
Abstract
Several cases of gastrointestinal symptoms including diarrhea and vomiting due to the consumption of Sarcocystis-infected venison have been reported in Japan. However, the control of case incidence is difficult, as epidemiological information concerning Sarcocystis in venison in Japan is insufficient. We examined the prevalence and parasite load of Sarcocystis in 89 samples of Yezo-deer (Cervus nippon yesoensis) venison in Hokkaido by quantifying the copy numbers of the 18S rRNA gene of Sarcocystis, followed by a statistical analysis that considered the sampling area, age, and sex to clarify the parameters related to the parasite load. The copy numbers per gram of venison in samples ranged from 4.8 to 8.8 log. Wilcoxon rank-sum test, the one-way factorial analysis of variance (ANOVA), Steel–Dwass test, and a two-way factorial ANOVA showed significant differences in the copy numbers among sampling areas, not by age or sex, suggesting that the load of Sarcocystis in wild deer depended on the sampling area in Hokkaido. Notably, more than 80% of Hokkaido venison has a higher gene copy number than the meat that caused Sarcocystis fayeri-food poisoning. This information is expected to contribute to the establishment of hygiene standards for safe venison consumption and the control of gastrointestinal symptom cases due to consumption of Sarcocystis-infected venison.
Introduction
In recent years, game meat has become popular fare in Japan. The game meat industry has been promoted to utilize the meat obtained from wild animals hunted for population control. The feeding damage caused by wildlife is estimated to equate to ∼20 billion yen (Ministry of Agriculture, Forestry and Fisheries, 2018), a considerable cost for Japanese agriculture and forestry. Therefore, local governments are taking measures to control the deer population by promoting game meat industries for the effective usage of wild animal carcasses as a resource (Ministry of Agriculture, Forestry and Fisheries, 2018). The deer species native to Japan is the sika deer (Cervus spp., fam. Cervidae).
Nowadays, many local governments throughout Japan have started to promote the venison industry, with the Hokkaido venison being the most widely distributed. This venison is obtained from Cervus nippon yesoensis, known as the Yezo-deer (Kaji et al, 2010). The first case of severe diarrhea caused by eating raw venison was reported in 2012, where Sarcocystis was detected in venison sashimi (Aoki et al, 2017).
Protozoa of the genus Sarcocystis infect two types of hosts during its life cycle, with herbivores serving as intermediate and carnivores as definitive hosts; sexual replication of the parasite occurs, resulting in the shedding of oocysts and/or sporocysts into the environment. Intermediate hosts become infected by ingesting grass or water contaminated with sporocysts; when sporocysts reach the small intestine, sporozoites excyst and undergo schizogony in the endothelium of capillaries. The merozoite forms at the periphery in the endothelial tissues and is then released from the final stage of schizogony. After repeated divisions, a sarcocyst filled with bradyzoites forms in the muscular tissues of the herbivorous host. The whole life cycle of Sarcocystis is completed by the intake of sarcocysts in the muscle of the intermediate host by definitive hosts (Dubey et al, 2015).
In 2011, a case of food poisoning caused by Sarcocystis-infected raw horse meat (basashi) was reported in Kyushu, Japan (Furukawa et al, 2016; Harada et al, 2013). The appearance of vomiting and diarrhea seems to depend on the intake of Sarcocystis (Harada et al, 2013). The causative horse meat contained Sarcocystis fayeri, of >1.2 × 106 copies of 18S rRNA gene per gram (Furukawa et al, 2016).
Regarding Sarcocystis toxicity, Irikura et al (2017) reported the diarrhea-inducing effect of actin-depolymerizing factor (ADF) from S. fayeri. As for other Sarcocystis species, an ADF-like protein that binds to anti-ADF antibodies was detected in S. truncata, a parasite of venison that caused symptomatic cases (Ota et al, 2019). However, the toxicity of Sarcocystis has not been confirmed except for in S. fayeri.
Epidemiological information on Sarcocystis spp. in wild animals remains insufficient; therefore, it is necessary to analyze epidemiological information on Sarcocystis in venison to control the occurrence of gastrointestinal symptomatic cases.
Since the pathogenicity of Sarcocystis spp. in deer has not yet been clarified, we assumed that the parasitic Sarcocystis spp. in deer in this study was pathogenic and therefore attempted to make a presumptive analysis.
In this study, we examined the parasite load of Sarcocystis spp. in venison from Hokkaido and analyzed the relationship between the parasite load and epidemiological factors, such as the sex, age, and region of origin of the deer, to contribute to the safe distribution of venison.
