
Abstract
High prevalence of livestock-associated methicillin-susceptible and -resistant Staphylococcus aureus (LA-MSSA and LA-MRSA, respectively) in livestock farms, particularly pig farms, is an increasingly serious threat to food safety and public health. In this study, 173 S. aureus (84 MRSA and 89 MSSA) isolates from healthy pigs, farm environments, and farm workers in Korea were examined to determine the (1) genetic diversity of S. aureus isolates (sequence type [ST], spa, and agr types), (2) staphylococcal cassette chromosome mec (SCCmec) types of MRSA isolates, and (3) multidrug resistance (MDR) phenotypes of MRSA and MSSA isolates. Clonal complex 398 (CC398) genotypes of MRSA and MSSA isolates, particularly CC398-spa type t571-agr I lineages, displaying MDR phenotypes were highly prevalent in pig farms. High prevalence of CC398-t571 MRSA and MSSA was more frequently associated with weaning piglets and growing pigs. Moreover, the same clonal lineages of S. aureus isolates colonized both pigs and farm workers, suggesting the transmission of antimicrobial-resistant CC398 MRSA and MSSA between pigs and humans in the pig farms. Furthermore, two dominant SCCmec types, SCCmec V and SCCmec IX, were identified in CC398 MRSA isolates colonizing healthy pigs. To the best of our knowledge, this is the first report of a CC398 LA-MRSA isolate carrying SCCmec IX in Korea. Collectively, these results suggest widespread distribution of the CC398 lineage among MRSA and MSSA isolates in pigs, farm environments, and farm workers in Korea.
Introduction
Colonization or infection with livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) has been increasingly reported in several livestock animals and human workers in close contact with these animals (Broens et al., 2012; Fitzgerald, 2012; Moon et al., 2019; Voss et al., 2005; Wulf and Voss, 2008). Although distinct clonal lineages of LA-MRSA have been found depending on the geographical areas and animal species, sequence type (ST) 398 LA-MRSA is prevalent globally and infects various species of animals, especially pigs (Armand-Lefevre et al., 2005; Back et al., 2020; Gilbert et al., 2012; Fitzgerald, 2012; Tenhagen et al., 2009).
In addition to methicillin resistance, which is mediated by mecA located in the staphylococcal cassette chromosome mec (SCCmec) (Hiramatsu et al., 2013), ST398 LA-MRSA from pig farms frequently displays a multidrug resistance (MDR) phenotype by carrying a broad range of antimicrobial resistance genes (Back et al., 2020; Moon et al., 2015; Oppliger et al., 2012). Since the first report of clonal complex 398 (CC398, ST398, and ST541) (Lim et al., 2012), a high prevalence of CC398 lineages among MRSA isolates from pig farms has been reported in Korea (Back et al., 2020; Moon et al., 2019).
Compared to the well-recognized LA-MRSA (Larsen et al., 2017; Larsen et al., 2015), livestock-associated methicillin-susceptible S. aureus (LA-MSSA) has not yet been studied extensively. Recently, it was suggested that MSSA and non-aureus staphylococci from farm environments play significant roles in propagation and transmission of antimicrobial resistance genes (Archer and Niemeyer, 1994; Eom et al., 2019; Yang et al., 2022). Similar to the highly prevalent ST398 LA-MRSA in pigs, ST398 MSSA isolates displaying MDR phenotypes have been reported in pigs, farm environments, and farm workers (Eom et al., 2019; Wardyn et al., 2015).
However, there is still limited information on the prevalence, clonal diversity, and antimicrobial resistance profiles of MSSA isolates collected from pig farms in Korea. Therefore, in this study, both MRSA and MSSA isolates were obtained from healthy pigs of four different age groups (lactating piglets, weaning piglets, growing pigs, and finishing pigs), farm environments, and farm workers. To analyze the clonal lineages of LA-MRSA and LA-MSSA isolates, the ST by multilocus sequence typing (MLST) scheme, staphylococcal protein A genotype (spa type), accessory gene regulator (agr) type, and SCCmec types were determined. In addition, the antimicrobial resistance profiles of pig farm-associated S. aureus isolates were analyzed.
