
Abstract
In this study, we investigated the inhibitory effects of coenzyme Q0 (CoQ0) on biofilm formation and the expression of virulence genes by Cronobacter sakazakii. We found that the minimum inhibitory concentration of CoQ0 against C. sakazakii strains ATCC29544 and ATCC29004 was 100 μg/mL, while growth curve assays showed that subinhibitory concentrations (SICs) of CoQ0 for both strains were 6.4, 3.2, 1.6 and 0.8 μg/mL. Assays exploring the inhibition of specific biofilm formation showed that SICs of CoQ0 inhibited biofilm formation by C. sakazakii in a dose-dependent manner, which was confirmed by scanning electron microscopy and confocal laser scanning microscopy analyses. CoQ0 inhibited the swimming and swarming motility of C. sakazakii and reduced its ability to adhere to and invade HT-29 cells. In addition, CoQ0 impeded the ability of C. sakazakii to survive and replicate within RAW 264.7 cells. Finally, real-time polymerase chain reaction analysis confirmed that nine C. sakazakii genes associated with biofilm formation and virulence were downregulated in response to CoQ0 treatment. Overall, our findings suggest that CoQ0 is a promising antibiofilm agent and provide new insights for the prevention and control of infections caused by C. sakazakii.
Introduction
C
The most common outcomes of foodborne C. sakazakii infection in infants are necrotizing enterocolitis, bacteremia, and meningitis (Drudy et al., 2006; Li et al., 2016), often leading to neurological sequelae and even death (Chandrasekaran et al., 2018). Recently, the Centers for Disease and Control Prevention estimated that C. sakazakii infection case fatality rates are as high as 40% (Kalyantanda et al., 2015).
Biofilms are complex microbial communities that adhere to biotic or abiotic surfaces. Within the biofilm, microbes are firmly embedded in an extracellular matrix composed of polysaccharides, proteins, and DNA (Roy et al., 2018). This matrix acts as a physical barrier against chemical agents and provides a protective ecological niche, enhancing the survival of biofilm-associated microorganisms (Capita et al., 2019).
Given the public health importance of C. sakazakii, its ability to form a biofilm has been the subject of much research. C. sakazakii forms a biofilm on various surfaces, including glass, stainless steel, silicon, latex, and polycarbonate, all of which are used in the processing and preparation of milk powder (Coughlan et al., 2016). Biofilm-associated C. sakazakii show increased resistance to antibiotics, detergents, and other environmental stresses (Odeyemi and Sani, 2019). As such, compared with planktonic cells, biofilm-based C. sakazakii infections are significantly more difficult to resolve (Hall and Mah, 2017). In an industrial setting, biofilm formation by C. sakazakii increases the risk of contamination of foodstuffs, which has significant health and financial repercussions (Díez-García et al., 2012).
Virulence factors are gene-mediated molecules produced by microorganisms that enhance their ability to invade a host, cause disease, or evade host defenses (Chen et al., 2012). Functions, such as motility, are also considered virulence factors. C. sakazakii produces a variety of virulence factors, including proteins involved in motility, host cell adhesion and invasion (Shi et al., 2017), and replication and survival within macrophages (Holý et al., 2019). Motility is essential for the virulence of C. sakazakii, allowing it to pass through the intestine and colonize more favorable host environments such as mucous membranes, the gastric and intestinal epithelia, and endothelial tissues (Joseph and Forsythe, 2011; Kim and Loessner, 2008; Tobe et al., 2011). Outer membrane proteins, OmpA and OmpX, were shown to play an important role in C. sakazakii adhesion to and invasion of human intestinal epithelial Caco-2 and HT-29 cells (Kim et al., 2010).
Further study confirmed that OmpA contributes significantly to the pathogenicity of C. sakazakii as an essential factor in the invasion of various epithelial and endothelial cells of human and animal origin (Forsythe et al., 2014), whereas OmpX plays a crucial role in the basolateral invasion of host cells (Singh et al., 2015). The ability of C. sakazakii to survive and replicate inside immune cells such as macrophages is critical for establishing infection and is the first step in the development of severe illnesses such as sepsis and meningitis (Eshwar et al., 2015).
Over the past decade, the use of plant-derived compounds as alternative antimicrobials has gained significant attention as a result of increasing concerns over the safety of synthetic antimicrobial agents and the emergence of antibiotic-resistant bacteria (Pate et al., 2020). Coenzyme Q0 (CoQ0; 2,3-dimethoxy-5-methyl-1,4-benzoquinone; C9H10O4), a family of phanquinone compounds consisting of benzoquinone rings conjugated with isoprenoid chains, is a benzoquinone compound extracted from the fungus Antrodia camphorata (Cheng-Han et al., 2014). It has demonstrated antitumor, anti-inflammatory, and antiangiogenic properties (Yang et al., 2016), as well as antibacterial activity against Staphylococcus aureus and Listeria monocytogenes (Fan et al., 2018; Zhao et al., 2014). In addition, CoQ0 successfully inhibited biofilm formation by Salmonella enterica serovar Typhimurium (Yang et al., 2019).
Although multiple reports show that CoQ0 has inhibitory effects against a variety of microorganisms, few studies have examined the effects of CoQ0 on C. sakazakii biofilm formation or the expression of virulence factors. In the current study, we first determined the minimum inhibitory concentration (MIC) and subinhibitory concentration (SIC) of CoQ0 against several C. sakazakii strains. Crystal violet staining, scanning electron microscopy (SEM), and confocal laser scanning microscopy (CLSM) were then used to study the effects of SICs of CoQ0 on C. sakazakii biofilm formation. We then assessed the effects of CoQ0 on various C. sakazakii virulence traits, including motility, adhesion to and invasion of HT-29 cells, and survival and replication within macrophages. Finally, the expression of genes associated with biofilm formation and other virulence factors following CoQ0 treatment was examined using real-time polymerase chain reaction (RT-PCR) analyses.
