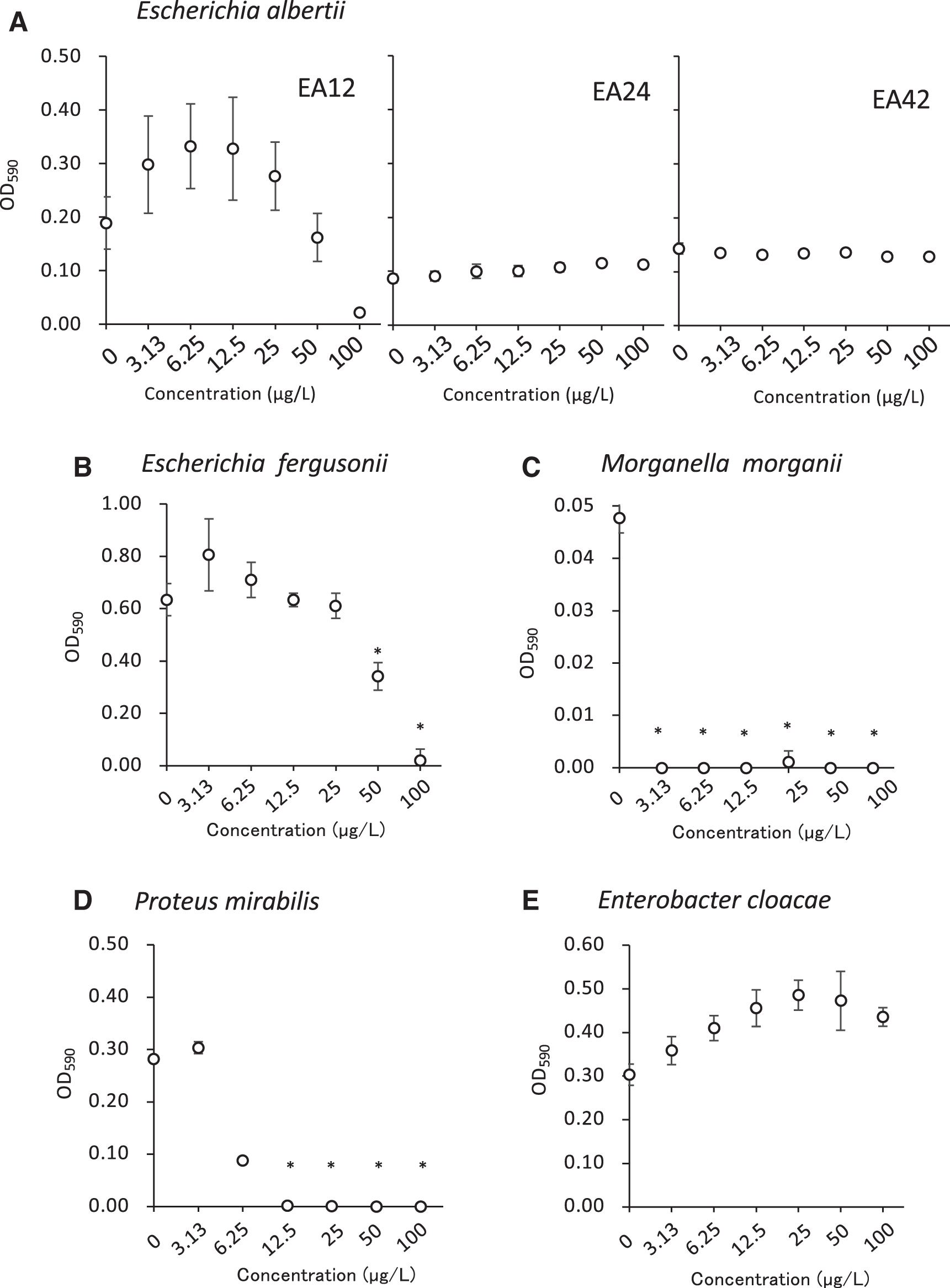
Effects of cefixime on the growth of Escherichia albertii and bacteria from food. The OD590 values of
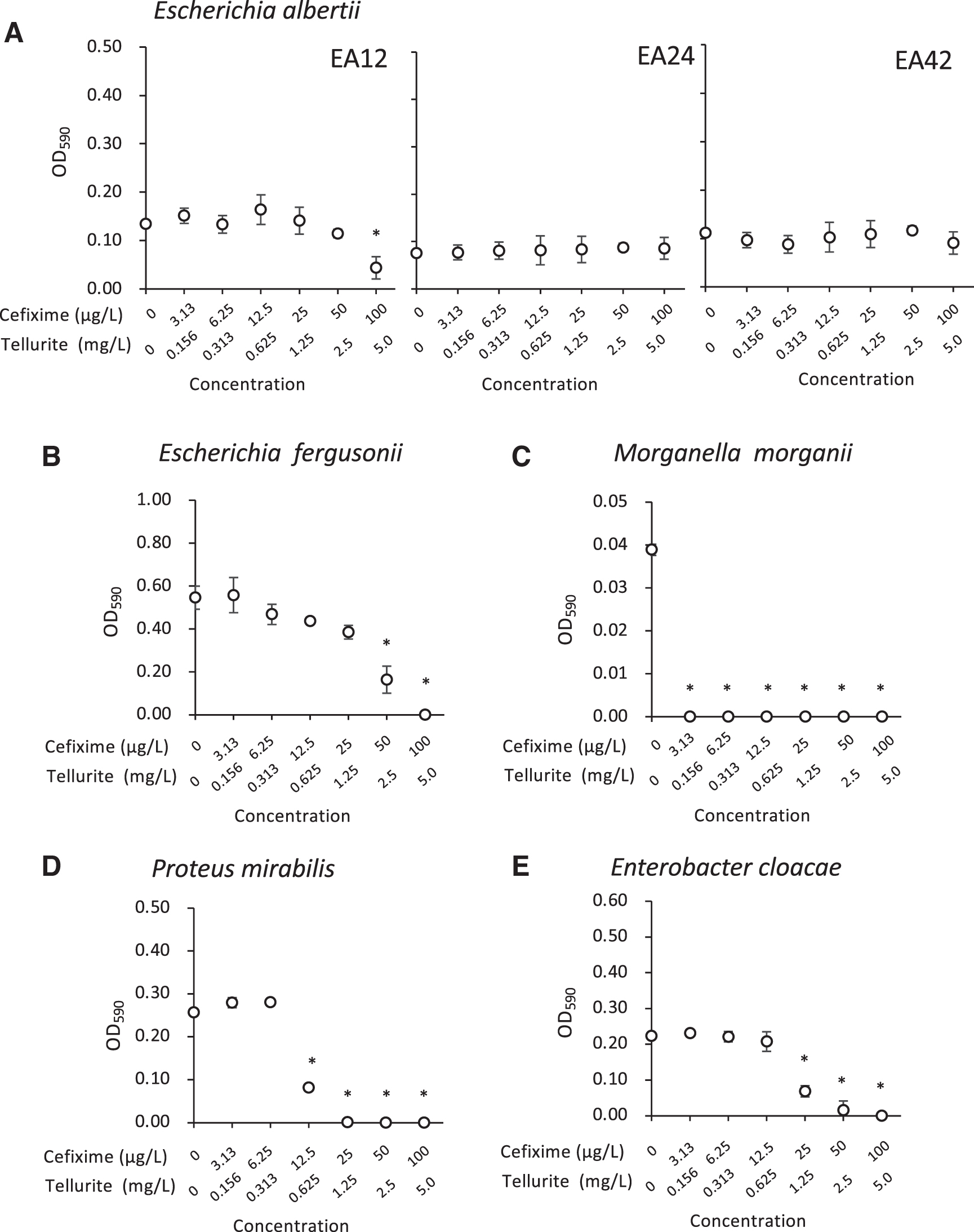
Effect of cefixime and tellurite mixture on the growth of Escherichia albertii and bacteria from food. The OD590 values of
On supplementation with cefixime, E. albertii EA24 and EA42 grew well with cefixime 3.13–100 μg/L, and E. albertii EA12 grew well with 3.13–50 μg/L cefixime but not with 100 μg/L (Fig. 1A). The growth of E. fergusonii, M. morganii, and P. mirabilis was significantly inhibited by cefixime 50–100, 3.13–100, and 12.5–100 μg/L, respectively (Fig. 1B–D). E. cloacae, however, grew well with 3.13–100 μg/L cefixime (Fig. 1E).
