
Abstract
Shigella sonnei, the causative agents of bacillary dysentery, remains a significant threat to public health. Litsea cubeba essential oil (LC-EO), one of the natural essential oils, exhibited promising biological activities. In this study, the antibacterial effects and possible mechanisms of LC-EO on S. sonnei and its application in lettuce medium were investigated. The minimum inhibitory concentration (MIC) of LC-EO against S. sonnei ATCC 25931 and CMCC 51592 was 4 and 6 μL/mL, respectively. The LC-EO could inhibit the growth of S. sonnei, and decreased S. sonnei to undetectable levels with 4 μL/mL for 1 h in Luria-Bertani broth. The antibacterial mechanism indicated that after the treatment of LC-EO, the production of reactive oxygen species and the activity of superoxide dismutase were significantly elevated in S. sonnei cells, and eventually led to the lipid oxidation product, the malondialdehyde content that significantly increased. Moreover, LC-EO at 2 MIC could destroy 96.51% of bacterial cell membrane integrity, and made S. sonnei cells to appear wrinkled with a rough surface, so that the intracellular adenosine triphosphate leakage was about 0.352–0.030 μmol/L. Finally, the results of application evaluation indicated that the addition of LC-EO at 4 μL/mL in lettuce leaves and 6 μL/mL in lettuce juice could decrease the number of S. sonnei to undetectable levels without remarkable influence on the lettuce leaf sensory quality. In summary, LC-EO exerted strong antibacterial activity and has the potential to control S. sonnei in food industry.
Introduction
S
In recent years, the prevalence of S. sonnei has risen rapidly and gradually become an advantageous strain, and in some areas of China, S. sonnei has become the main endemic flora (Xu et al., 2018a).
Currently, the main methods to control S. sonnei contamination in food or food processing are ionizing radiation, high-temperature sterilization, disinfectants, etc. (Singla et al., 2011), however, these methods have certain limitations. Ionizing radiation is used to ensure food safety and quality, but it remains controversial and public acceptance is low due to concerns about residual radiation (Munir and Federighi, 2020). As an extensive and effective sterilization method, high-temperature sterilization is widely used in the food industry, whereas high temperature will lead to the destruction of vitamins and other nutrients in food and irreversible damage to food senses (Van Impe et al., 2018). Bacteria treated with chemical disinfectants often show resistance, and their residues on the food surface will bring risks to food safety (Josewin et al., 2018).
In conclusion, due to the limitations of these methods, it is of great significance to find an efficient, safe, low-cost sterilization method that does not affect the food quality and sensory properties for the control of S. sonnei in food (Cui et al., 2021).
Litsea cubeba, which belongs to the Lauraceae family, is a kind of plant spice resource unique to China, and its fruit can be used as medicine (Cui et al., 2019; Kamle et al., 2019). L. cubeba essential oil (LC-EO) is a plant essential oil extracted from L. cubeba, which is a nontoxic light yellow oily liquid (Si et al., 2012). Its main active ingredient is citral, which has strong biological activities, including anti-inflammatory and antioxidant activities (Ju et al., 2018). Study has shown that LC-EO has a good inhibitory effect on Escherichia coli, and it changes the cell structure and morphology of E. coli (Nguyen et al., 2018).
However, the antibacterial activity of LC-EO against foodborne pathogenic bacteria has not been reported in detail, and the antibacterial effect and mechanism of LC-EO against S. sonnei have not been investigated. In this study, the inhibitory effect and underlying mechanisms of LC-EO on S. sonnei were evaluated. Besides, the inactivation effects of LC-EO on S. sonnei in contaminated lettuce leaves and lettuce juice were determined, and the sensory evaluation of lettuce treated with LC-EO was carried out.
Materials and Methods
Reagents
Luria-Bertani (LB) agar and broth were purchased from Land Bridge Technology Co., LTD. (Beijing, China). LC-EO (high-performance liquid chromatography ≥95%, CAS 68855-99-2) was purchased from Yuanye Bio-Technology Co., LTD. (Shanghai, China), and LC-EO was dissolved in dimethyl sulfoxide (DMSO) [final concentration of 0.5% (v/v) DMSO in this assay]. All other chemicals and reagents were of analytical grade.
Bacterial strains and culture
S. sonnei (Strain No. 25931) was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). S. sonnei (Strain No. 51592) was purchased from the National Center for Medical Culture Collections (CMCC, China). To prepare bacterial suspension, the strain was streaked on LB agar and incubated at 37°C for activation. After 18 h, the strain was inoculated into sterile LB broth and shaken at 130 rpm for 16 h at 37°C. The bacterial suspension was then centrifuged (4°C, 8000 × g, 5 min), washed twice with phosphate-buffered solution (PBS), and resuspended in LB broth or PBS to achieve the wavelength of 600 nm (OD600nm) of 0.5 (∼3 × 108 CFU/mL).
