
Abstract
Listeria monocytogenes (Lm) mainly infect pregnant women, children, the elderly, and other populations with low immunity causing septicemia and meningitis. Healthy people can tolerate higher doses of Lm and only cause gastrointestinal symptoms such as abdominal pain and diarrhea after infection. Compared to the above population, healthy people have a richer and more diverse gut microbiota. In this study, we show that the microbiota in the large intestine and the feces of mice can significantly inhibit the growth of Lm compared to the microbiota in the small intestine. Bacteria larger than 1 μm in the gut microbiota play an important role in inhibiting Lm growth. 16s rRNA sequencing results show that these bacteria are mainly composed of Clostridiales under the phylum Firmicutes, including Ruminiclostridium, Butyricicoccus, Lachnoclostridium, Roseburia, Coprooccus, and Blautia. Thus, we demonstrate that there are some potential functional bacteria in the gut microbiota that can increase resistance against Lm.
Introduction
L
The protective role of stable and healthy gut microbiota has been demonstrated in the setting of pathogen invasion. Destruction of community structure and function alters the balance between the microbiota and the host, resulting in altered susceptibility to infection (Becattini et al., 2017; Koren et al., 2012). The gut microbiota can resist the invasion of pathogens by direct and indirect ways.
Indirectly, the microbiota can stimulate host mucosal immunity against invasion and infection by pathogenic bacteria, and the development, function, and response of the mucosal immune system are influenced by the presence of the native microbiota (Round and Mazmanian, 2009). For example, commensal bacteria interact with host factors to protect the host from infection by selectively sorting cargo in Paneth cells, and depletion of the microbiota treated with antibiotics can lead to lysosomal degradation of lysozyme, which increases the invasion of bacteria Lm (Zhang et al., 2015). Directly, commensal microbes can inhibit potential pathogens by producing bacteriocins and depleting nutrients (Buffie and Pamer, 2013).
It is now appreciated that different bacterial species provide resistance to specific pathogens through different mechanisms. For example, Jacobson showed that Bacteroides spp. mediate colonization resistance to Salmonella Typhimurium infection through the production of the short-chain fatty acid propionate, which directly limits pathogen growth by disrupting intracellular pH homeostasis (Jacobson et al., 2018). Piewngam showed that Bacillus bacteria eliminate Staphylococcus aureus by inhibiting S. aureus quorum sensing (Piewngam et al., 2018).
Furthermore, Bacteroides spp. products did not limit the growth of other pathogens such as Lm, Yersinia pseudotuberculosis, or Clostridioidies difficile (Jacobson et al., 2018). Likewise, it was shown that the Clostridiales order isolated at house from the intestinal contents of mice exerts antilisterial activity in vitro and confers resistance in vivo after transfer to germ-free mice, but that the consortium of commensal bacteria did not limit growth from a laboratory strain of Escherichia coli (DH-5α) (Becattini et al., 2017).
Commensal bacteria can be developed as probiotics to help prevent and even treat Listeria gastrointestinal Listeriosis. The preparations of commensal bacteria from the cecal contents of healthy adult broiler chickens have been used successfully in the poultry industry to prevent the spread of human pathogens in chicken intestines (Kerr et al., 2013). Our hypothesis is that we can find more commensal bacteria to resist or even treat Lm infection. In this study, we isolated the gut microbiota and metabolites from the feces, and the microbiota was divided into multiple levels by filter membranes to identify key components that play a role in colonization.
Gut microbiota filtered with 1 μm filter membrane cannot effectively inhibit Lm growth. Sequencing of the microbiota after filtration with more than 1 μm of filter membranes revealed that these bacteria are mainly composed of Clostridiales under the phylum Firmicutes. We identified functional bacteria that can inhibit the growth of Lm from the gut microbiota, suggesting that potential probiotics can be developed from the gut microbiota to resist infection by pathogens. This study may provide new insights in identifying functional bacteria from the gut microbiota to resist pathogen infections.