Materials and Methods
Chemicals and reagents
CoQ0 (high-performance liquid chromatography ≥99%; CAS 605-94-7) was obtained from J&K Scientific Co., Ltd. (Beijing, China) and dissolved in dimethyl sulfoxide (DMSO) for use in all experiments. The final concentration of DMSO in all sample solutions was 0.1% (v/v), which has no apparent effect on the growth of C. sakazakii. All other chemicals were of analytical grade and were unaltered.
Bacterial strains and culture conditions
C. sakazakii strains ATCC 29004 and ATCC 29544 were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). Strain ATCC 29004, which is a relatively strong biofilm producer, was used in the biofilm assay. Before each assay, bacteria were inoculated onto tryptic soy agar (TSA) medium and incubated at 37°C for 12 h. To obtain fresh overnight cultures, a single colony was inoculated into 30 mL of tryptic soy broth (TSB) medium and incubated with shaking at 130 rpm for 12 h at 37°C. Following incubation, cultures were centrifuged (4°C, 8000 × g, 5 min), washed three times with sterile phosphate-buffered saline (PBS), and diluted in TSB medium to an optical density at 600 nm (OD600) of 0.5 [∼4 × 108 colony-forming units (CFU)/mL].
MICs and SICs determinations
The MICs and SICs of CoQ0 against C. sakazakii ATCC 29004 and ATCC 29544 were determined as described previously (Shi et al., 2018), with some modifications. Briefly, overnight bacterial culture was diluted 400 × in TSB medium (∼1 × 106 CFU/mL) before 125 μL of the diluted culture were added to individual wells of a 96-well plate. Equal volumes of CoQ0 solution were gently added to each well to achieve final CoQ0 concentrations of 0 (control), 0.8, 1.6, 3.2, 6.4, 12.8, 25.6, and 51.2 μg/mL. TSB medium containing 0.1% DMSO was used as the negative control. Plates were incubated at 37°C for 24 h, and cell growth was monitored at 600 nm at 1-h intervals using a microplate reader (Model 680; Bio-Rad, Hercules, CA, USA). The MIC of CoQ0 was defined as the lowest concentration at which there was no visible growth of C. sakazakii. SICs of CoQ0 were defined as concentrations at which no significant inhibition of C. sakazakii growth was observed.
Inhibition of specific biofilm formation assay
The inhibition of specific biofilm formation (SBF) assay was carried out using a Crystal Violet staining method as described previously (Naves et al., 2008), with minor modifications. Briefly, an overnight culture of C. sakazakii ATCC 29004 was diluted in TSB medium to an OD600 of 1.0. CoQ0 was added to culture aliquots to obtain final concentrations of 0 (control), 1.6, 3.2, and 6.4 μg/mL. Aliquots (200 μL) of the mixtures were then pipetted into individual wells of a 96-well plate. Uninoculated TSB containing 0.1% DMSO was used as the negative control. Plates were incubated statically at 37°C or 25°C for 24, 72, or 120 h and the absorbance of the mixtures was monitored using a microplate reader (Model 680; Bio-Rad) at 630 nm. Following incubation, bacterial cultures were aspirated and the plates were washed with 300 μL of distilled water before being air-dried for 30 min. The wells were stained with 250 μL of 1% (w/v) Crystal Violet (Tianjin Kermel Chemical Regent Co., Ltd, Tianjin, China) for 20 min and any excess stain was removed using sterile distilled water.
After drying for 30 min, 250 μL of 33% (v/v) glacial acetic acid was added to each well and the plates were shaken at ambient temperature for 20 min. The biofilm biomass in each well was quantified by measuring the OD at 570 nm. The SBF was determined from the ratio of the OD570 and OD630 values.
SEM observation
Confirm the effects of CoQ0 on biofilm formation, the C. sakazakii ATCC 29004 biofilms were further analyzed by SEM as described previously (Fan et al., 2018), with minor modifications. Briefly, C. sakazakii ATCC 29004 cultures (OD600 = 1.0) mixed with CoQ0 solution (6.4, 3.2, 1.6, or 0 μg/mL) were added to individual wells of a 24-well plate containing sterile glass slides of the same diameter as the bottoms of the wells. Culture without CoQ0 was used as a control. Following incubation at 25°C for 48 h, culture supernatants were removed. The samples were fixed with 2 mL of 2.5% (v/v) glutaraldehyde overnight at 4°C. The glass slides were then removed and washed with sterile PBS, followed by treatment with 1% (v/v) osmic acid at 4°C for 5 h. Subsequently, the glass slides were dehydrated using a graded ethanol series (30%, 50%, 60%, 70%, 80%, 90%, and 100%). After being dried and coated with gold, the slides were examined under a field emission scanning electron microscope (S-4800; Hitachi, Tokyo, Japan) at 4000 × and 1500 × magnification.
CLSM observation
To examine the effects of CoQ0 on the viability of biofilm-associated C. sakazakii ATCC 29004 cells, biofilms were next strained using the LIVE/DEAD BacLight Bacterial Viability Kit (Thermo Fisher Scientific, Waltham, MA, USA), consisting of SYTO 9 and Propidium Iodide (PI) dyes. Biofilms were cultured on stainless steel coupons in a 24-well plate in the presence or absence of CoQ0, as described in SEM observation. Following removal of the supernatant, the plate was washed twice with sterile water. Stainless steel coupons were then stained with STYO 9 and PI in the dark for 8 min as per the manufacturer's instructions. After being washed with sterile water, the stained biofilms on the stainless steel coupons were examined using a confocal laser scanning microscope (A1; Nikon, Tokyo, Japan).
Motility assay
Swimming and swarming motility assays were conducted as described previously (Li et al., 2014), with minor modifications. Bacterial swimming motility assays were conducted using 20 mL of LB broth [Luria-Bertani (broth)] containing 0.3% (w/v) agar, while swarming motility assays were conducted in 20 mL of LB broth supplemented with 0.5% (w/v) agar and 0.5 (w/v) glucose. CoQ0 was added to the warm media (45°C) to achieve final concentrations of 0 (control), 1.6, 3.2, and 6.4 μg/mL, and the resulting mixtures poured into Petri dishes. The resulting agar plates were dried at ambient temperature for 1 h. Aliquots (5 μL) of bacterial cultures (OD600 = 0.5) were then inoculated onto the center of each plate and incubated upright at 37°C for 7 h. Images of the resulting bacterial halos and swarm areas were obtained using a Gel Imaging System (Bio-Rad).