Antibacterial activity
Determination of minimum inhibitory concentration and minimum bactericidal concentration
The minimum inhibitory concentrations (MICs) of LC-EO against S. sonnei ATCC 25931 and CMCC 51592 were determined using the broth microdilution method according to the Clinical and Laboratory Standards Institute (CLSI, 2021) guideline. Briefly, LC-EO diluted in LB broth was mixed with equivalent volumes of prepared bacterial suspension (5 × 105 CFU/mL) to make the final LC-EO concentrations of 0 (Control), 1, 2, 3, 4, 5, and 6 μL/mL, and then, the mixtures (200 μL) were added to the wells of a 96-well microtiter plate. LB broth containing different concentrations of LC-EO or 0.5% DMSO was used as the negative control. The OD600nm of each well was measured by a microtiter plate reader (Model 680; Bio-Rad, Hercules, CA, USA), and OD600nm measurements were repeated after being incubated at 37°C for 24 h.
The MIC of LC-EO against S. sonnei was defined as the lowest concentration that resulted in ΔOD600nm (final OD600nm value minus initial OD600nm value) of ≤0.05. For minimum bactericidal concentration (MBC) determinations, 100 μL of bacterial suspension from each well was plated onto an LB agar plate. Following incubation for 48 h at 37°C, the MBC was defined as the lowest antimicrobial concentration that did not allow any bacterial growth.
Growth curves
Growth curves were constructed according to the modified method primarily described by Tian et al. (2021). To be specific, the S. sonnei ATCC 25931 suspension (2 × 106 CFU/mL) was mixed with different volumes of LC-EO (dissolved in 0.5% DMSO) to make the final LC-EO concentration 0 (Control), 0.00625 (1/64 MIC), 0.0125 (1/32 MIC), 0.25 (1/16 MIC), 0.5 (1/8 MIC), 1 (1/4 MIC), 2 (1/2 MIC), and 4 (MIC) μL/mL, respectively. The negative control was established by a combination of LB broth and LC-EO without any pathogens. The samples were then incubated in a fully automated Bioscreen C Plate Reader (Labsystems, Helsinki, Finland) at 37°C, and the growth rate was measured at 600 nm at 1-h intervals for 24 h.
Inactivation effect of LC-EO on S. sonnei in LB broth
LC-EO was tested to determine the inactivation effect on S. sonnei as described by Shivaprasad et al. (2021), with little modifications. Briefly, the LC-EO solutions were added to the bacterial suspension (1 × 106 CFU/mL) diluted in LB broth at final LC-EO concentrations of 0 (Control), 2, 3, and 4 μL/mL. Subsequently, after being incubated at 37°C for 0, 0.5, 1, 2, 4, 6, and 8 h, the mixed bacterial suspension was plated onto LB plates, and incubated at 37°C for 24 h before count.
Detection of the intracellular reactive oxygen species level
The reactive oxygen species (ROS) assay kit based on 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA; Beyotime Institute of Biotechnology, Shanghai, China) oxidation was used to determine the ROS level, according to the methods reported by Lin et al. (2019), with minor modifications. Before LC-EO treatment, S. sonnei cells (1 × 107 CFU/mL) were collected, washed with PBS, and incubated at 37°C in dark for 10 min in PBS containing DCFH-DA (final concentration was 5 μM). After LC-EO treatment (0 [Control], 1/2 MIC, MIC, 2 MIC) for 10 min, cells were collected by centrifugation at 12,000 × g for 10 min and washed twice with PBS. ROS fluorescence was measured on the multimode microplate reader platform (Spark®; Tecan, Männedorf, Switzerland), at an excitation and emission wavelength of 488 and 525 nm, respectively.
Measurement of intracellular adenosine triphosphate concentrations
The intracellular adenosine triphosphate (ATP) concentration was measured as described by Zheng et al. (2020), slightly modified. In short, S. sonnei suspension (3 × 108 CFU/mL) was mixed with 0 (Control), 1/2 MIC, MIC, and 2 MIC of LC-EO, followed by incubation at 37°C for 10 min. After a 7-min sonication on ice, all samples were heated at 100°C for 2 min and then centrifuged to collect the supernatant (8000 × g, 5 min, 4°C). Then the ATP concentration was measured using an ATP detection kit (Beyotime Bioengineering Institute, Shanghai, China).
In short, the extracted ATP-containing supernatant and ATP standard solution (100 μL) were added to a white 96-well microtiter plate. Then, an equal volume of ATP detection reagent was added, after which the luminescence signal was measured by a multimode microplate reader. The intracellular ATP concentration of the sample was calculated based on the standard curve generated by the luminescence value of the ATP standard solution.
Detection of the superoxide dismutase activity
The superoxide dismutase (SOD) activity was carried out by an SOD activity detection kit (NBT method; Beyotime Bioengineering Institute, Shanghai, China), according to a previously published protocol (Martínez et al., 2020). The bacterial suspension (3 × 108 CFU/mL) was prepared and treated with LC-EO solution [0 (Control), ½ MIC, MIC, and 2 MIC] at 37°C for 10 min. After being centrifuged, cells were lysed with lysozyme (Solarbio, Beijing, China) at 37°C for 6 min. Then the supernatant was collected by centrifugation (10,000 × g, 3 min, 4°C). NBT/enzyme working solution, the extracted supernatant, and the reaction initiation working solution were sequentially added to the wells, and the 96-well plate was incubated for 30 min at 37°C, and then the absorbance was measured at 560 nm using a microplate reader (Spark®; Tecan, Männedorf, Switzerland).