Materials and Methods
Ethics statement
This study was carried out in accordance with the principles of the Basel Declaration and recommendations of the Institutional Animal Care and Use Committee [IACUC], (the Tongji University School of Medicine). The protocol was approved by the (local ethics committee of Tongji University School of Medicine).
Animals
Six-week-old male specific pathogen-free (SPF) BALB/c mice purchased from Shanghai Jie Si Jie Animal Laboratory Ltd., China, were used after 1 week of acclimatization. All animals received laboratory chow and water.
Feces filtration
Fecal materials from mice were resuspended in reduced phosphate buffered saline (PBS) solution (100 mg/mL) and centrifuged at 1000°C for 4 min. Then, the supernatant was filtered through micropore filters of 5, 3, 1, 0.45, and 0.22 μm at a time, and the filtrate (100 μL/well) obtained each time was co-cultured with Lm using plates of 96 wells and supplemented with 200 μL Brian Heart Infusion (BHI) medium. In addition, the filtrate was sequenced for 16S.
Growth inhibition in vitro
Lm that was grown overnight was concentrated by centrifugation at 4°C and resuspended in PBS before adjustment to give an optical density at 600 nm (OD600) of 1. The corresponding Lm concentration at OD600 = 1 was 1010 colony-forming units (CFU)/mL, determined by plate count. Intestinal contents or fecal pellets were resuspended at 100 mg/mL in reduced PBS and 100 CFU or 104 CFU Lm were inoculated into 200 μL of the supernatant or into sterile filtered aliquots of intestinal contents using 96-well plates. In some experiment, Lm and gut microbiota were co-cultured and supplemented with 200 μL of BHI medium. Listeria survival was evaluated after 3, 6, 12, or 24 h. To determine the Lm concentration, the supernatant was serially diluted in PBS and plated on CHROMagar Listeria chromogenic agar plates.
DNA extraction and 16S sequencing
Microbial DNA was extracted from samples using the E.Z.N.A.® soil DNA Kit (Omega Bio-tek, Norcross, GA) according to the manufacturer's protocols. The V3–V4 hypervariable regions of the bacteria 16S rRNA gene were amplified with primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) by thermocycler PCR system (GeneAmp 9700; ABI). 16S sequencing and data processing were completed by Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China).
Raw fastq files were quality filtered by Trimmomatic and merged by FLASH with the following criteria: (1) reads were truncated at any site with an average quality score <20 in a 50 bp sliding window; (2) sequences with a >10 bp overlap and mismatch of no more than 2 bp were merged; (3) the sequences of each sample were separated according to barcodes (exact match) and primers (up to two mismatched nucleotides). Reads containing ambiguous bases were removed.
Operational taxonomic units (OTUs) were clustered with 97% similarity cutoff using UPARSE (version 7.1) with a novel “greedy” algorithm that performs chimera filtering and OTU clustering simultaneously. The taxonomy of each 16S rRNA gene sequence was analyzed by RDP Classifier algorithm against the Silva (v138) 16S rRNA database using a 70% confidence limit.
Statistical and data analysis
Statistical analysis of biological information is usually performed at the similar level of 97% of OTU. Significant differences between the two microbial communities were performed using GraphPad Prism version 7 with unpaired t-test with Welch's correction. Statistical analysis between multiple samples was performed using one-way analysis of variance (ANOVA). Differences were considered significant if the p-value was <0.05. All data are expressed as mean CFU/organ (log10) ± standard error of the mean. The data of colony count are representative of at least three independent experiments.
Results
The microbiota in different intestinal regions inhibit the growth of Lm
To determine which intestinal microbiota provide resistance against the growth of Lm, we collected microorganisms from the small intestine (SI), large intestine (LI), and feces and co-cultured them with Lm. Inoculation with 102 CFUs or 104 CFUs in different contents resulted in different inhibition results (Fig. 1A, B). At the low inoculum size, gut microbiota in the SI, LI, and feces killed Lm within 3 h (Fig. 1A). In contrast, the growth of Lm in sterilized intestinal contents was similar to that in the BHI medium, which was not inhibited (Fig. 1A).