Materials and Methods
Venison samples and DNA extraction
Diaphragm samples were collected from 89 Yezo-deer (C. nippon yesoensis) that were hunted between March 2013 and July 2014 in 3 areas of Hokkaido (northern area n = 15; eastern area, n = 15; and southern area, n = 59). The sampling areas were located ≥200 km from each other (Fig. 1). Detailed information about the samples is shown in Supplementary Data S1. All samples were collected within 2 h after hunting and were delivered to cold storage (4°C) at Iwate University (Morioka, Japan) within 48 h. DNA extraction was performed according to the previously reported protocol (Yamazaki et al, 2021). Ten-gram diaphragm samples were homogenized in 30 mL of phosphate-buffered saline for 1 min at 5000 rpm, using an Excel Auto Homogenizer (NIHONSEIKI KAISHA LTD., Tokyo, Japan). Genomic DNA was extracted from 200 μL of the homogenate using a Qiagen DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol.

Sampling areas in this study. The sampling areas of this study were located in the north, east, and south of Hokkaido Island, Japan. The distance between each of the areas was ∼300 km.
Conventional polymerase chain reaction for the 18S rRNA gene of Sarcocystis
An 1800-bp DNA fragment of 18S ribosomal RNA of genus Sarcocystis was amplified by conventional polymerase chain reaction (PCR), using an EX taq Hot Start kit (Takara Bio, Kusatsu Shiga) with the following set of Sarcocystis-specific primers: forward, 5′-AGCCATGCATGTCTAAGTATAAG-3′; reverse, 5′-TTCCTCTAAGTGTTAAGGTTCAC-3′ (Yamazaki et al, 2021). DNA template (1 μL) was added to 20 μL of reaction mixture containing 2 μL of 10 × Ex Taq Buffer, 4 μL of each dNTP (2.5 mM), 2 μL of each primer (10 mM), 0.2 μL of 5 U/μL Ex Taq HS, and 8.8 μL of water. The following cycling parameters were used: initial denaturation at 94°C for 10 min; 40 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 90 s; and a final extension at 72°C for 5 min.
Distilled water was used as the negative control and S. fayeri genomic DNA as the positive control. The PCR products were visualized under ultraviolet light after staining with GRG-1000 (BIO CRAFT, Tokyo, Japan) following agarose gel electrophoresis.
Sarcocystis species-specific PCR: All 18S rRNA Sarcocystis-positive samples were subject to species-specific PCR for the mitochondrial cytochrome c oxidase subunit I gene (cox1) of Sarcocystis (Abe et al, 2019). PCR amplifications of cox 1 were performed in a final volume of 25 μL containing 1.25 U of TaKaRa Ex Taq HS, 1 × Ex Taq buffer, 2 mM MgCl2, 250 μM of each dNTP, 0.2 μM of each primer, and 2.5 μL of the DNA sample. PCR was conducted under the following conditions: the mixtures were denatured at 94°C for 5 min and then subjected to 40 cycles of 94°C for 30 s, 65°C or 64°C for types 2, 6, and 7 of Sarcocystis species for 30 s, and 72°C for 1 min, followed by a final extension at 72°C for 7 min.
Quantitative real-time PCR for the partial 18S rRNA gene of Sarcocystis
Quantitative real-time PCR was conducted, using an SYBR qPCR kit (GeneAce SYBR qPCR Mix α; NIPPON GENE, Tokyo), with the following Sarcocystis-specific primers: forward, 5′-CGACTTCTCCTGCACCTTATGA-3′; reverse, 5′-TTCAGCCTTGCGACCATACTC-3′, following a previous report (Yamazaki et al, 2021). The PCR conditions were as follows: denaturation at 95°C for 10 min, 45 cycles of 95°C for 30 s, and 60°C for 60 s. A plasmid, pMD20, with an inserted 18S rRNA gene fragment obtained by conventional PCR was used as a positive control. A calibration curve was constructed using the Ct value and copy number of positive controls.
Statistical analyses
Differences in the copy numbers of Sarcocystis 18S rRNA gene from diaphragm samples were statistically analyzed. The explanatory variables were the age, region of origin, and sex of the deer. The age was estimated by the hunters based on tooth replacement. The normality of the copy numbers in all samples was analyzed by the Shapiro–Wilk test, and then, the variance of the copy numbers of an explanatory variable group was analyzed by an F test and Bartlett's test. The statistical methods were chosen according to the equality of variance, sample size, and variability in each group (Table 1).