Malondialdehyde concentration generation in S. sonnei
The content of malondialdehyde (MDA) concentration was detected by the kit (Solarbio, Beijing, China) as previously described by Shi et al. (2021) with modification. The bacteria (3 × 108 CFU/mL) exposed to LC-EO (0 [Control], 1/2 MIC, MIC, and 2 MIC) for 10 min were centrifuged (10,000 × g, 10 min, 4°C) and the supernatant was collected for MDA measurement. Then the supernatant and TBA working solution were mixed and heated at 100°C for 1 h. Then 200 μL of supernatant was added into a 96-well plate after being heated and cooled to room temperature. MDA concentration of each experimental group was calculated by measuring the absorptions at 450, 532, and 600 nm, and the result was calculated according to the formula below: where ΔOD532, ΔOD600, and ΔOD450 refer to the difference between the absorbance of the positive control group and blank control ground at 532, 600, and 450 nm, respectively.
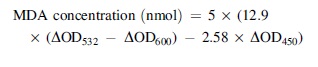
Flow cytometric analysis of bacterial cells
Flow cytometry was used to construct the percentage of live and dead bacteria after cells were stained with the LIVE/DEAD® BacLight™ Bacterial Viability Kit (Molecular Probes, Eugene, OR, USA), following the method reported previously (Inoue et al., 2020). In brief, the bacterial suspension was harvested (8000 × g, 5 min, 4°C), washed twice with 0.85% (m/v) NaCl, and resuspended at 1 × 106 CFU/mL. After 30 min of LC-EO treatment (0 [Control], 1/2 MIC, MIC, and 2 MIC) at 37°C, the mixture was centrifuged for 2 min at 10,000 × g and then the pellet resuspended in 200 μL of 0.85% NaCl.
Suspensions of untreated and 70% (v/v) isopropyl alcohol-treated cells served as the negative control and the positive control, respectively. After 1 mL of the suspension was incubated with 10 μM of SYTO9 and 60 μM of PI at 37°C for 15 min, live and dead bacterial cells were counted on a flow cytometer (CytoFLEX; Beckman Coulter, Inc., USA) with 525 and 620 nm channels for SYTO 9 and PI fluorescence detection, respectively.
Field emission scanning electron microscopy analysis
Field emission scanning electron microscopy (FESEM) is used to observe cell morphology and surface changes. This experiment is based on the design of Cai et al. (2019). The bacterial suspension (3 × 108 CFU/mL) was treated with 0 (Control), 1/2 MIC, MIC, and 2 MIC of LC-EO. Supernatants were discarded after centrifugation, and cell pellets were washed twice with PBS and fixed in 2.5% (v/v) glutaraldehyde overnight at 4°C. The fixed cells were centrifuged (5000 × g, 5 min, 4°C) and washed three times with PBS, and then washed with different concentrations of ethanol sequentially (30%, 50%, 70%, 90%, and 100%, v/v). Finally, the bacterial suspension was dried and fixed on supports. After being sputter-coated with gold inane ion coater, cells were observed using FESEM (S-4800; Hitachi, Tokyo, Japan) at × 15,000 magnification.
Antibacterial effect of LC-EO on lettuce leaves and lettuce juice
Sample preparation and inoculation
Lettuce was purchased in the local supermarket (Yangling, China) and stored at 4°C until use. The lettuce sample preparation was performed as previously described by Xu et al. (2021). Washed twice with ultrapure water, green leaves were cut into 5 × 5 cm2 strips. Before treatment, lettuce strips were soaked in alcohol (75% v/v) for 30 min to minimize the amount of potential initial bacterial contamination. Then, the lettuce leaves were rinsed with ultrapure water to dilute any residual ethanol and preservative that might be present and could inhibit bacterial growth. The lettuce juice was prepared as described by Kang et al. (2022), with 45 g of lettuce being juiced with 200 mL of sterile water. After being irradiated (6 kGy, 2 s) to remove the background bacteria, the lettuce juice was stored at 4°C and used up within 1 week.
The bacterial suspension of S. sonnei ATCC 25931 and CMCC 51592 was diluted to a concentration of 1.2 × 106 CFU/cm2 and 1 × 106 CFU/mL in lettuce leaves and lettuce juice, respectively.
Bactericidal effects of LC-EO on lettuce leaves and lettuce juice
The bactericidal effect of LC-EO against S. sonnei on lettuce was used similarly to that previously described (Ferreira and Domingues, 2016). LC-EO was mixed in aseptic sealed bags and lettuce juice so that the final concentrations of LC-EO in lettuce leaves and lettuce juice were 0 (Control), 2.5, 3.75, and 5 μL/mL and 0 (Control), 5, 6, and 8 μL/mL, respectively. For bactericidal effects of LC-EO on lettuce leaves, at 0, 10, 30, 60, 90, and 120 min, and at 0, 0.5, 1, 2, 4, 6, and 8 h for bactericidal effects of LC-EO on lettuce juice, the samples were taken out from 25°C and shaken in a homogenizer for 2 min, the collection fluid (1 mL) serially diluted in PBS, plated on LB agar, and incubated at 37°C for 24 h, and the numbers of S. sonnei colonies were counted.