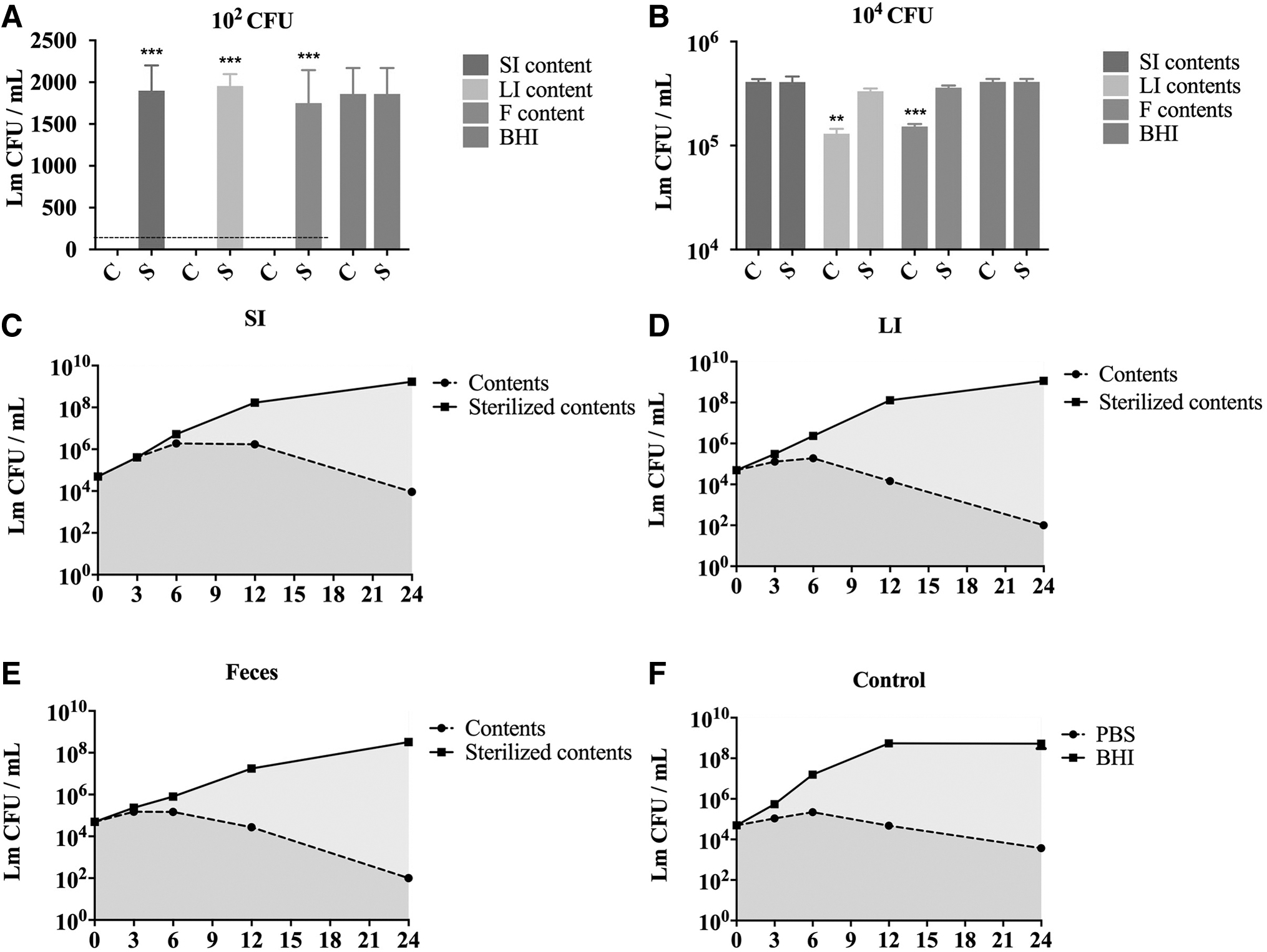
The gut microbiota inhibits the growth of Lm.