The Statistical Analysis of Epidemiological Factors Associated with the Sarcocystis Parasite Load in Venison
ANOVA, analysis of variance; ns, not significant.
Comparative analyses with the S. fayeri-food poisoning threshold
It has been reported that food poisoning caused by S. fayeri occurs at an intake of ≥6.1 log (1.2 × 106) copies of Sarcocystis 18S rRNA gene per gram of horse meat (Furukawa et al, 2016). Since the amount of Sarcocystis ingested in symptomatic cases of venison consumption is not known, the ingested amount in cases of food poisoning with horse meat was used as the food poisoning threshold in this study. The prevalence of deer with a parasite load above the S. fayeri-food poisoning threshold of each area was analyzed using Fisher's exact test (extended) and Steel–Dwass test (Supplementary Data S2). It should be noted that the pathogenicity of Sarcocystis spp. in Yezo-deer is still unknown, and this risk analysis is therefore a presumptive one, assuming that all Sarcocystis spp. in Yezo-deer are pathogenic to humans.
Results
Detection and quantification of the Sarcocystis 18S rRNA gene and cox 1 gene in venison by PCR
The Sarcocystis spp. 18S rRNA gene was detected in the diaphragm samples of all 89 Yezo-deer. The copy numbers ranged from 4.7 to 8.8 log. The median gene copy numbers in the three areas were as follows: north, 6.9 log; east, 7.4 log; and south, 6.6 log (Fig. 2). The tendency increased with age. The median gene copy numbers in each age group were as follows: <1 year, 6.3 log; 1 year, 6.3 log; 2 years, 7.0 log; 3 years, 6.8 log; 4 years, 6.9 log; 5 years, 7.4 log; 6 years, 7.2 log; and 8 years, 7.3 log (Fig. 3A). The copy numbers of the male group ranged from 4.8 log to 8.8 log, while the copy numbers of the female group ranged from 4.9 log to 8.6 log. The median gene copy number was 6.8 log in males and 6.9 log in females (Fig. 3B).

Box-plot of the copy number of the 18S rRNA gene of Sarcocystis in venison (area). The copy number of Sarcocystis 18S rRNA gene per 1 g of venison was quantified by real-time PCR. PCR, polymerase chain reaction. An asterisk (*) indicates a p-value of less than 0.01.
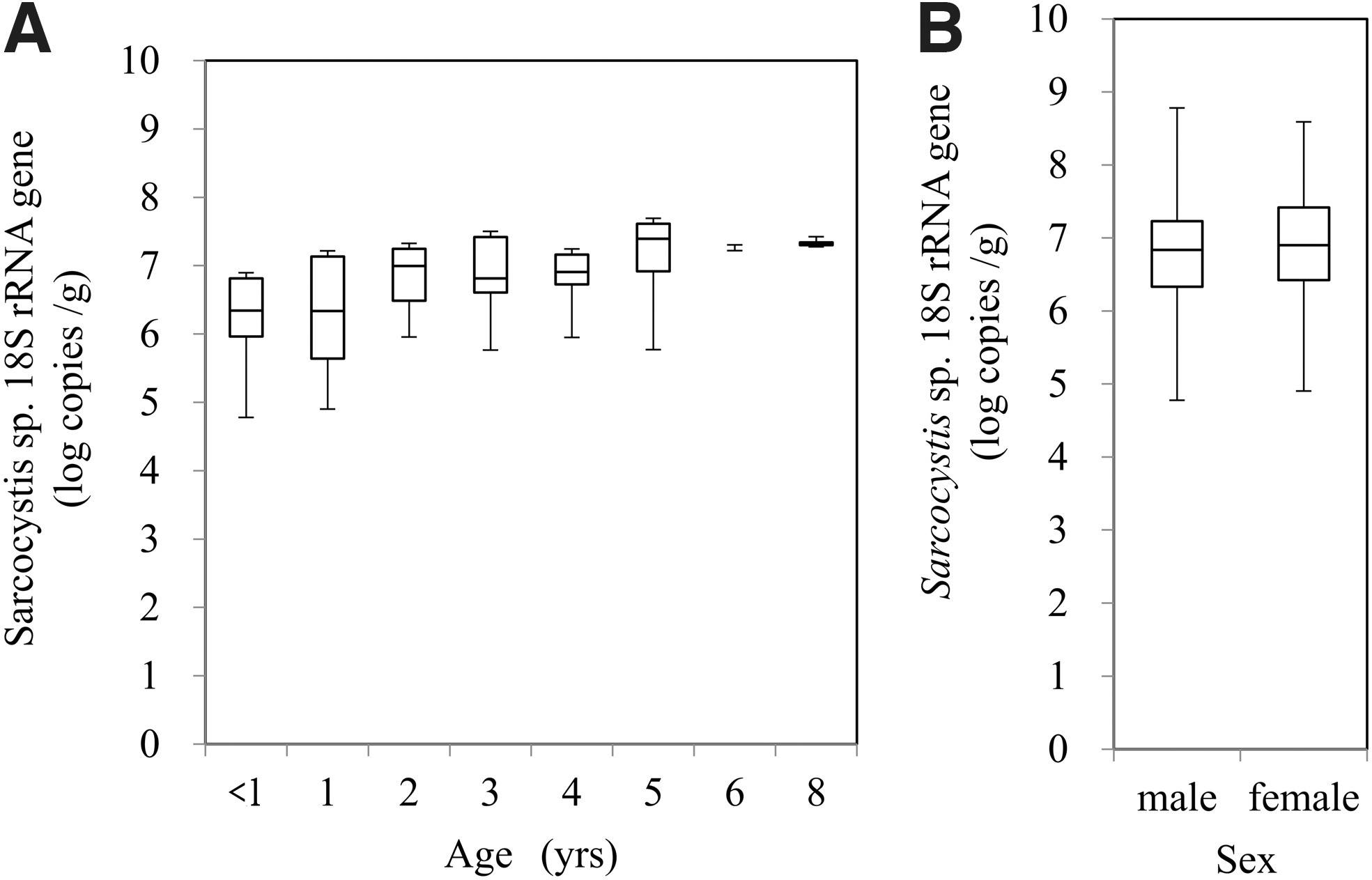
Box-plots of the copy number of the 18S rRNA gene of Sarcocystis in venison according to age