Materials and Methods
Sample collection
A total of 173 S. aureus isolates were collected from 8 randomly selected pig farms located in 5 provinces in Korea from March to August, 2020. For each pig farm, 40 nasal swabs from 4 different age groups of pigs (lactating piglets [n = 10], weaning piglets [n = 10], growing pigs [n = 10], and finishing pigs [n = 10]), and 12 swabs from farm environments (sewages, fences, drinking systems, floors, and ventilators) were collected. In addition, a total of 27 nasal swabs from pig farm workers (2–4 workers from each pig farm) were obtained with the participants' consent. All swabs from pig farms were kept at 4°C in ice-cooled containers and sent to the laboratory for the culture of MRSA and MSSA within 24–36 h of sample collection. This study was reviewed and approved by IRB/IACUC at Chung-Ang University.
Isolation and identification of S. aureus
S. aureus strains were isolated from the collected swabs, as previously described (Lee et al., 2022). In brief, all nasal and environmental swab samples were used to inoculate 5 mL of fresh tryptic soy broth (Difco Laboratories, Detroit, MI, USA) containing 10% sodium chloride (tryptic soy broth-10% NaCl) and pre-enriched at 37°C for 18–20 h. Next, ∼25 μL of the enriched cultures were streaked onto the Baird-Parker agar (Difco Laboratories) supplemented with potassium tellurite and egg yolk emulsion (Difco Laboratories). After 36–48 h of incubation at 37°C, putative S. aureus colonies displaying clear halos around the black colonies were selected and subcultured for subsequent confirmation. All S. aureus isolates were identified using a matrix-assisted laser desorption ionization-Biotyper real-time bacterial classification system (Bruker Daltonics, Bremen, Germany) and 16S rRNA sequencing, as previously described (Lee et al., 2022).
Antimicrobial susceptibility tests
Antimicrobial susceptibility assays were performed for all S. aureus isolates, according to the Clinical and Laboratory Standards Institute (CLSI) guidelines, CLSI VET01S (CLSI, 2020) and M100 (CLSI, 2022). Sixteen antimicrobial agents were included for the standard disc diffusion assays using Mueller-Hinton agar (Difco Laboratories): ampicillin (AMP, 10 μg), cefoxitin (CEF, 30 μg), penicillin (PEN, 10 μg), chloramphenicol (CHL, 30 μg), ciprofloxacin (CIP, 5 μg), clindamycin (CLI, 2 μg), erythromycin (ERY, 15 μg), fusidic acid (FUS, 10 μg), gentamicin (GEN, 30 μg), mupirocin (MUP, 200 μg), sulfamethoxazole-trimethoprim (SXT, 23.73–1.25 μg), quinupristin-dalfopristin (SYN, 15 μg), rifampicin (RIF, 5 μg), and tetracycline (TET, 30 μg). Except for MUP (Oxoid, Hampshire, United Kingdom), all antimicrobial discs were purchased from BD BBL (Becton Dickinson, Franklin Lakes, NJ, USA).
The minimum inhibitory concentrations of vancomycin (VAN), linezolid (LZD), oxacillin (OXA), teicoplanin (TEC), and tigecycline (TIG) were determined using a standard Etest (AB Biodisk, Dalvagen, Sweden). S. aureus ATCC® 25923 and S. aureus ATCC 29213 were used as reference strains for antimicrobial susceptibility tests.
Molecular typing of MRSA and MSSA isolates
MLST analysis was performed, as previously described (Enright et al., 2000). In brief, the internal fragments of seven housekeeping genes (aroE, arcC, gmk, glpF, pta, tpi, and yqiL) were amplified by polymerase chain reaction (PCR) and sequenced (BIONICS, Seoul, Korea) to assign specific STs based on the S. aureus MLST database (
All S. aureus isolates displaying resistance to CEF were subjected to mecA detection, as previously described (Geha et al., 1994). For mecA-positive MRSA strains, SCCmec typing was performed using a multiplex PCR analysis, as previously described (Kondo et al., 2007). For SCCmec type IX MRSA strains, a specific primer set (5′-GGTCGCACTGATAAGCGAAA-3′ and 5′-GAGCATTAACTTGCCTGTTG-3′, 1590 bp) was designed to amplify the chromosomal cassette recombinase (ccr) gene based on the published sequences of JCSC6690 (NCBI GenBank accession no. AB705452).
The agr types of MRSA and MSSA isolates were determined as described previously (Gilot et al., 2002).