Around 3 h after inoculation with 104 CFUs, LI and feces contents inhibited Lm growth significantly compared to the BHI medium, with small intestinal contents having no significant effect in reducing Lm CFUs (Fig. 1B). Large intestinal contents and feces had more variable inhibitory activity after 24 h, with the number of Lm undetectable (<102 CFU) (Fig. 1D, E). In contrast, small intestinal contents demonstrated less activity in reducing Lm CFUs (Fig. 1C). The survival rate of Lm in the small intestinal contents was similar to that of PBS (Fig. 1C, F). These results suggest that the intestinal microbiota inhibits and possibly kill Lm, and there was a significant difference after 3 h of inoculation.
Microorganisms in the LI and feces exhibited the best inhibitory effect on Lm growth, and there were few differences between the two. Furthermore, Lm grew exponentially in the sterilized intestinal contents, indicating that there was no lack of nutrients in the initial suspension (Fig. 1C).
Inhibition of the gut microbiota of different concentrations on the growth of Lm
The composition and amount of microbiota in the gastrointestinal tract are not uniform. The concentration of bacteria in the stomach reached 102–104 bacteria/mL and the upper two-thirds of the SI, while the ileum has a concentration of 107–108 bacteria/mL and the colon has a concentration of 1010–1011 bacteria/mL (Aguirre et al., 2011; Stearns et al., 2011). To investigate the intestinal microbiota inhibition of different concentrations on the growth of Lm, 25, 50, 100, 250, and 500 mg/mL of feces content were, respectively, co-cultured with Lm for 24 h (Fig. 2A).
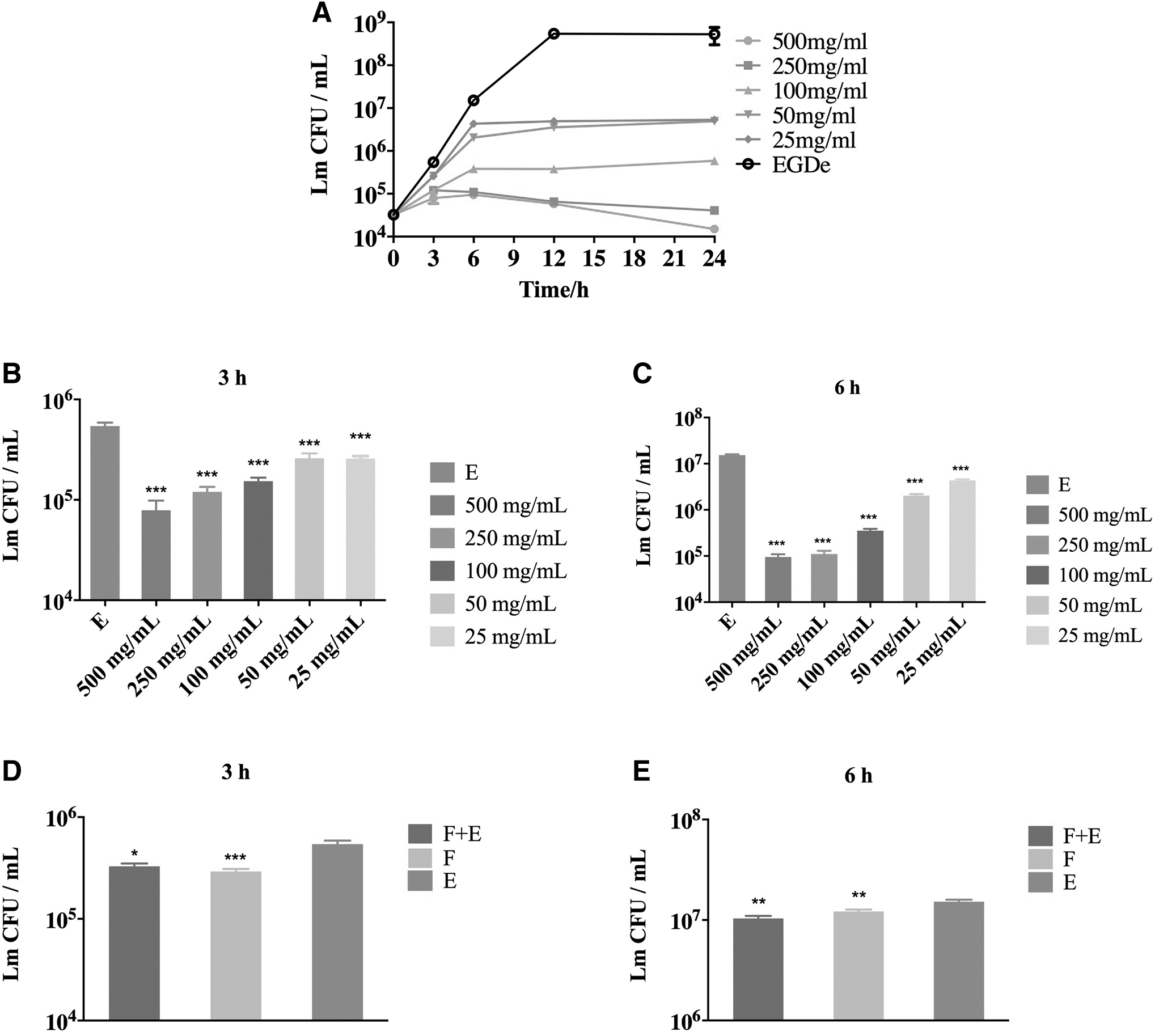
Inhibition of the gut microbiota of different concentrations on the growth of Lm.