Measurement of lettuce leaf browning index and chlorophyll content
The browning index of lettuce after LC-EO treatment was determined using two pieces of each sample (Kang and Song, 2021). The samples (1 g) were mixed with 10 mL of 10% (m/v) trichloroacetic acid, ground evenly, and centrifuged at 8000 × g for 10 min. The absorbance of supernatants, which were measured in triplicate at 420 nm, is the browning index.
The chlorophyll content of treated leaves was determined by the method reported by Han et al. (2021), with minor modifications. The treated lettuce sample (1 g) was ground with 10 mL of methanol and then rinsed repeatedly with acetone to fully collect the grinding fluid. Then the samples were centrifuged for 10 min at 8000 × g. The absorbance was measured at 665 and 652 nm, respectively, and the concentration of chlorophyll was calculated.
Statistical analysis
The whole experiments were repeated three times independently. IBM SPSS statistics software version 26.0 (SPSS, Inc., Chicago, IL, USA) was used to statistically analyze, and the results were expressed as mean ± standard deviation. Analysis of variance (ANOVA) procedure and Tukey's test were used to determine statistically significant (p < 0.05) differences among treatments.
Results
MICs and MBCs
According to the analyses performed, the MIC of LC-EO against S. sonnei ATCC 25931 and CMCC 51592 was 4 μL/mL (3.532 mg/mL) and 6 μL/mL (5.298 mg/mL), respectively. The MBC values of all the tested strains in this study were identical to their MIC values. S. sonnei ATCC 25931 was selected as a model organism for use in all the subsequent experiments.
Growth curves
The effect of LC-EO on the growth curves of S. sonnei ATCC 25931 in LB broth is shown in Figure 1. It can be seen that the OD600nm values of all the LC-EO-treated groups were lower than the control group, indicating that normal bacterial growth was inhibited, especially in the MIC group. By comparison of different growth curves, it was demonstrated that higher concentrations of LC-EO resulted in a lower growth rate compared with the control group.

Growth curves of Shigella sonnei ATCC 25931 cultured in LB broth with various concentrations of LC-EO. LB, Luria-Bertani; LC-EO, Litsea cubeba essential oil.
Inactivation effect of LC-EO on S. sonnei in LB broth
As presented in Figure 2, the initial number of bacteria was similar among the treatment groups (6.0 ± 0.3 log CFU/mL). The counts of S. sonnei in the control group reached 8.6 ± 0.2 log CFU/mL within 0–4 h and remained stable until 8 h. Whereas at the 2 μL/mL concentration of LC-EO, S. sonnei ATCC 25931 remained stable within 0–1 h and reached 8.2 ± 0.1 log CFU/mL at 8 h. After S. sonnei was treated with LC-EO at 3 μL/mL for 4 h, the cell count was 6.1 ± 0.2 log CFU/mL and remained stable until 8 h. After LC-EO treatment at 4 μL/mL for 1 h, the number of bacteria decreased to undetectable levels.

Inactivation of Shigella sonnei by LC-EO in LB broth. **Indicate statistical significance at p < 0.01 compared with the control. LB, Luria-Bertani; LC-EO, Litsea cubeba essential oil.
Detection of the intracellular ROS level
As illustrated in Figure 3, the intracellular ROS levels in the control group were 8.5 ± 0.8. After treatment of S. sonnei ATCC 25931 with LC-EO at 0, 1/2 MIC, MIC, and 2 MIC, the intracellular ROS levels increased significantly (p < 0.01) to 14.9 ± 1.1, 16.2 ± 1.8, and 18.8 ± 2.4, respectively.