For spa typing, the short sequence repeats of polymorphic X regions in spa were sequenced, as described before (Shopsin et al., 1999). The spa types of staphylococcal isolates were assigned based on the variable number tandem repeats in the SpaServer database (
Statistical analysis
Pearson's chi-square test was used to analyze the differences in antimicrobial resistance between different genotypic or phenotypic groups of S. aureus isolates. One-way analysis of variance (ANOVA) and Scheffe's posthoc test were performed to assess the differences in prevalence of S. aureus isolates among growth phases. IBM SPSS Statistics 23 software (IBM SPSS Statistics 23 for Windows, Chicago, IL, USA) with a statistical significance of p < 0.05 was used for the analysis.
Results
Prevalence of MRSA and MSSA in pig farms
As shown in Table 1, 173 S. aureus (84 MRSA and 89 MSSA) isolates were obtained from pigs (n = 149), farm environments (n = 16), and farm workers (n = 8). In this study, the isolation rates of S. aureus were 46.6% (149/320) in pig nasal swabs, 16.7% (16/96) in environmental swabs, and 29.6% (8/27) in human nasal swabs, respectively. The highest prevalence of S. aureus was observed in weaning and growing pigs, which were colonized in 76.3% and 50% of the pigs in the groups, respectively. The overall prevalence of MRSA among 173 S. aureus isolates was 48.6% (84/173 strains), and 75 of the 84 MRSA isolates (89.3%) were obtained from healthy pigs (Table 1).
Prevalence of Methicillin-Resistant Staphylococcus aureus and Methicillin-Susceptible Staphylococcus aureus in Pigs, Pig Farm Environments, and Farm Workers in Korea
Significant difference (a > b > c, p < 0.05), values with same letter indicate no significant differences.
MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-susceptible S. aureus.
Similar to the S. aureus isolation rates, the highest prevalence of MRSA was observed in weaning (36.3%) and growing pigs (33.8%). In this study, the prevalence of MRSA in pig farm workers was only 7.4% (2/27 swabs).
Genetic characteristics of MRSA and MSSA isolates
MLST analyses of 84 MRSA and 89 MSSA isolates revealed that all 173 isolates belonged to CC398 (ST541 and ST398) clonal lineages (Fig. 1A, B). While 2 dominant STs, ST398 (n = 56) and ST541 (n = 28), were identified in the MRSA isolates, all 89 MSSA isolates belonged to ST398 (Fig. 1B). All the ST398 MRSA isolates had SCCmec V for methicillin resistance, while all ST541 MRSA isolates carried SCCmec IX. As shown in Table 2, agr typing of 173 S. aureus isolates revealed that all MRSA and MSSA isolates were classified into agr type I.

Clonal lineages of methicillin-resistant and -susceptible Staphylococcus aureus strains isolated from healthy pigs, farm environments, and farm workers in Korea. Genetic relatedness of 84 MRSA
Genotypic Diversities of Methicillin-Resistant Staphylococcus aureus and Methicillin-Susceptible Staphylococcus aureus Isolates Obtained from Pig Farms in Korea
MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-susceptible S. aureus; SCCmec, staphylococcal cassette chromosome mec.
In contrast to the single predominant agr I, four different spa types were identified in ST398 MRSA (t571, t18102, t011, and t2876) and ST398 MSSA (t571, t14581, t034, and t1928) isolates (Table 2). Among the seven different spa types, only one type (t571) was detected in both MRSA and MSSA isolates. Unlike the multiple spa types in ST398 isolates, all 27 ST541 MRSA isolates were assigned to t2876.
Antimicrobial resistance profiles of MRSA and MSSA isolates
All 173 S. aureus isolates showed resistance to AMP and PEN (Table 3). As shown in Figure 2A and B, all 84 MRSA and 89 MSSA isolates exhibited multiple drug resistance (MDR) phenotypes to more than three different classes of antibiotics. While the MRSA isolates displayed relatively higher rates of resistance to CEF, FUS, and GEN than the MSSA isolates, resistance to CIP, CLI, ERY, SYN, and TET was significantly higher in the MSSA isolates (Fig. 2A). Correlating with the antimicrobial resistance profiles, although the highest rates of MDR were observed in MRSA, a higher number of MSSA isolates exhibited MDR phenotypes to six to nine different classes of antibiotics than the MRSA isolates (87.6% vs. 47.6%) (Fig. 2B).