Lm growth was significantly inhibited by all concentration of substances at 3 h postinoculation, and the higher the concentration, the better the inhibition effect (Fig. 2B). Results after 6 h of inoculation were similar to Figure 2B (Fig. 2C). Lm was killed at high content concentration (250 and 500 mg/mL) for 24 h after co-cultivation. In contrast, low concentration can only inhibit Lm growth, but cannot kill Lm completely (Fig. 2A).
Furthermore, the 100 mg/mL content mixed with 104 CFU Lm (group F+E) or just the content collected from feces (group F) was added to 20 mL of BHI medium and cultivated at 37°C under anaerobic conditions for 24 h. Thereafter, the medium was filtered through 0.22 μm micropore filters and the sterile filtrate was co-cultured with Lm (Fig. 2D, E). Lm growth was significantly inhibited by both filtrates, but inhibition of cultured content without Lm addition was better (Fig. 2D).
Intestinal microbiota prefiltrated with micropore filters inhibit the growth of Lm differently
To identify intestinal commensal species that provide resistance to colonization against Lm, we used filter membranes to multistage filter the bacteria in the contents and then co-culture with Lm, respectively (Fig. 3). In addition, the content that was filtered through the 0.22 μm micropore filter and cleared of bacteria was used to detect the inhibition of Lm growth by microbial metabolites. The filtrate (5 and 3 μm) can significantly control the growth of Lm after 6 h of inoculation (Fig. 4B, C). There was no increase in the number of Lm after 12–24 h of inoculation (Fig. 4A).

Experimental setup. Fecal materials (100 mg/mL) were filtered through micropore filters of 5, 3, 1, 0.45, and 0.22 μm at a time, and the filtrate obtained each time was co-cultured with Lm. The filtrate was sequenced for 16S. *The bacteria present in the filtrate were compared to the filtrate filtered with smaller micropore filters.