Cell culture
HT-29 human colon carcinoma cells (ATCC) were maintained in Dulbecco's modified Eagle's medium/Nutrient Mixture F-12 (DMEM/F-12) (Gibco, Grand Island, NY, USA). RAW 264.7 murine macrophage cells (ATCC) were cultured in DMEM (Gibco). Culture media were prepared as described by Shi et al. (2018). Cell lines were maintained at 37°C in a humidified 5% CO2 atmosphere.
Adhesion and invasion assay
The effects of CoQ0 on bacterial adhesion and invasion were examined as described previously (Moroni et al., 2006), with some modifications. For both assays, HT-29 cells in DMEM/F-12 were inoculated into 24-well plates (105 cells/well) and incubated overnight before being washed twice with sterile PBS. Aliquots of C. sakazakii ATCC 29544 culture (OD600 = 0.5) were mixed with CoQ0 solution (6.4, 3.2, 1.6, 0.8, or 0 μg/mL final concentration) and cultured at 37°C for 6 h. Bacterial cultures were then washed with sterile PBS and the cell pellets resuspended in TSB medium to an OD600 of 0.5 (∼4 × 108 CFU/mL). Cultures were diluted 40 × in DMEM/F-12 and inoculated onto the HT-29 cell monolayers at a multiplicity of infection (MOI) of 10. Following centrifugation (600 × g, 5 min), the plates were incubated in a humidified, 5% CO2 incubator at 37°C for 2 h.
For adhesion assays, the incubated plates were rinsed three times with sterile PBS and the cells lysed by the addition of 1 mL of 0.1% (v/v) Triton X-100 (Amresco, Solon, OH, USA) followed by incubation at 4°C for 20 min. The lysed cells were then serially diluted in sterile PBS and plated on TSA for colony counting. For invasion assays, the incubated plates were washed once with sterile PBS and 1 mL of DMEM/F-12 containing gentamicin (100 μg/mL; Amresco) was added to each well. The plates were then incubated for a further 45 min to kill the extracellular bacteria. Following incubation, the cells were washed three times before being lysed by the addition of 1 mL of 0.1% (v/v) Triton X-100 followed by incubation at 4°C for 20 min. The lysed bacteria were then serially diluted and plated on TSA for colony counting. Results were expressed as the percentage of colonies on the treatment plates relative to those on the control plates.
Intracellular survival and replication assay
The effects of CoQ0 on the intracellular survival and replication of C. sakazakii ATCC 29544 in RAW 264.7 cells were examined as described previously (Naves et al., 2008), with some modifications. RAW 264.7 cells cultured in DMEM were seeded into 24-well plates (105 cells/well) and incubated at 37°C in a humidified atmosphere with 5% CO2 for 16 h. Aliquots of bacterial suspension (OD600 = 0.5) were mixed with CoQ0 at a final concentration of 0 (control), 0.8, 1.6, 3.2, or 6.4 μg/mL and incubated at 37°C for 6 h. Following incubation, the suspensions were washed once with sterile PBS and the resulting cell pellets resuspended in TSB to an OD600 of 0.5 (∼4 × 108 CFU/mL). The cell suspensions were then diluted in DMEM to a density of 1 × 106 CFU/mL and inoculated onto the RAW 264.7 cell monolayers at a MOI = 10. Following incubation at 37°C in the presence of 5% CO2 for 2 h, the plates were rinsed once with sterile PBS, and 1 mL of DMEM containing gentamicin (100 μg/mL) was added to each well. The plates were then incubated for a further 45 min.
For the intracellular survival assays, the incubated plates were rinsed with sterile PBS and the cells lysed by the addition of 1 mL of 0.1% (v/v) Triton X-100 followed by incubation at 4°C for 20 min. The lysed cells were serially diluted before being plated on TSA plates. The results are expressed as the number of viable C. sakazakii cells (CFU/mL) before and after CoQ0 treatment. Infected cells that were not treated with CoQ0 were used as the control. For the intracellular replication assay, the incubated plates were again rinsed with sterile PBS, and 1 mL of DMEM containing gentamicin (10 μg/mL) was added to each well. Following incubation at 37°C in the presence of 5% CO2 for either 24 or 48 h, the cells were lysed, serially diluted, and plated on TSA plates, as described in the intracellular survival assay.
Isolation of RNA and RT-PCR assays
To assess the effects of CoQ0 on the expression of virulence genes (Table 1), C. sakazakii ATCC 29004 was cultured in the presence (1.6 or 3.2 μg/mL) or absence (control) of CoQ0 at 37°C for 8 h. Total RNA was then extracted from each culture using the RNA Prep Pure Cell/Bacteria Kit (Tiangen, Beijing, China) as per the manufacturer's instructions. The quality and concentration of the extracted RNA were determined using a nucleic acid and protein spectrophotometer (Nano-200; Aosheng Instrument Co., Ltd., Hangzhou, China).
Primers Used in This Study
RNA was then reverse transcribed into cDNA using the TaKaRa PrimeScript RT Reagent Kit (TaKaRa, Dalian, China) as per the manufacturer's instructions. RT-PCR assays were carried out in 25-μL reaction volumes with SYBR Green reagents (TaKaRa) using an IQ 5 Multicolor Real-Time PCR Detection System (Bio-Rad). The reaction parameters included an initial denaturation at 95°C for 30 s, followed by 40 cycles of 95°C for 5 s and 60°C for 30 s, with a dissociation step of 95°C for 15 s and 60°C for 30 s. Specific primers corresponding to each of the target genes (Table 1) were based on those from previous reports (Dai et al., 2022; Forsythe et al., 2014).
The 16S rRNA gene was used as an internal control for normalization of gene expression. The 2−ΔΔCt method was used to compare the expression of genes from different samples (Wei et al., 2012). The means and standard deviations (SD) were calculated from triplicate experiments.