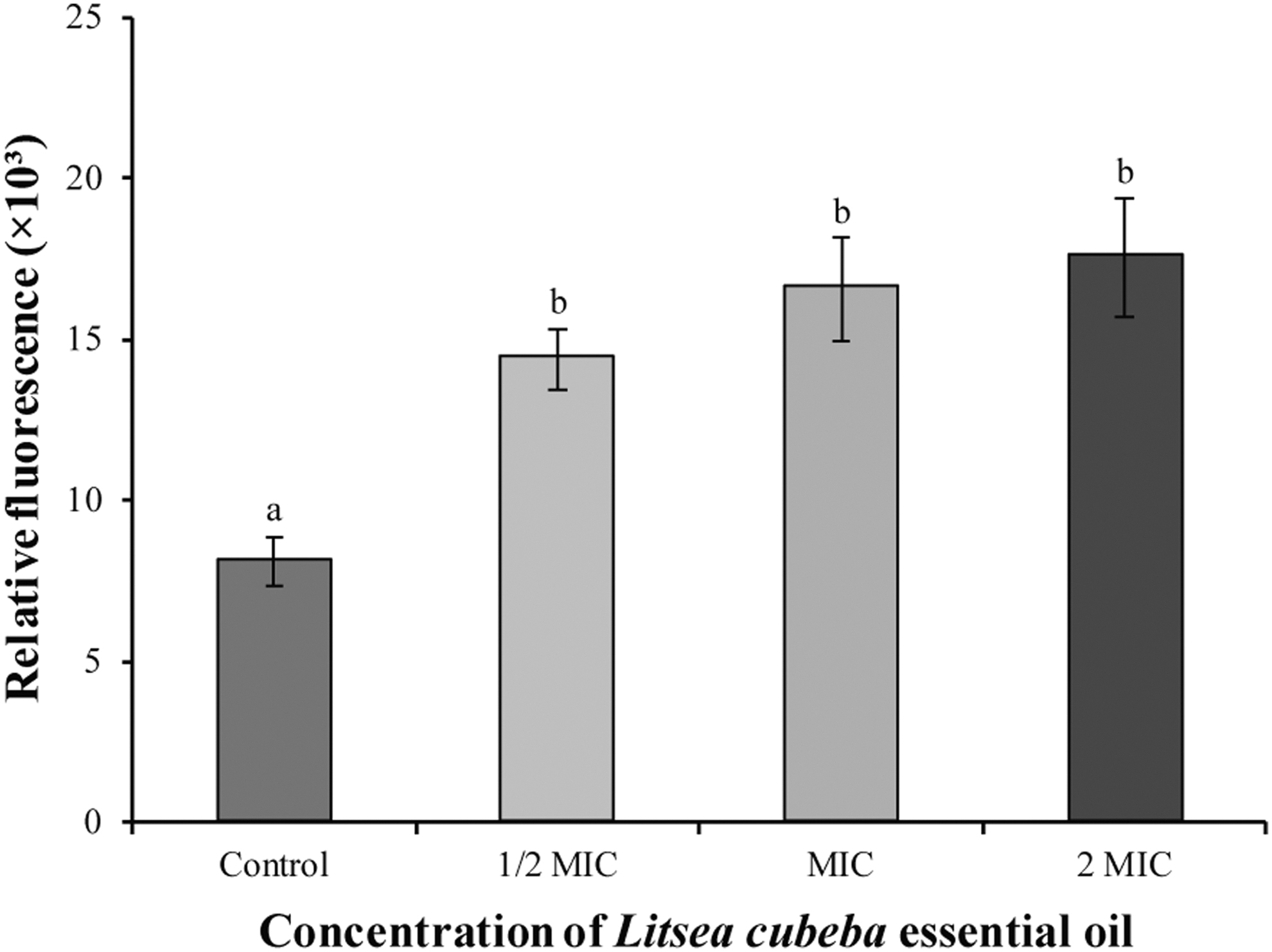
Effect of LC-EO treatment on intracellular ROS levels in Shigella sonnei ATCC 25931. Different lowercase letters indicate statistically significant differences between the means (p < 0.01). LC-EO, Litsea cubeba essential oil; ROS, reactive oxygen species.
Changes in intracellular ATP concentrations
There was a linearity between the relative luminescence units and ATP concentration (y = 321,226x + 2549; R 2 = 0.9991, the standard curve is not shown). As presented in Figure 4, compared with the effects of the control (0.352 ± 0.002 μmol/L), the intracellular ATP concentrations of S. sonnei ATCC 25931 decreased significantly (p < 0.01) following exposure to LC-EO at 1/2 MIC (0.151 ± 0.001 μmol/L), MIC (0.093 ± 0.004 μmol/L), and 2 MIC (0.030 ± 0.001 μmol/L).

Effects of LC-EO on intracellular ATP concentrations in Shigella sonnei ATCC 25931. Different lowercase letters indicate statistically significant differences between the means (p < 0.01). ATP, adenosine triphosphate; LC-EO, Litsea cubeba essential oil.
Effect of citral on SOD activity of S. sonnei
As shown in Figure 5, the SOD activity of S. sonnei without LC-EO treatment was 1.07 ± 0.05 U, and the level of SOD activity of S. sonnei treated with LC-EO significantly (p < 0.01) increased.

Effects of LC-EO on SOD activity in Shigella sonnei ATCC 25931. Different lowercase letters indicate statistically significant differences between the means (p < 0.01). LC-EO, Litsea cubeba essential oil; SOD, superoxide dismutase.
Effect of LC-EO on extracellular MDA content of S. sonnei
As shown in Figure 6, the level of extracellular MDA content of S. sonnei treated with LC-EO increased significantly (p < 0.01). The MDA concentration of S. sonnei without LC-EO treatment was 0.006 ± 0.018 nmol/mL. While the action of LC-EO at 1/2 MIC, MIC, and 2 MIC caused the MDA concentration to be raised significantly to 2.930 ± 0.757, 4.568 ± 0.096, and 6.080 ± 0.180 nmol/mL, respectively.

Effects of LC-EO on extracellular MDA contents in Shigella sonnei ATCC 25931. Different lowercase letters indicate statistically significant differences between the means (p < 0.01). LC-EO, Litsea cubeba essential oil; MDA, malondialdehyde.
Flow cytometric analysis of bacterial cells
As shown in Figure 7, the changes of the cell membrane integrity of S. sonnei after the action of LC-EO were observed and determined by flow cytometry. The integrity of bacterial cell membrane decreased after treatment with LC-EO (Fig. 7B–D), and the proportion of damaged bacteria (R1) gradually increased. After that, the number of damaged cells increased from 21.06% (Fig. 7B) to 96.51% (Fig. 7D), which indicated that the LC-EO could damage the cell membrane integrity of S. sonnei.