Six samples were identified as S. gjerdei n. sp., cf. S. tarandi, S. japonica n. sp., and S. pilosa by species-specific PCR targeting the cox 1 gene (Supplementary Data S5).
Influence of the area, sex, and age on the Sarcocystis spp. parasite load in venison
A one-way factorial analysis of variance (ANOVA) revealed a significant difference in the Sarcocystis 18S rRNA gene copy numbers among the three areas, with the copy numbers being particularly high in the east area. The p-values between the areas determined by the Steel–Dwass test were as follows: east and south; p < 0.05, east and north; p < 0.05, and north and south; p > 0.05 (Table 1). The statistical comparison of the age groups did not indicate a significant difference (Table 1). Wilcoxon rank-sum test revealed no significant difference in copy number between the sex groups (Table 1).
In this study, most samples from 0- to 2-year-old deer were collected in the south area, while most samples of 3- to 8-year-old deer were collected in the east and north areas (Supplementary Data S3). A two-way ANOVA was performed to verify the cause–effect relationship between the age and area groups. The age–area relationship did not show a significant difference in copy number variance; however, younger deer were concentrated in the south area (Table 1).
The ANOVA that included the age and area (Table 1) showed that the area factor was strongly related to the parasite load of Sarcocystis in Yezo-deer in Hokkaido.
Comparative analysis of the gene copy number with the S. fayeri-food poisoning threshold
A total of 73 (82%) samples contained ≥6.1 log (1.2 × 106) copies/g (Fig. 4). In each sampling area, the number of samples with a higher gene copy number than that in the meat that caused S. fayeri-food poisoning was 13 of 15 (87%) in the north, 15 of 15 (100%) in the east, and 45 of 59 (76%) in the south (Supplementary Data S4). Fisher's exact test (extended) did not show any significant difference in the prevalence of deer with a parasite load ≥6.1 log (1.2 × 106) copies/g between areas. Mann–Whitney's U test showed significant differences in gene copy number between the lower and higher than 6.1 log (1.2 × 106) copies/g groups in each area: north, p < 0.02 and south, p < 0.01 (Supplementary Data S2).