Statistical analyses
All experiments were carried out independently three times. The data were presented as mean ± SD and analyzed using SPSS 23.0 software (IBM, New York, NY, USA). A Student's t-test was used to analyze differences between means. Differences were considered statistically significant at p < 0.05 and extremely significant at p < 0.01.
Results
Minimum inhibitory concentrations
CoQ0 showed strong antibacterial activity against C. sakazakii ATCC 29004 and ATCC 29544, with an observed MIC of 100 μg/mL for both strains (data not shown).
SICs and growth curve analyses
The effects of CoQ0 on the growth of C. sakazakii strains ATCC 29004 and ATCC 29544 are shown in Figure 1. At a CoQ0 concentration of 51.2 μg/mL, the lag phase of both C. sakazakii cultures was longer compared with the control culture grown in the absence of CoQ0. However, the growth curves of C. sakazakii ATCC 29004 and ATCC 29544 treated with 6.4, 3.2, 1.6, or 0.8 μg/mL CoQ0 were not significantly different from that of the untreated control (p > 0.05). Therefore, these concentrations were used as SICs in subsequent biofilm formation, virulence, and RT-PCR assays.
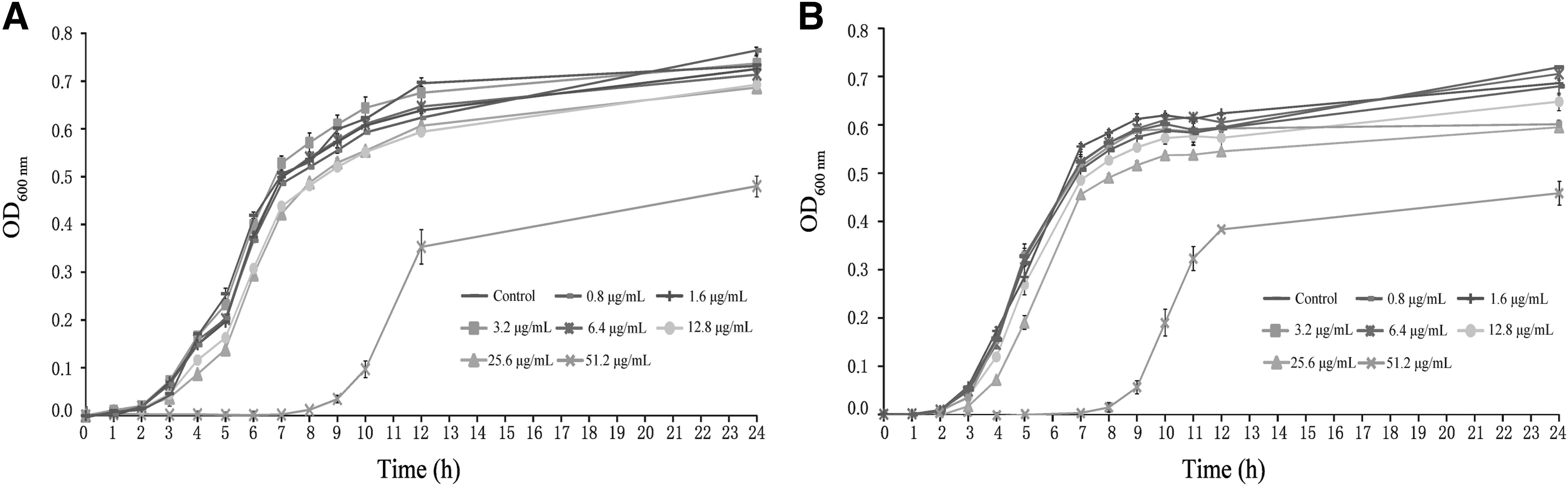
Growth curves of Cronobacter sakazakii ATCC 29004
Effects of CoQ0 on C. sakazakii biofilm formation
The effects of CoQ0 on the ability of C. sakazakii ATCC 29004 to form biofilms following incubation at 25°C or 37°C for 24, 72, or 120 h are shown in Figure 2. The total biofilm biomass was significantly (p < 0.05) decreased compared with the control following treatment with CoQ0 at 6.4 μg/mL and incubation at 25°C (Fig. 2A). At 37°C, biofilm biomass was significantly (p < 0.05) decreased compared with the control at a CoQ0 concentration of 1.6 μg/mL, with further decreases in biomass observed with increasing CoQ0 concentration (Fig. 2B).