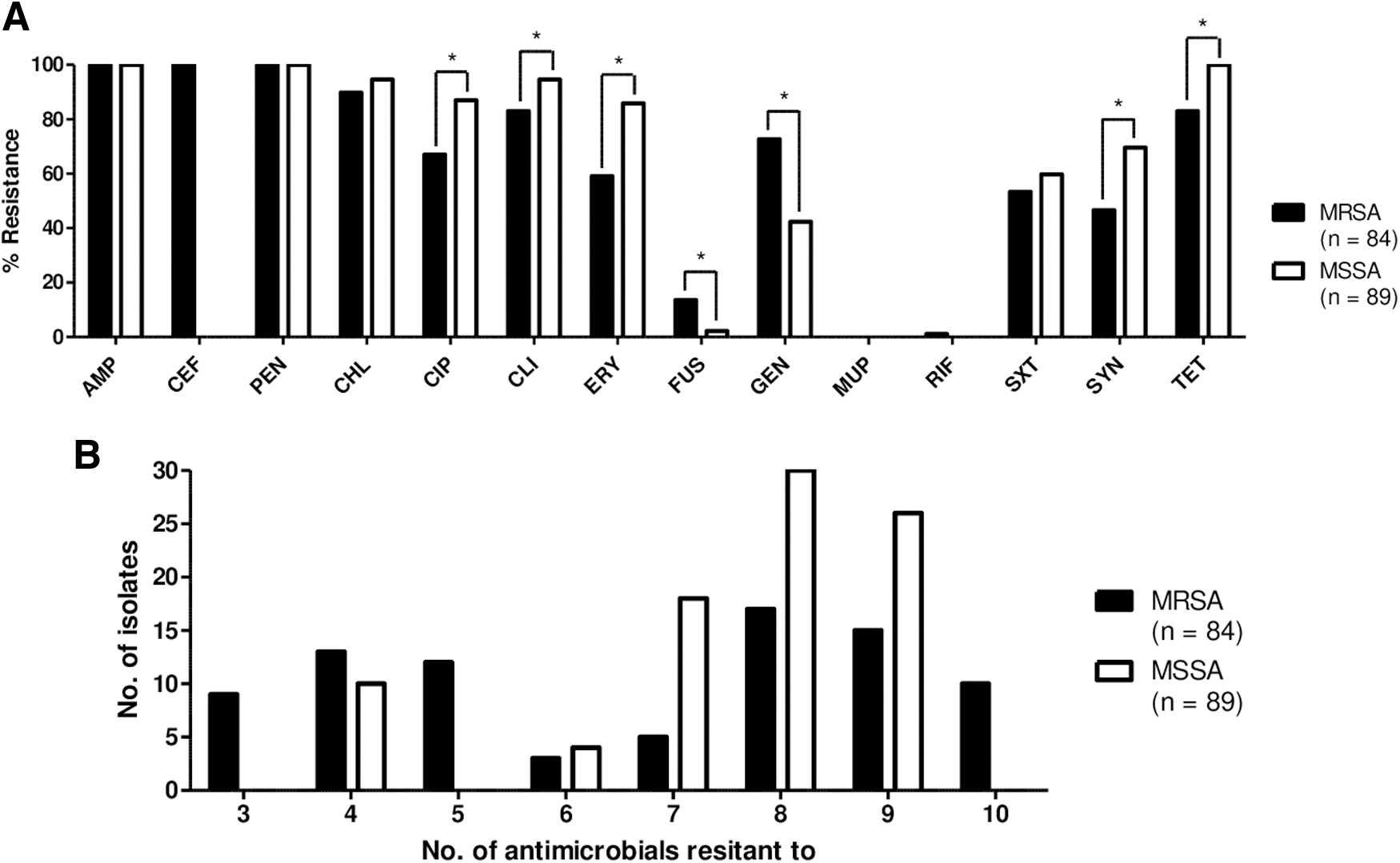
Antimicrobial resistance
Antimicrobial Resistance Profiles of Methicillin-Resistant Staphylococcus aureus and Methicillin-Susceptible Staphylococcus aureus Isolates from Pig Farms in Korea
MDR: Staphylococcus aureus isolates that were resistant to three or more subclasses of antimicrobial drugs are defined as MDR isolates. AMP, ampicillin; CEF, cefoxitin; PEN, penicillin; CHL, chloramphenicol; CIP, ciprofloxacin; CLI, clindamycin; ERY, erythromycin; FUS, fusidic acid; GEN, gentamicin; MUP, mupirocin; RIF, rifampicin; SXT, trimethoprim-sulfamethoxazole; SYN, quinupristin-dalfopristin; TET, tetracycline.