On supplementation with tellurite, E. albertii EA12, EA24, and EA42 grew well with 0.156–5.0 mg/L tellurite (Fig. 2A). The growth of E. fergusonii, M. morganii, and E. cloacae was significantly inhibited by 1.25–5.0, 0.156–5.0, and 0.625–5.0 mg/L tellurite, respectively (Fig. 2B, C, E). P. mirabilis grew well with 0.156–5.0 mg/L tellurite (Fig. 2D).
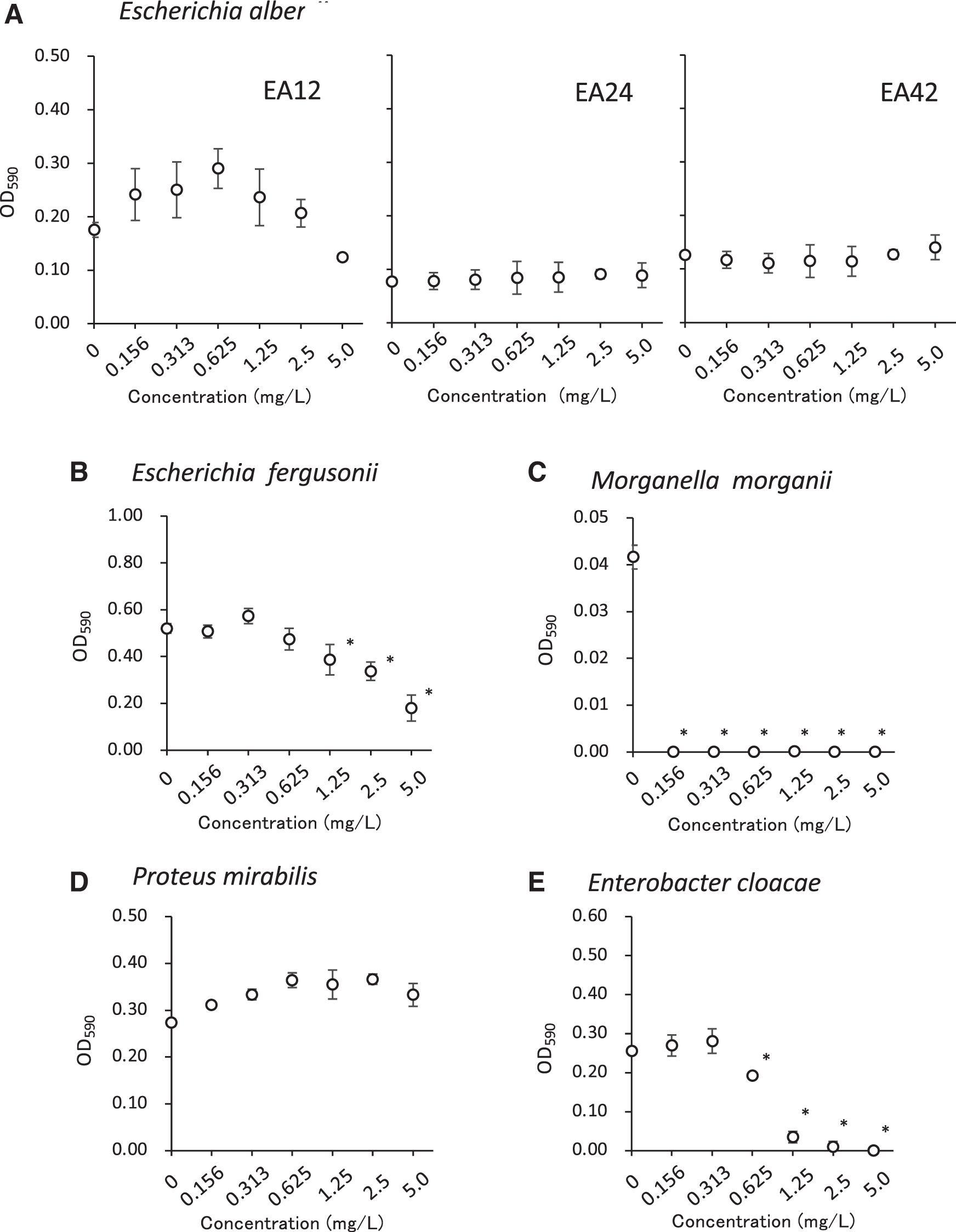
Effects of tellurite on the growth of Escherichia albertii and bacteria from food. The OD590 values of
On supplementation of mEC with combinations of cefixime and tellurite, E. albertii EA12, EA24, and EA42 grew well with 3.13–50 μg/L cefixime and 0.156–2.5 mg/L tellurite (Fig. 3A). The growth of E. fergusonii, M. morganii, P. mirabilis, and E. cloacae was significantly inhibited by 50–100 μg/L cefixime and 2.5–5.0 mg/L tellurite, 3.13–100 μg/L cefixime and 0.156–5.0 mg/L tellurite, 12.5–100 μg/L cefixime and 0.625–5.0 mg/L tellurite, and 25–100 μg/L cefixime and 1.25–5.0 mg/L tellurite, respectively (Fig. 3B–E).
Optimal concentration of selective reagents to isolate E. albertii from chicken-enriched culture
On RX-DHL-streaked culture of E. albertii-inoculated chicken, more abundant noncolored colonies suspected as E. albertii were formed with culture in mEC supplemented with cefixime and tellurite than those without the reagents (Supplementary Fig. S1A). These noncolored colonies suspected as E. albertii were confirmed as E. albertii by EA-rtPCR. Noncolored colonies, suspected as E. albertii, were not formed on RX-DHL-streaked culture of E. albertii-noninoculate chicken in mEC (Supplementary Fig. S1B).