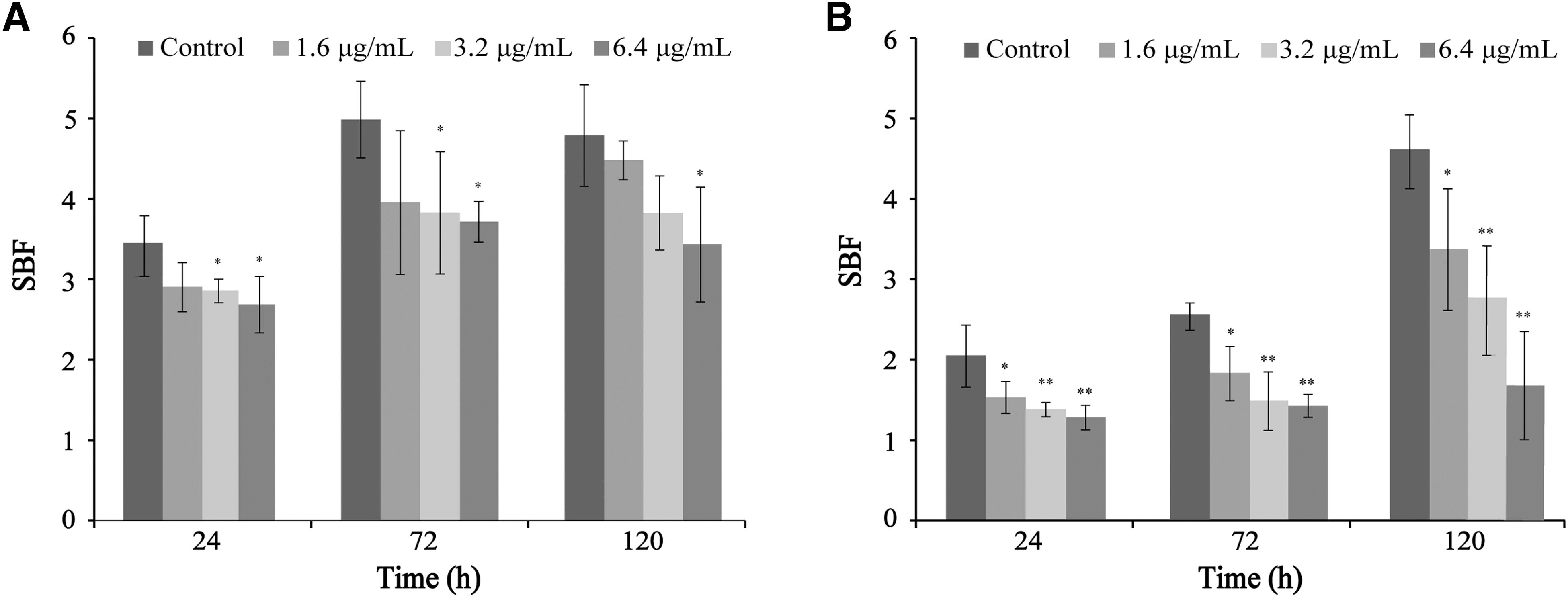
Effects of CoQ0 on Cronobacter sakazakii ATCC 29004 biofilm formation at 25°C
SEM observation
SEM images of CoQ0-treated C. sakazakii biofilms at 4000 × and 1500 × magnification are shown in Figure 3. In the absence of CoQ0, the biofilms exhibited a typical three-dimensional morphology with thick aggregates. In comparison, the biofilms became monolayers and cell clusters became looser with increasing CoQ0 concentrations, indicative of significant disruption.
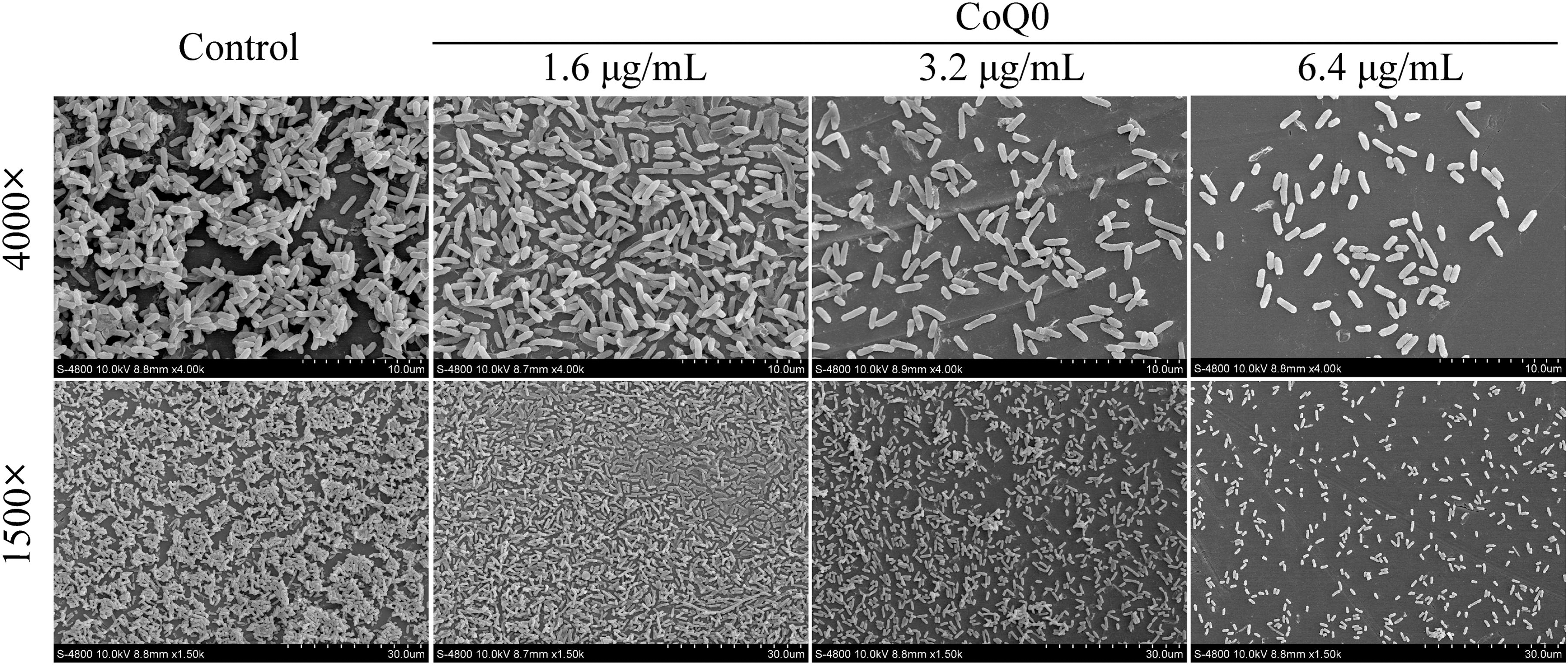
SEM micrographs of Cronobacter sakazakii ATCC 29004 biofilms formed on glass surfaces in the presence and absence of SICs of CoQ0. Magnification, 4000 × and 1500 × . Magnification is shown on the left and CoQ0 concentration on the upper side of the figure. CoQ0, coenzyme Q0; SEM, scanning electron microscopy; SICs, sub-inhibitory concentrations.
CLSM observation
As shown in Figure 4, the untreated biofilm appeared almost completely green, indicative of live cells, with few bacteria exhibiting PI fluorescence. However, increasing levels of red fluorescence (PI) were observed with increasing concentrations of CoQ0, suggesting a breakdown in membrane integrity. The highest level of red fluorescence intensity was observed at a CoQ0 concentration of 6.4 μg/mL (Fig. 4d).