Flow cytometry determination of the cell membrane of Shigella sonnei after treatment with 0 (Control)
FESEM analysis
Untreated S. sonnei ATCC 25931 cells are shown in Figure 8A, and there were significant morphological differences between treated cells (Fig. 8B–D) and the control (Fig. 8A). Untreated S. sonnei cells were rod-shaped with a smooth surface (Fig. 8A), while S. sonnei cells treated with LC-EO at the 1/2 MIC appeared wrinkled with a rough surface (Fig. 8B), while the outer membrane of cells exposed to LC-EO at MIC possessed a more wrinkled surface and notable depressions (Fig. 8C). When cultures were treated with LC-EO at 2 MIC, normal cell morphology was absent (Fig. 8D).

FESEM observations ( × 15,000 magnification) of Shigella sonnei ATCC 25931. Untreated for 2 h
Bactericidal effects of LC-EO on lettuce leaves and lettuce juice
As shown in Figure 9A, the initial number of viable S. sonnei on lettuce leaves was 5.6 ± 0.1 log CFU/cm2. The bacterial count of the control group was retained at about 6 log CFU/cm2 for 120 min, while the bacterial count of 2.5 and 3.75 μL/mL groups decreased to 3.6 ± 0.1 and 3.2 ± 0.3 log CFU/cm2, respectively. The number of bacteria decreased to undetectable levels in 5 μL/mL groups in 60 min on lettuce leaves.
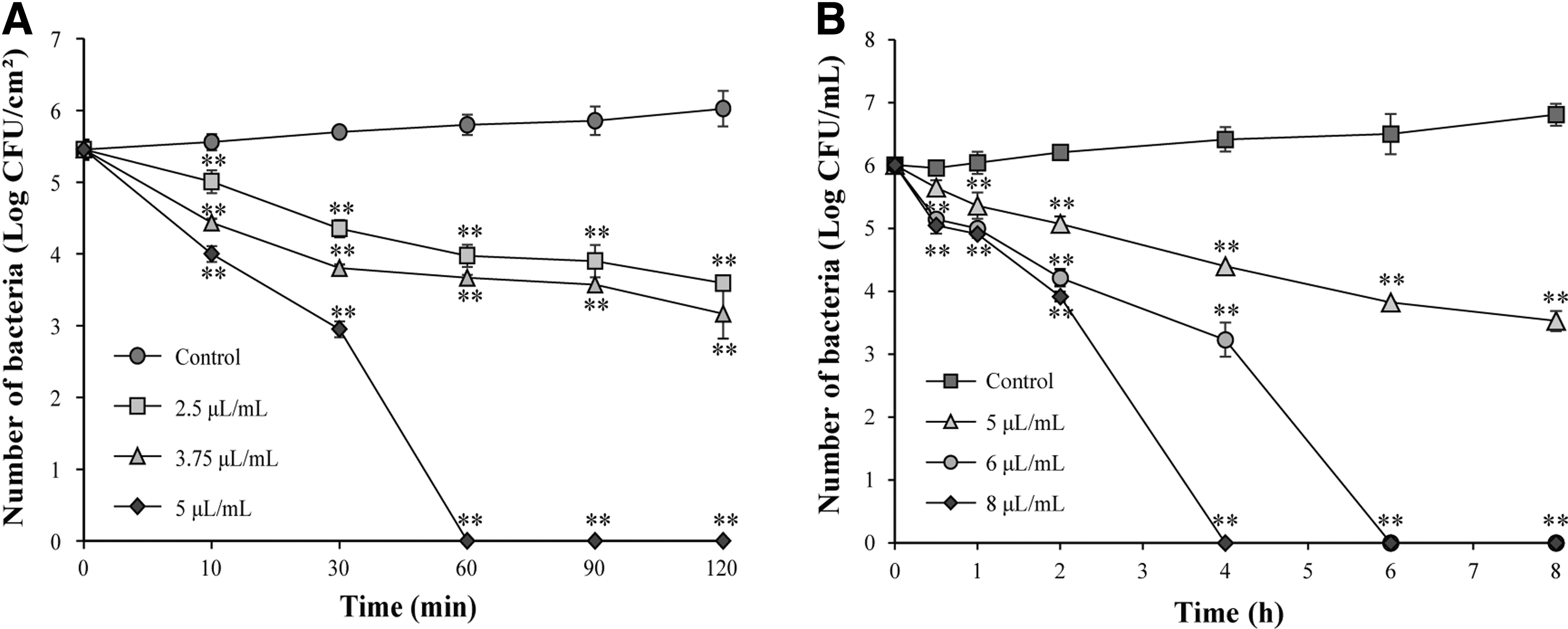
Inactivation of Shigella sonnei on lettuce leaves
For bactericidal effects of LC-EO in lettuce juice (Fig. 9B), the bacterial count of the control group increased from 6.1 ± 0.1 to 6.8 ± 0.1 log CFU/mL within 8 h, while the bacterial count of 5 μL/mL groups decreased to 3.5 ± 0.2 log CFU/mL, and the number of bacteria decreased to undetectable levels in 6 μL/mL groups and 8 μL/mL groups in 4 and 6 h, respectively.