In quantitative analysis, the bacterial population of competitive bacteria in chicken culture in mEC supplemented with 50 μg/L cefixime and 2.5 mg/L tellurite significantly decreased compared with culture in mEC without reagents (Fig. 4). The bacterial population of E. albertii EA12 and EA24 in mEC supplemented with 12.5–50 μg/L cefixime and 0.625–2.5 mg/L tellurite, and 25–50 μg/L cefixime and 1.25–2.5 mg/L tellurite, respectively, significantly increased compared with culture in mEC without reagents (Fig. 4).
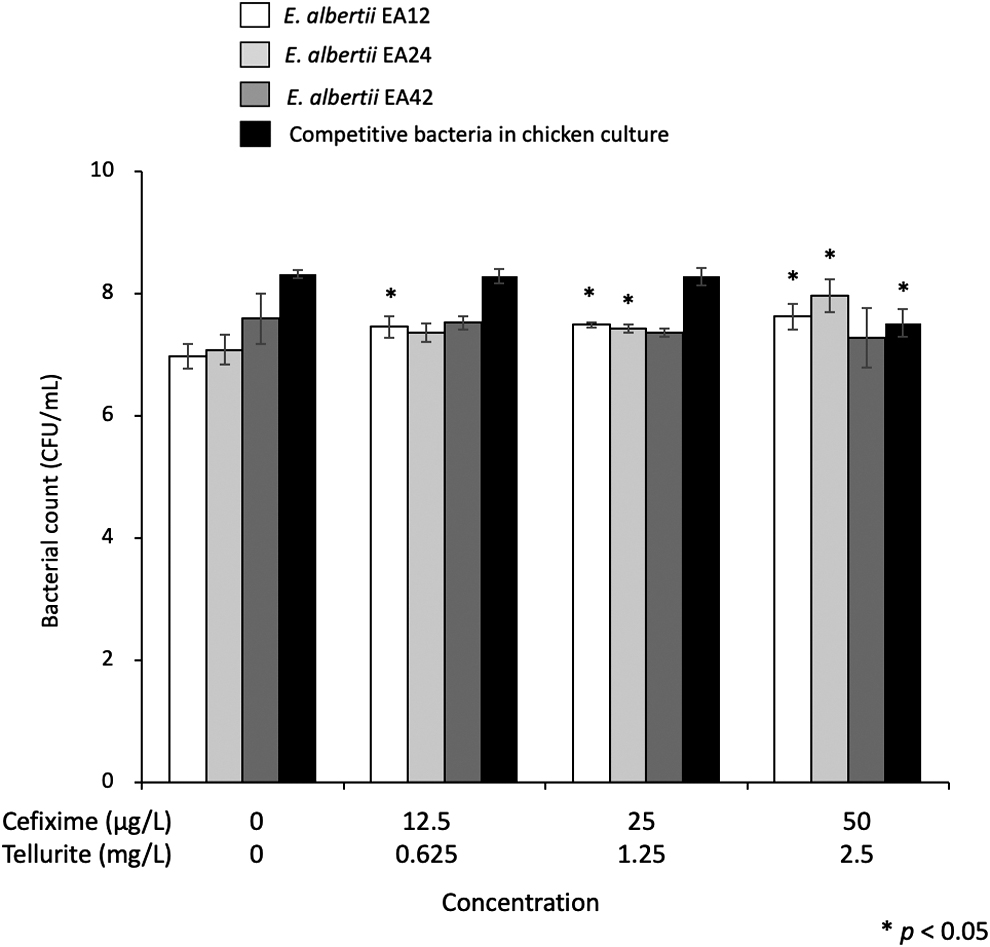
Growth of Escherichia albertii and competitive bacteria from chicken meat in mEC supplemented with combinations of various concentrations of cefixime and tellurite. Noncolored colonies and other colonies were counted as E. albertii and other bacteria from chicken meat, respectively. The bars indicate the standard deviations of the bacterial number. Statistical significance between bacterial count of culture in mEC without cefixime and tellurite and those with cefixime and tellurite at the p < 0.05 level is indicated using asterisks (*). mEC, modified EC broth.
Growth of E. albertii and other bacterial strains in CT-mEC and CT-NmEC
All 195 E. albertii strains grew well in CT-mEC and CT-NmEC (Table 1). Three out of eight E. fergusonii strains grew in CT-mEC and CT-NmEC, and other three strains did not grow in CT-mEC and CT-NmEC. The rest two strains grew in CT-NmEC but not CT-mEC. In addition, two E. fergusonii strains grown both in CT-mEC and CT-NmEC showed robust growth in CT-NmEC than that in CT-mEC. The growth of one M. morganii, three P. mirabilis, four H. alvei, one E. cloacae derived from foods, and one S. flexneri strain were inhibited in both CT-mEC and CT-NmEC. Two E. coli strains grew in CT-mEC and CT-NmEC (Table 1).
Growth of E. albertii cocultured with E. coli in chicken-enriched culture
The E. albertii populations of all strains in chicken culture in CT-mEC with the E. coli strains did not significantly change compared with that in cultures without the E. coli strains (Supplementary Fig. S2). Competitive bacterial populations also did not significantly change by coculture with the E. coli strains.