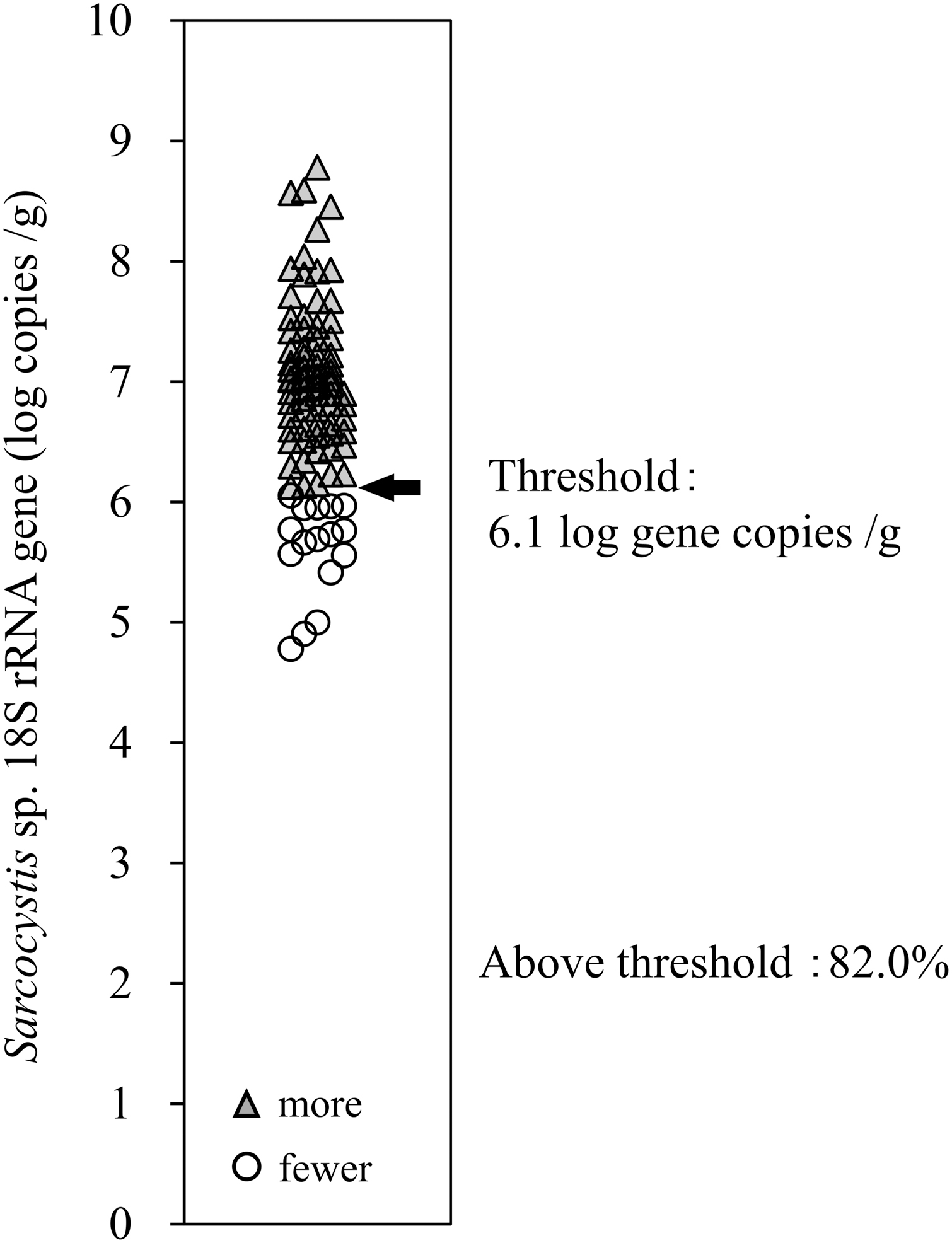
Ratio of venison associated with higher gene copy number than the meat causing Sarcocystis fayeri-food poisoning. The copy number of Sarcocystis 18S rRNA gene per 1 g of venison was quantified by real-time PCR. Black arrow shows the threshold line of 6.1 log (1.2 × 106) gene copies of Sarcocystis 18S rRNA gene per 1 g of venison. White circles show the samples with fewer gene copies than the threshold, while the gray triangles correspond to samples with more genes than the threshold. In total, 82% (73 samples) of the venison samples copy numbers above the threshold.