MDR, multidrug resistance; MLST, multilocus sequence typing; MRSA, methicillin-resistant S. aureus; MSSA, methicillin-susceptible S. aureus.
When the two ST groups of MRSA isolates, ST398 and ST541, were compared for antimicrobial resistance, CIP, FUS, and TET resistance rates were higher in the ST398 MRSA isolates, whereas GEN and STX resistance rates were higher in ST541 MRSA isolates (Fig. 3A–C). Interestingly, except for CEF and GEN, ST398 MSSA isolates exhibited similar or higher rates of antimicrobial resistance than the ST398 or ST541 MRSA isolates. All 84 MRSA and 89 MSSA isolates were susceptible to MUP, VAN, LZD, TEC, and TIG.
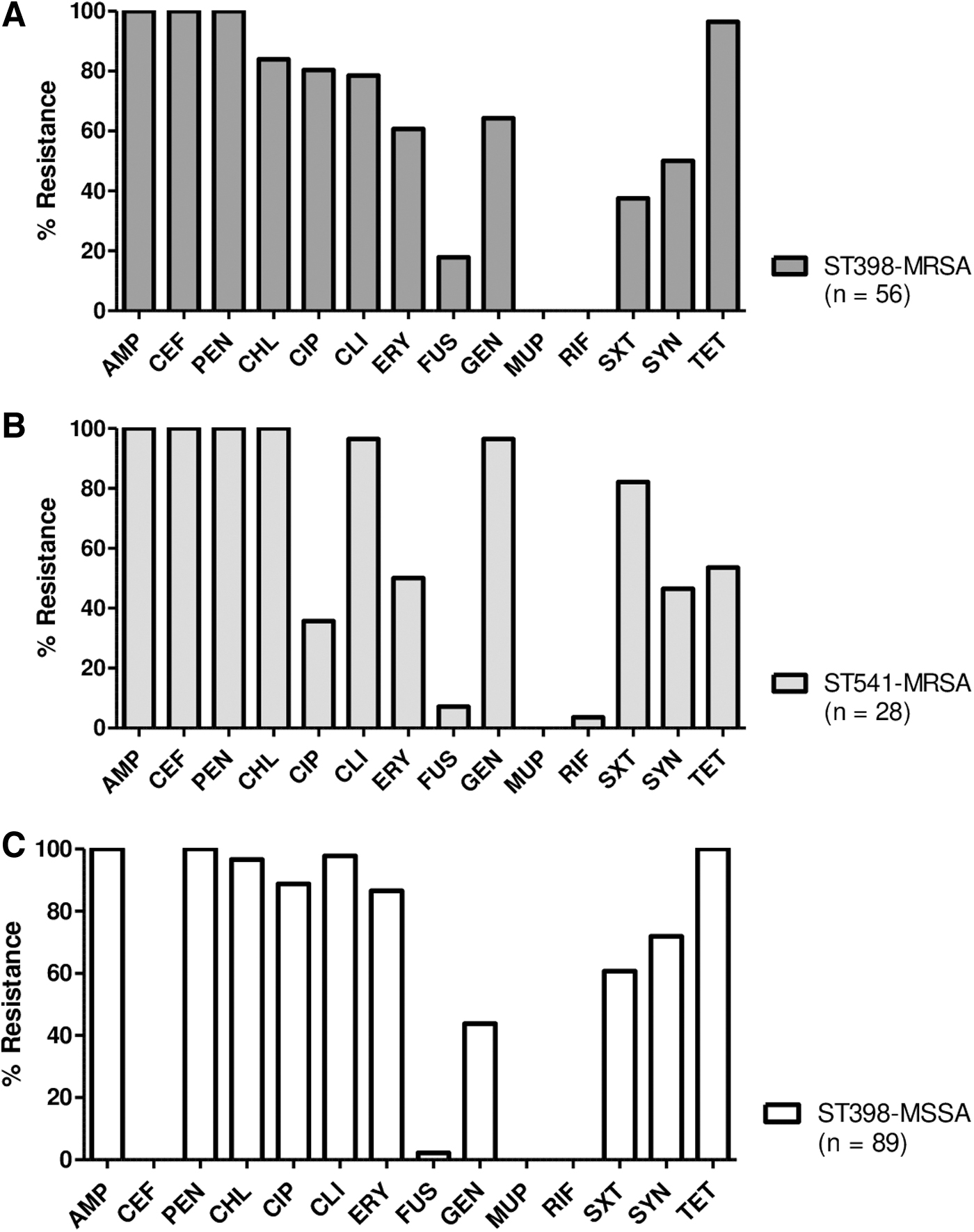
Antimicrobial resistance profiles of ST398 MRSA
Discussion
The current study describes the prevalence and genetic lineages of MRSA and MSSA isolates obtained from pig farms, including healthy pigs, farm environments, and farm workers. The overall isolation rate of S. aureus was 39.1% (173 isolates from 443 swab samples), and 48.5% (84/173 isolates) of S. aureus isolates were MRSA (Table 1). The prevalence of MRSA in pigs (23.4%, 75 MRSA/320 nasal swabs) observed in this study was still lower than that reported in previous studies from North America (49%) (Smith et al., 2009), Germany (49%) (Tenhagen et al., 2009), and the Netherlands (38.7%) (De Neeling et al., 2007), but significantly higher than that (∼3.2–4.4%) reported in two previous studies in Korea (Lim et al., 2012; Moon et al., 2019).
Similar to previous studies in Korea (Moon et al., 2019), which reported a higher prevalence of MRSA in weaning piglets and growing pigs than pigs of other growth stages, weaning piglets and growing pigs displayed higher rates of MRSA colonization than lactating piglets and finisher pigs (Table 1). It has been suggested that higher prevalence of MRSA in weaning piglets and growing pigs might be associated with increased use of antibiotics due to the decrease in maternal immunity in younger pigs (Moon et al., 2019; Pletinckx et al., 2013).