Measurement of lettuce leaf browning index and chlorophyll content
The changes in the browning index and chlorophyll content in postcut LC-EO-treated or nontreated lettuce during storage at 4°C are shown in Table 1. The chlorophyll content was significantly low in control group lettuce. The chlorophyll content increased with increasing the LC-EO treatment concentration. The browning index was significantly low in 1/2 MIC-treated lettuce, while control group increased more rapidly than that in the other treatments. In addition, the browning index of LC-EO-treated lettuce showed lower than that of control group lettuce. These results indicated that LC-EO has a retarding effect against the browning of fresh-cut lettuce.
Changes in the Quality of Lettuce Leaves After Treatment
Any means in the same column followed by different letters (A–D) are significantly different (p < 0.05) by Duncan's multiple range test.
Any means in the same row followed by different letters (a–d) are significantly different (p < 0.05) by Duncan's multiple range test.
Control: Nonwashed samples, 1/2 MIC: 2 μL/mL LC-EO, 3/4 MIC: 3 μL/mL LC-EO, MIC: 4 μL/mL LC-EO.
MIC, minimum inhibitory concentration.
Discussion
In the present study, the LC-EO showed the antibacterial activity against S. sonnei ATCC 25931 and CMCC 51592, and the measured MIC values were 4 (3.532 mg/mL) and 6 (5.298 mg/mL) μL/mL, respectively. Previous studies have investigated the inhibitory effect of other natural active substances on S. sonnei. The MIC of Centella asiatica extract against S. sonnei determined by Sieberi et al. (2020) was 52.1 mg/mL. The extract of Acacia nilotica at a concentration of 25 mg/mL had a slight inhibitory effect on S. sonnei (Banso, 2009). Javed et al. (2020) found that at a concentration of 100 mg/mL, tannin analogs from leaves of Salix alba had a significant inhibitory effect on S. sonnei. Compared with the above three natural active substances, the LC-EO had good inhibitory activity against S. sonnei.
The LC-EO exhibited an antibacterial effect against S. sonnei in LB broth in a concentration-depend manner. As shown in Figure 1, the growth of S. sonnei was completely inhibited at a concentration of 4 μL/mL of LC-EO, which was consistent with the result that the MBC of LC-EO against S. sonnei ATCC 25931 was 4 μL/mL. In addition, LC-EO also showed antibacterial activity against S. sonnei in contaminated lettuce leaves and juice (Fig. 9). Moreover, in this study, the number of bacteria decreased to below the detection limit (1 CFU/mL) after in LB broth, lettuce leaves, and lettuce juice with 4, 5, and 8 μL/mL of LC-EO treated for 1, 1, and 4 h, respectively.
Thus, the inhibitory effect of LC-EO on S. sonnei in different media was in the order of LB broth > lettuce leaves > lettuce juice, which may be caused by the kind of nutrients that vary among mediums and the potential correlations among minerals, vitamins, and other components with LC-EO (Kang et al., 2022).
The results (Fig. 3) revealed that the ROS level of LC-EO increased sharply after a 10-min treatment (p < 0.01), which was similar to the study of Song et al. (2021). Wang et al. (2021) demonstrated that thymoquinone increased the intracellular ROS level of Bacillus cereus in a time-dependent and concentration-dependent manner. Although ROS are often used as signaling molecules in the rich physiological metabolism of cells, high levels of ROS can cause a variety of intracellular damage, damaging organelles and membranes, thereby threatening membrane integrity (Jeon and Ha, 2020; Yu et al., 2016). Therefore, LC-EO may inhibit S. sonnei by increasing ROS.
ATP is the energy source for cells to perform basic functions, and the level of ATP in intact cells is stable (Tian et al., 2021). As shown in Figure 4, under the action of LC-EO, the ATP content of S. sonnei decreased in a concentrate-dependent manner, which was consistent with the result of 2.5 mg/mL of protocatechuic acid treating Yersinia enterocolitica for 2 h (Wu et al., 2022). It has been found that changes in cell membrane permeability may lead to a decrease in ATP concentration. Increased ATPase activity results in the release of ATP from cellular contents when bacterial cell membranes and walls are damaged (Cui et al., 2021). Overall, these studies suggested that LC-EO damages the cell membrane integrity of S. sonnei, leading to ATP leakage, while increased proton pump ATPase activity led to accelerated ATP hydrolysis, further leading to reduced ATP concentration.
SOD enzymes present in aerobic and anaerobic organisms are responsible for the breakdown of superoxide radicals and can be used to detect cellular defense responses to external stresses (Kong et al., 2013; Shirwaikar et al., 2007). Figure 5 shows that the activity of the SOD enzyme was significantly increased (p < 0.05) after being treated with LC-EO. Similarly, Zhu and Zhang (2020) found a significant increase in intracellular SOD enzyme activity in E. coli after acidophilus treatment. In the natural state, the ROS within the bacteria are in a very low dynamic equilibrium (Song et al., 2016). When ROS are produced in large quantities, bacteria produce more antioxidants, thus enhancing the defense system and reducing oxidative damage (Lushchak, 2011). Therefore, the enhanced SOD enzyme activity indicated that S. sonnei was in a state of oxidative stress during the treatment with LC-EO.