E. albertii isolation from food associated with a foodborne outbreak
The cultures of vermicelli Chinese salad (No. 9) and the mixture of foods (No. 11) in BPW were positive for EA-rtPCR (Table 2). The second culture of vermicelli Chinese salad in CT-mEC showed the lower cycle threshold (Ct) values of EA-rtPCR than those incubated in mEC, NmEC, and CT-NmEC (Table 2). As a result of streaking enriched cultures of vermicelli Chinese salad and the mixture of foods on RX-DHL, noncolored colonies suspected as E. albertii were isolated from the enriched culture in CT-mEC but not mEC. Three noncolored colonies isolated from cultures of vermicelli Chinese salad and the mixture of foods on RX-DHL were positive for EA-rtPCR.
Discussion
Isolation of foodborne pathogens from food in bacterial foodborne outbreaks is important for identifying the causative pathogen and food. Few studies have reported on the detection of E. albertii. Although we previously reported that RX-DHL is efficient in isolating E. albertii from chicken meat (Arai et al., 2021), the growth of competitive bacteria in food hampers the isolation of E. albertii. Therefore, superior selective enrichment broth is required to improve E. albertii isolation. We focused on cefixime and tellurite, particularly their optimal concentrations, to investigate a selective enrichment broth to detect E. albertii in food, and inhibit the growth of bacteria other than E. albertii, using a pure culture of bacterial strains. The growth of E. fergusonii, M. morganii, P. mirabilis, and E. cloacae were inhibited by cefixime and/or tellurite (Figs. 1 and 2), similar to a previous study (Toptchieva et al., 2003).