In this study, although the prevalence of S. aureus in pig farm workers was 29.6%, only 2/27 (7.4%) farm workers were colonized by MRSA strains, indicating that the rate of MRSA colonization in pig farm workers was still lower than those of previous reports from Germany (56–85.8%) (Goerge et al., 2017), the Netherlands (23.7–38.2%) (Goerge et al., 2017), and Korea (16.7%) (Moon et al., 2019). Nevertheless, these results indicate that the increased incidence of S. aureus, especially MRSA, in pigs should be monitored as a potential source of transmission to human workers and the food chains in Korea.
In the current study, MLST analysis of 84 MRSA and 89 MSSA isolates identified two predominant genetic lineages, ST398 and ST541, which can be grouped into CC398 (Fig. 1A, B). While two clonal lineages (56 ST398 and 28 ST541) were found in MRSA isolates, all 89 MSSA isolates belonged to ST398 lineage (Fig. 1A, B). Unlike these results, previous studies from Korea reported occurrence of two different types of MRSA, CC398 LA-MRSA carrying SCCmec V and community-associated MRSA (ST72 and ST188 MRSA) carrying SCCmec IV, in pigs and pig farm workers (Moon et al., 2019).
Notably, although CC398 LA-MRSA isolates from pig farms have frequently been associated with SCCmec V in Korea (Back et al., 2020; Lim et al., 2012; Moon et al., 2019; Moon et al., 2015), 2/56 ST398 MRSA isolates and all 28 ST541 MRSA isolates carried SCCmec IX containing the ccrA1B1 complex and class type C2 mec complex (Table 2). Since the first report of a human isolate (Li et al., 2011), several previous studies have identified LA-MRSA isolates harboring SCCmec IX in ST9 and ST398 lineages in pig and pork meat samples in Thailand (Larsen et al., 2012; Vestergaard et al., 2012). MRSA-SCCmec IX isolate was also reported in an outpatient clinic in Thailand (Lulitanond et al., 2013), suggesting the potential transmission of MRSA clones between humans and animals.
To the best of our knowledge, this is the first study to report the occurrence of ST541 LA-MRSA carrying SCCmec IX in pig farms in Korea. The increased frequency of CC398 MRSA and diversification of SCCmec elements necessitates nationwide surveillance for antimicrobial-resistant S. aureus in pig farms.