CLSM images of Cronobacter sakazakii ATCC 29004 biofilms following treatment with CoQ0 at 0
Motility
As shown in Figure 5, CoQ0 treatment resulted in a decrease in the swimming motility of C. sakazakii compared with the untreated control, with the observed decreases in motility occurring in a dose-dependent manner. Similarly, the swarming motility of C. sakazakii decreased compared with that of the control in response to increasing CoQ0 concentration (1.6–6.4 μg/mL).

Effects of SICs of CoQ0 on the swimming
Adhesion to and invasion of HT-29 cells
We next examined the effects of CoQ0 on the ability of C. sakazakii ATCC 29544 to adhere to and invade HT-29 cells, and the results are shown in Figure 6. As shown in Figure 6A, CoQ0 significantly (p < 0.01) impeded the ability of C. sakazakii ATCC 29544 to adhere to host cells, with the decreases in adherence compared with the control occurring in a dose-dependent manner. At 0.8, 1.6, 3.2, and 6.4 μg/mL, CoQ0 treatment decreased the adherence of C. sakazakii ATCC 29544 by 36.8%, 39.3%, 40.2%, and 56.2%, respectively, compared with the control. Furthermore, CoQ0 at concentrations ≥1.6 μg/mL significantly (p < 0.05) inhibited the ability of C. sakazakii ATCC 29544 to invade host cells (Fig. 6B). Specifically, 5.3%, 21.2%, 34.8%, and 42.8% decreases in the percent relative invasion of host cells were observed at CoQ0 concentrations of 0.8, 1.6, 3.2, and 6.4 μg/mL, respectively.
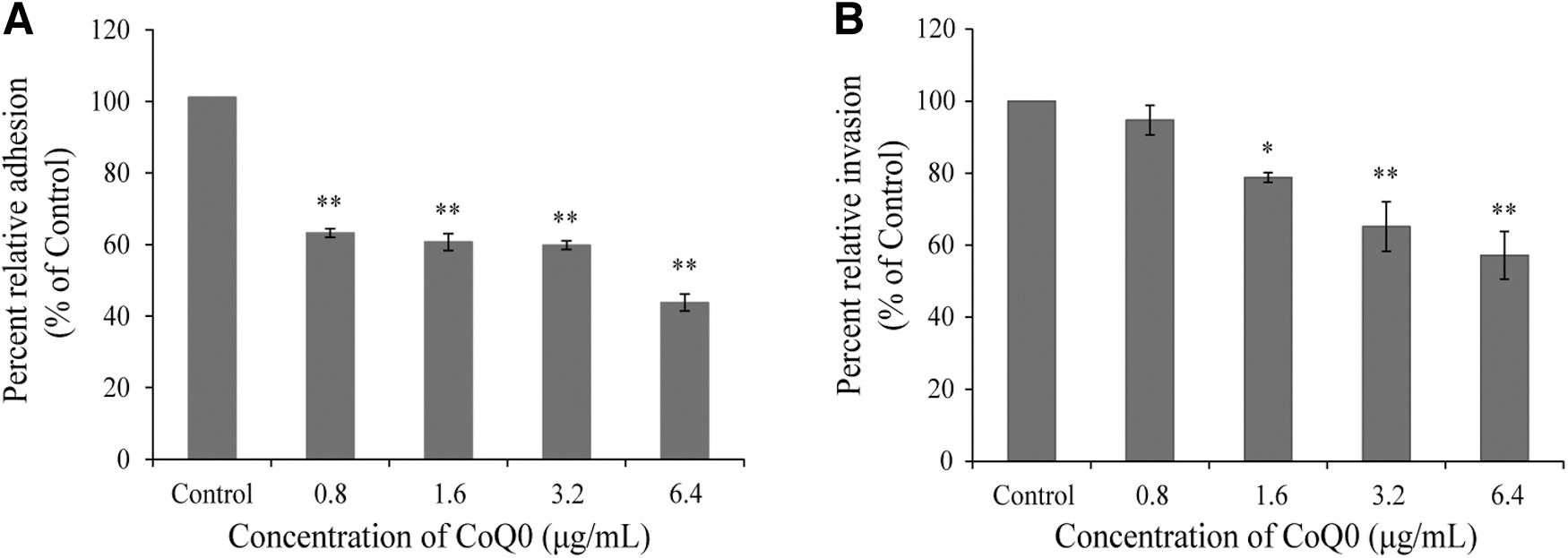
Effects of CoQ0 on the adhesion to
Intracellular survival and replication in RAW 264.7 cells
The effects of CoQ0 on the intracellular survival and replication of C. sakazakii in RAW 264.7 cells are shown in Figure 7. At concentrations of 3.2 and 6.4 μg/mL, CoQ0 significantly (p < 0.05) decreased the intracellular survival of C. sakazakii ATCC 29544 compared with the control at 48 h postinoculation. However, at CoQ0 concentrations of 0.8 and 1.6 μg/mL, the intracellular survival and replication of C. sakazakii ATCC 29544 did not significantly differ from the control at 24 and 48 h postinoculation.