Discussion
Genus Sarcocystis is a historically well-known protozoa that causes eosinophilic myositis, which results in severe economic damage in the cattle industry (Vangeel et al, 2013). Sarcocystis also causes gastrointestinal symptoms in humans after the consumption of infected meat as reported in Europe, South America, and North America (Dubey et al, 2015; Pal, 2017). In 2011, S. fayeri-infected horse meat caused food poisoning in Japan (Harada et al, 2013). Another outbreak occurred in 2012 with the consumption of Sarcocystis-infected venison (Aoki et al, 2017; Ota et al, 2019). The causative food in both cases was raw or uncooked meat (sashimi). Considering that the development of Sarcocystis disease is intake-dependent (Furukawa et al, 2016), it is noteworthy that a quantitative statistical analysis of Sarcocystis spp. in venison showed that the parasite load was dependent on regional factors.
We hypothesize that this is because of differences in the number of Sarcocystis species distributed among individual areas. For example, S. truncata-like (Type 1), S. tarandi-like (Type 2), and S. pilosa (Type 3) have been detected in Yezo-deer (Irie et al, 2019). Isolation of sporocysts of S. ovalis from jungle crows was reported in 2017 (Irie et al, 2017). Various mammalian and bird species are recognized as definitive hosts of genus Sarcocystis (Dubey et al, 2015). These findings indicate that more epidemiological studies are necessary to clarify the reasons for differences in the parasite load of Sarcocystis in Yezo-deer. In addition, a possible limitation associated with this study must be noted, namely, that the sensitivity of the species-specific PCR method was possibly hampered by the presence of large amounts of deer genomic DNA in the samples. Therefore, a novel Sarcocystis species-specific quantified PCR method is currently being designed for future epidemiological studies.
As mentioned above, 6.1 log (1.2 × 106) copies/g of the gene is the estimated threshold value for S. fayeri-food poisoning (Furukawa et al, 2016). In this study, a total of 73 samples (82%) were over the threshold value (Fig. 4). If all Sarcocystis spp. parasites in Yezo-deer are as pathogenic as S. fayeri, these observations suggest that a high percentage of gastrointestinal symptoms may occur when this venison is served and consumed when it is undercooked. Even in the area with the lowest copy number, the prevalence of deer with a parasite load above the estimated threshold for S. fayeri-food poisoning is 76% (south), which indicates that Yezo-deer meat generally has a high Sarcocystis spp. parasite load. It was clarified that diarrhea in individuals with food poisoning from S. fayeri-infected horse meat was induced by ADF (Irikura et al, 2017; Kamata et al, 2014).
Anti-ADF of S. fayeri antibody-stained bradyzoites in the venison consumed by the patients suggested the presence of an ADF-like substance in Sarcocystis-infected deer (Ota et al, 2019) and possibly revealed an underlying pathogenetic mechanism. This implies that raw venison (shikasashi) harbors a similar risk of food poisoning to horse meat (basashi). However, the analysis in this study is a presumptive one because the pathogenicity of Sarcocystis in sika deer remains unknown. Therefore, the role of the ADF-like substance as a toxic factor in deer meat infected with Sarcocystis still needs to be elucidated.
Game meat industries have been promoted by the Japanese government, but producers and consumers alike need to be aware of the presumptive risk of gastrointestinal symptom from Sarcocystis and other pathogenic microorganisms if venison is consumed undercooked. There is still no precise information concerning the species of Sarcocystis and their parasite load in Yezo-deer from other regions of Japan, and in other subspecies of sika deer. To clarify these points, further scientific investigation of wild deer throughout Japan is necessary.
Conclusion
In this study, we considered the onset of gastrointestinal symptom due to consumption of venison to be intake-dependent and evaluated the presumptive risk associated with consuming venison using the Sarcocystis 18S rRNA gene copy number as an indicator. The results showed that the Sarcocystis parasite load in Hokkaido venison was related to regional factors.
Footnotes
Authors' Contributions
A.Y.: Original draft (lead); formal analysis (lead); and writing—review and editing (equal). Y.S.: Investigation (lead) and writing—review and editing (equal). Y.K.: Conceptualization (lead); supervision (lead); and writing—review and editing (lead).
Compliance with Ethical Standards
The samples used in this study were collected from Yezo-deer that were legally hunted for human consumption. None of the samples, materials, or methods of our study involved human participants.
Disclosure Statement
No competing financial interests exist.
Funding Information
Our work was supported by a Health Labour Sciences Research Grant for Young Scientists (JPMHKA H30-SHOKUHIN WAKATE 003) and a KAKENHI Grant-in-Aid for Young Scientists (19K15974), Japan.
Supplementary Material
Supplementary Data S1
Supplementary Data S2
Supplementary Data S3
Supplementary Data S4
Supplementary Data S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.