Unlike previous reports (Lim et al., 2012; Moon et al., 2019), which reported spa types t034 and t571 as the most frequently associated with CC398 LA-MRSA isolates in Korea, t2876 (n = 30), t18102 (n = 24), t571 (n = 22), and t011 (n = 8) types were found in CC398 MRSA isolates in this study (Table 2). Interestingly, all MRSA isolates harboring SCCmec IX were t2876 type, suggesting close genetic relatedness between the ST541-SCCmec IX and ST398-SCCmec IX isolates. In contrast to the four major spa groups in MRSA isolates, the single most dominant clonal lineage of ST398-t571 (n = 67) was found in the MSSA isolates (Table 2). ST398 MSSA-t571 lineage is one of the most common MSSA genotypes in tertiary hospitals in China (Chen et al., 2010).
Moreover, ST398 MSSA-t571 was shown to be associated with skin infections in a livestock farmer in Iowa (Wardyn et al., 2018), indicating that this genotype can be transmitted to human host. Consistent with these data, ST398 MRSA-t571 (n = 1) and ST398 MSSA-t571 (n = 4) isolates were detected in pig farm workers in this study. Therefore, the potential contribution of t571 spa to increased virulence and infectivity of ST398 S. aureus isolates should be investigated in future studies. It has also been hypothesized that ST398-t571 strains are highly receptive to horizontal transfer and acquisition of antimicrobial resistance genes (Bonnet et al., 2018), conferring the strains competitive advantage under antibiotic selection pressure.
Except for the t571 type, all other spa types were detected only in MRSA or MSSA isolates, suggesting that different factors are involved in spa type diversification between CC398 MRSA and MSSA isolates. In contrast to the diversified spa types in MRSA and MSSA isolates, all S. aureus isolates were agr type I.
Most CC398 S. aureus isolates are resistant to antimicrobial agents commonly used in livestock farms, especially TET, macrolides, and β-lactams. As shown in Figure 3A and B, all 84 MRSA and 89 MSSA isolates exhibited MDR phenotype with high levels of resistance to AMP, PEN, CHL, CLI, and TET. ST398 MSSA group displayed significantly higher levels of resistance to CIP, CLI, ERY, SYN, and TET than the CC398 MRSA group (Fig. 2A). When the three major genotypes of S. aureus isolates, ST398-MRSA-SCCmecV, ST541-MRSA-SCCmec IX, and ST398-MSSA, were compared for AMR phenotypes, ST398 isolates showed significantly higher levels of resistance to CIP and TET than ST541 isolates, regardless of methicillin resistance (Fig. 3A–C). Although the ST398 and ST541 isolates are genetically related, their ST-specific resistance profiles to CIP and TET should be monitored in future studies.
Previous studies have suggested that human-adapted ST398 MSSA strains, particularly ST398 MSSA-t571, tend to be TET-sensitive (Price et al., 2012) and exhibit increased adherence to human keratinocytes (Uhlemann et al., 2012). However, all six human isolates in this study, including the four ST398 MSSA-t571 and one ST398-MRSA-SCCmec V-t571, were resistant to TET (Table 3). Moreover, except for one human MSSA strain (ST398-t1928), the same clonal lineages of all other human isolates were detected in pigs (Table 2), indicating that these isolates might have been transmitted between pigs and farm workers.
Our results are based on MRSA and MSSA isolates obtained from a limited number of pig farms during a 6-month sampling period. Thus, future investigations with nationwide sampling of S. aureus over extended periods are necessary. Moreover, additional information on antibiotic consumption in pig farms would help analyze the relationship between antibiotic use and clonal diversity profiles of MRSA and MSSA isolates. Furthermore, persistent colonization of farm workers with CC398 S. aureus should be evaluated over extended period of time.
Conclusions
Our results suggest that (1) CC398 (ST398 and ST541) MRSA and MSSA isolates with MDR phenotypes are highly prevalent in pig farms in Korea; (2) ST398-t571 genetic lineages of MRSA and MSSA are most frequently associated with healthy pigs, especially weaning piglets and growing pigs; (3) CC398 MRSA and MSSA isolates can potentially be transmitted from pigs to pig farm workers; and (4) in addition to the previously reported CC398-MRSA-SCCmec V strains, CC398-MRSA-SCCmec IX strains are also found in pig farms in Korea.
Footnotes
Authors' Contributions
All authors read and approved the final draft of the article and agreed to its publication.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Research of Korea Centers for Disease Control and Prevention (Grant Nos. 2020ER540500 and 2021ER220100).