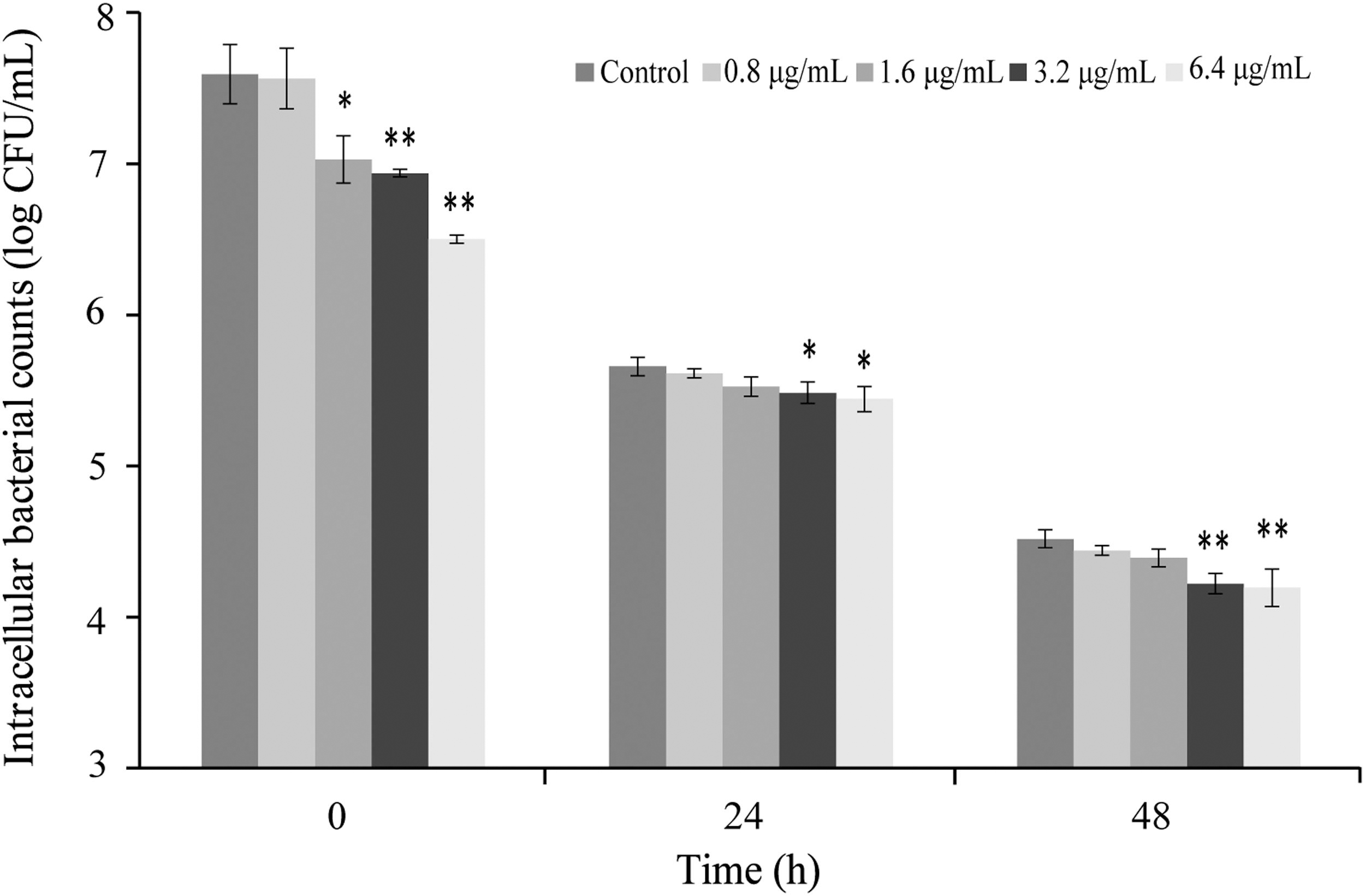
Inhibitory effects of CoQ0 the intracellular survival and replication of Cronobacter sakazakii ATCC 29544 in RAW 264.7 cells. Values are the mean ± SD of three independent experiments. *p < 0.05; **p < 0.01 versus the control. SD, standard deviation.
RT-PCR analyses
The RT-PCR analyses are shown in Figure 8. The results showed that following treatment with CoQ0 at 3.2 μg/mL, the expression of uvrY (adherence and invasion) in C. sakazakii cells was significantly (p < 0.05) downregulated compared with the untreated control (Fig. 8). Similarly, the mRNA levels of flgJ and fliD (flagellar assembly), motA and motB (flagellar motor proteins), bcsG (cell biosynthesis and biofilm formation), bcsA (cellulose synthase operon), and ompX and lpxB (LPS biosynthesis) were significantly (p < 0.01) lower than those in the control. However, no differences in the levels of transcription of flgJ, uvrY, and motB compared with the control were observed at CoQ0 concentrations ≤1.6 μg/mL.

Effects of CoQ0 on the relative transcription of virulence-associated genes in Cronobacter sakazakii ATCC 29004. *p < 0.05; **p < 0.01 versus the control.
Discussion
C. sakazakii, with its array of virulence factors, poses a significant threat to the health of infants and young children because of its ability to form biofilms on surfaces in facilities that process milk and dairy products (Beuchat et al., 2009). With the global increase in antibiotic resistance and safety concerns surrounding the use of chlorine-based disinfectants, the antibacterial properties of plant-derived compounds are increasingly being investigated (Kang and Song, 2018; Siroli et al., 2015; Zhou et al., 2021). CoQ0, derived from A. cinnamomea, has attracted attention for its potential as a natural food preservative and as a therapeutic antibiotic (Cheng-Han et al., 2014). Based on growth curve analysis, we identified appropriate SICs of CoQ0 that had no effect on growth for use in subsequent assays. These experiments showed that at SICs, CoQ0 has significant antibiofilm and antibacterial activities that cannot be attributed to inhibition of bacterial growth, indicating that low concentrations of CoQ0 affect bacterial virulence factors instead.
Biofilm formation contributes to the survival of C. sakazakii in suboptimal environments, is involved in evading and circumventing the host immune system, and provides protection against antibiotics and disinfectants (Povolotsky and Hengge, 2012). Therefore, identifying potential antibiofilm strategies has been a major focus for controlling the growth of C. sakazakii (Capita et al., 2019). In the current study, the effects of CoQ0 on the formation of C. sakazakii biofilms was first evaluated through Crystal Violet staining. The results showed that at SICs, CoQ0 significantly reduced C. sakazakii biofilm biomass at 25°C (normal room temperature) and biofilm formation at 37°C (the optimal temperature for C. sakazakii growth) (Fig. 2). In addition, CoQ0 significantly reduced the expression of genes related to biofilm formation, including bcsA and bcsG (Fig. 8). bcsA encodes the catalytic subunit of cellulose synthase, while bcsG encodes a conserved hypothetical protein involved in cellulose biosynthesis. Cellulose is an important component of the extracellular matrix of C. sakazakii biofilms and is essential for biofilm formation (Zheng et al., 2020).