MDA, one of the products of lipid peroxidation, was quantified as the indicator of lipid peroxidation of bacterial cell membranes (Luo et al., 2022; Xu et al., 2018b). In the present study, the MDA levels in Figure 6 were observed to show the same trend as the ROS levels in Figure 3. Alkyl gallate ester (0.4 mM) treatment significantly increased (p < 0.05) 1.45-fold MDA in E. coli, which was thought to be caused by the ·OH radical acting as an initiator of lipid peroxidation (Shi et al., 2021). Under oxidative stress conditions, excessive accumulation of ROS in bacterial cells may cause damage to important molecules such as lipids, proteins, and DNA, thereby posing a threat to bacterial cells and even causing bacterial death (Liu et al., 2022; Shi et al., 2021). Thus, LC-EO promoted lipid oxidative damage by exposing S. sonnei to oxidative stress and increasing the intracellular ROS levels in bacteria.
The changes of cell membrane integrity of S. sonnei cells with or without LC-EO treatment were measured via flow cytometry. The results revealed that while the control group maintained a higher proportion of viable cells, S. sonnei exposed to the LC-EO had a greater proportion of damaged cells (Fig. 7). A similar study showed that about 44.51% of bacterial cell membrane integrity of B. cereus was damaged after 0.25 mg/mL of saponins from Chenopodium quinoa Willd treatment (Dong et al., 2020). Shan et al. (2020) treated Vibrio parahaemolyticus with 10 μM of novel antimicrobial peptides extracted from zebra fish for 30 min, and 95.34% of V. parahaemolyticus cell membrane integrity was impaired.
Studies have also shown that the accumulation of ROS can lead to the oxidation of proteins and fats, thereby promoting oxidative damage to cell membranes and disrupting cell membrane integrity (Jeon and Ha, 2020). This suggested that the destruction of LC-EO to the cell membrane integrity of S. sonnei may be caused by the irreversible damage caused by the increase of ROS level.
In this study, FESEM was used to observe obvious shrinkage in S. sonnei cells after treatment with LC-EO (Fig. 8). Guo et al. (2020) observed that the cell membranes of Candida sakazakii exposed to coenzyme Q 0 were rough and had obvious depressions. A study by Kang et al. (2018) showed that Shigella flexneri exposed to gallic acid at MICs had a fragmented cell morphology with leaked cellular components. In summary, the results showed that LC-EO leads to cell membrane damage by causing serious changes in cell membrane permeability, which further leads to leakage of S. sonnei contents and changes in bacterial morphology, causing irreversible damage to bacteria.
Simple processed foods such as fresh-cut produce are popular for their convenience and freshness; however, these products are often eaten raw, making them also a source of foodborne illness (Pang et al., 2017). In this study, LC-EO was observed to have antibacterial activity against S. sonnei in contaminated lettuce leaves and lettuce juice (Fig. 9), and it did not cause negative effects on the quality of lettuce leaves (Table 1). Similarly, Dai et al. (2021) showed that 1 mg/mL of LC-EO did not affect the appearance and color of bitter gourd juice, carrot juice, cucumber juice, and spinach juice within 4 d. These results indicated that LC-EO was a natural bacteriostatic agent without reducing the quality of lettuce.
Conclusion
In summary, this study indicated that the antibacterial activity of S. sonnei was influenced significantly by LC-EO. Among them, the mechanism of action of LC-EO on S. sonnei may be that LC-EO affected the intracellular enzyme activity of bacteria. S. sonnei was in a state of oxidative stress, which led to an increase in intracellular SOD enzyme activity, while the ROS level and MDA content increased. Excessive ROS led to impaired cell membrane integrity and leakage of intracellular material (decreased ATP content), which eventually led to cell apoptosis and bacterial atrophy. In addition, LC-EO showed a great antibacterial activity also in food matrix (lettuce leaves and lettuce juice), and there is no significant negative impact on the quality of lettuce. Therefore, it shows that the LC-EO has great potential for application in food industry.
Footnotes
Acknowledgment
We thank Dr. Zhang Guoyun (State Key Laboratory of Crop Stress Biology for Arid Areas, Northwest A&F University, Yangling, China) for FESEM experimental assistance.
Authors' Contributions
X.B.: Conceptualization, methodology, formal analysis, project administration, writing—original draft, and writing—review and editing. T.C.: Resources, investigation, data curation, and visualization. X.L.: Resources, methodology, and investigation. Z.L.: Data curation, resources, and methodology. R.M.: Formal analysis and resources. R.S.: Methodology, supervision, and software. X.L.: Visualization, formal analysis, and validation. X.L. and X.X.: Supervision and project administration. C.S.: Project administration, supervision, and funding acquisition. All the authors have read and agreed to the published version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (32272445 and 31801659) and the class General Financial Grant from the Shaanxi Postdoctoral Science Foundation (2018BSHEDZZ150).