Furthermore, the optimal combinations of cefixime and tellurite were investigated. The growth of one of the three E. albertii strains (EA12) was inhibited in mEC supplemented with 100 μg/L cefixime and 5.0 mg/L tellurite, but not with 50 μg/L cefixime and 2.5 mg/L tellurite, which strongly inhibited the growth of E. fergusonii, M. morganii, P. mirabilis, and E. cloacae (Fig. 3). Furthermore, a combination of 50 μg/L cefixime and 2.5 mg/L tellurite was examined for the growth of additional strains (Table 1) in mEC and NmEC, and it suggested that the combination was useful as a supplement to the enrichment broth.
Although Arai et al. (2021) have reported that E. albertii strains do not grow on agar plates supplemented with 50 μg/L cefixime and 2.5 mg/L tellurite, it has been reported that there is a difference in the sensitivity of agar medium- and liquid medium-supplemented cefixime and tellurite on the growth (Catarame et al., 2003). The chicken meat enrichment culture supplemented with 50 μg/L cefixime and 2.5 mg/L tellurite was found to inhibit the growth of competitive bacteria, but not that of E. albertii (Fig. 4 and Supplementary Fig. S1).
In addition, E. coli strains did not inhibit E. albertii growth in chicken culture in CT-mEC (Supplementary Fig. S2). Therefore, mEC supplementation with 50 μg/L cefixime and 2.5 mg/L tellurite (CT-mEC) was the first selection to enrich food for the detection of E. albertii.
Enrichment in NmEC is appropriate for the detection of E. albertii in chicken meat (Arai et al., 2021). Wakabayashi et al. (2021) reported novobiocin–cefixime–tellurite-supplemented modified TSB for the selective enrichment of E. albertii. In the present study, some E. fergusonii strains from food, however, showed robust growth in CT-NmEC but not in CT-mEC (Table 1). In addition the Ct values for EA-rtPCR of the second culture in CT-mEC using vermicelli Chinese salad-enriched culture were lower than that from the culture in CT-NmEC (Table 2). These results suggested that novobiocin is not essential for the selective growth of E. albertii, and supplementation with 50 μg/L cefixime and 2.5 mg/L tellurite would suffice. We isolated E. albertii from vermicelli Chinese salad by using selective enrichment in CT-mEC but not mEC. These results suggest that selective enrichment in CT-mEC is appropriate for the detection of E. albertii in food. Further analyses with pulsed-field gel electrophoresis and whole genome sequence are needed to reveal the relation between the isolates of E. albertii from vermicelli Chinese salad and isolates from patients.
The occurrence of E. albertii foodborne outbreaks, the contamination of food or environments, and their prevalence in livestock or wild animals is still unclear. Therefore, the effective enrichment developed in this study would be useful for investigating these aspects. Furthermore, evaluation of CT-mEC in an interlaboratory study might be needed for establishment of E. albertii detection method in food.
Conclusions
We have developed an effective enrichment for the selective detection of E. albertii using CT-mEC by investigations of the optimal concentration and combination of cefixime and tellurite. Thus, CT-mEC was evaluated to detect E. albertii in food associated with a foodborne outbreak, and E. albertii was successfully isolated from a food by using selective enrichment in CT-mEC but not in other broths. The method would improve the detection of E. albertii in food and advance research on E. albertii infection.
Footnotes
Authors' Contributions
S.H. and Y.H.-K. designed the study. S.H. performed the experiments. Y.N. provided samples of foodborne outbreak. S.A. provided competitive bacteria from foods. S.H., S.A., and Y.H.-K wrote the article. Y.H.-K. provided funding. All authors approved the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Ministry of Health, Labor, and Welfare, Japan (Health Labor Sciences research grant no. H30-syokuhinippan-001).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.