Previous studies have shown that cell-free supernatants derived from goat milk-origin lactobacilli cultures can prevent the formation of C. sakazakii biofilms (Singh et al., 2019), while pomegranate and rosemary extracts were shown to have a synergistic effect in combination with traditional antibiotics against biofilm formation by Pseudomonas aeruginosa (El-Wafa et al., 2020). Shi et al. (2018) reported that thymoquinone significantly inhibited the biofilm formation of C. sakazakii by reducing the production of cellulose. Therefore, our results suggest that CoQ0 reduces the transcription of genes required for cellulose biosynthesis, ultimately inhibiting C. sakazakii biofilm formation.
With increasing CoQ0 concentration, C. sakazakii biofilms contained fewer cells overall and a higher percentage of damaged and dead cells compared with the control (Fig. 3). These SEM- and CLSM-based findings supported the results of quantitative Crystal Violet staining analysis, confirming that CoQ0 inhibits C. sakazakii biofilm formation. Similarly, Yang et al. (2019) used SEM analysis to show that shikimic acid (0.3125 mg/mL) effectively inhibits the formation of Salmonella Typhimurium biofilms, while CLSM observations by Nair et al. (2018) demonstrated that selenium damages Escherichia coli O157:H7 biofilms and decreases live cell counts.
Bacterial invasion of host cells requires motility, which is an important factor in the early stages of infection (Zhu et al., 2021). Swimming motility allows bacteria to move through liquid media, while swarming and clustering are observed in broth/semisolid media and on solid surfaces/agar, respectively (Odeyemi and Sani, 2019). Our findings demonstrated that both the swimming and swarming motilities of C. sakazakii were decreased in a concentration-dependent manner compared with the control as a result of CoQ0 treatment (Fig. 5). Consistent with this observation, motility-associated genes, motA, motB, flgJ, and fliD, were significantly downregulated compared with the control following CoQ0 treatment (Fig. 8). Other researchers have shown that the swimming and swarming motility of L. monocytogenes can be significantly reduced compared with morin-treated controls following treatment with a Bifidobacterium-derived bacteriocin (Dai et al., 2021), while the inhibition of flagellum biosynthesis and function as a result of citral treatment helped reduce the motility of C. sakazakii (Shi et al., 2017).
Flagella are the main structures responsible for bacterial motility and are essential for maximum virulence (Proudy et al., 2008). fliD and flgJ encodes the flagellum capping protein and a muramidase involved in flagella rod assembly, respectively. Lan et al. (2015) reported that motA and motB forms a bicistronic operon encoding two proteins that form the flagellum matrix. Therefore, we predict that SICs of CoQ0 inhibit C. sakazakii flagellum biosynthesis, reducing its mobility and capacity to invade host cells.
The ability of C. sakazakii to adhere to and invade host cells, including various epithelial cell lines and brain endothelial cells, is an important virulence factor in the establishment of infection (Alsonosi et al., 2019). In C. sakazakii, outer membrane protein OmpX also contributes to host cell invasion. In the current study, we found that CoQ0 significantly inhibited the adhesion and invasion of C. sakazakii in HT-29 cells, likely by downregulating the transcription of ompX and uvrY, two genes that are positively correlated with bacterial adhesion/invasion of epithelial cells (Fig. 8).
Similarly, CoQ0 treatment downregulated the expression of adhesion/invasion-associated genes in L. monocytogenes, resulting in a significant decrease in the adherence to and invasion of Caco-2 cells (Fan et al., 2018). Pediococcus acidilactici K10, isolated from kimchi, was also shown to reduce the adherence of Salmonella Typhimurium KCTC 1925 and E. coli O157:H7 ATCC 35150 to HT-29 cells (Kim et al., 2019). In light of these findings, it is likely that CoQ0 plays a significant role in reducing C. sakazakii adhesion to and invasion of enterocytes by regulating associated gene expression.
Clinical C. sakazakii isolates have been observed to continuously reproduce in macrophages and microglia (Almajed and Forsythe, 2016). Similar studies have reported that C. sakazakii tolerates the intracellular environment of macrophages, hiding inside these cells to evade the immune response and, ultimately, invade other organs (Alsonosi et al., 2019; Kim et al., 2015). In the present study, SICs of CoQ0 prominently inhibited the survival and replication of C. sakazakii inside macrophages (Fig. 7), supporting findings by Yang et al. (2019) who reported that the ability of Salmonella Typhimurium to survive and reproduce intracellularly was impacted by CoQ0 treatment. Based on these findings, we speculate that CoQ0 can reduce the ability of C. sakazakii to overcome host barriers and evade the immune response.
In summary, the current study confirms that CoQ0 is an ideal antibiofilm and antivirulence agent. We demonstrated that SICs of CoQ0 effectively inhibited the initial formation of biofilms at 25°C and 37°C, prevented the adherence of the biofilm to glass slides, and caused significant collapse of the biofilm structure. In addition, CoQ0 inhibited the swimming and swarming motilities of C. sakazakii cells and prevented their adhesion to and invasion of HT-29 cells. As well as decreasing the intracellular survival and replication of C. sakazakii in RAW 264.7 macrophages, SICs of CoQ0 downregulated the transcription of virulence-associated genes. Altogether, these activities significantly impacted the virulence of C. sakazakii. Our findings suggest that CoQ0 is a promising broad-spectrum antivirulence therapeutic agent that could be used to control C. sakazakii biofilm pollution and provide new avenues for the prevention and control of infections.
Footnotes
Acknowledgment
The authors thank Dr. Zhang Guoyun (State Key Laboratory of Crop Stress Biology for Arid Areas, Northwest A&F University, Yangling, China) for SEM experimental assistance.
Authors' Contributions
C.S., N.G., and Y.S. conceived and designed the experiments. N.G., Y.S., H.T., Y.Y., and J.L. performed the experiments. N.G., Y.S., and D.G. analyzed the data. C.S., X.W., and Z.S. contributed reagents, materials, and analysis tools. N.G. and Y.S. wrote the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (32272445 and 31801659), and the class General Financial Grant from the Shaanxi Province Postdoctoral Science Foundation (2018BSHEDZZ150).