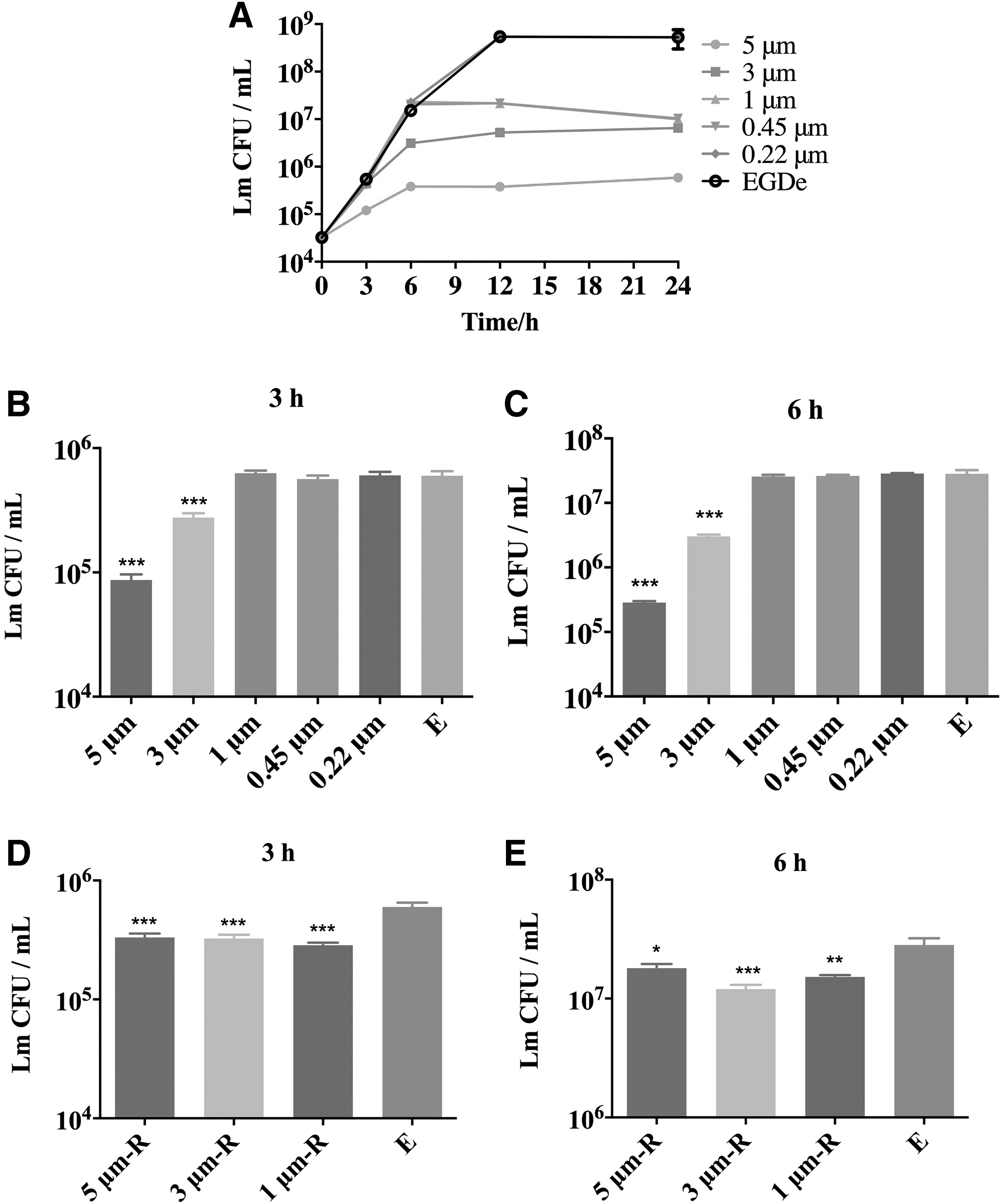
Inhibition of Lm growth by different filtrates.
In contrast, the filtrate filtered by membrane with aperture smaller than 3 μm (1 and 0.45 μm) did not inhibit the growth of Lm at 6 h after inoculation compared to the BHI medium (Fig. 4B, C). This demonstrated that intestinal bacteria smaller than 3 μm had no inhibitory effect on Lm growth. Interestingly, the number of Lm also did not increase in the 1 μm group after 12–24 h of inoculation (Fig. 4A). The possible reason was that unfiltered intestinal bacteria competed with Lm for nutrition after 12 h of growth. Furthermore, the Lm growth curve in content filtered through a 0.22 μm micropore filter was the same as in the BHI medium. The result suggests that microbial metabolites had no inhibitory effect on Lm growth.
To investigate whether bacteria larger than 1 μm could inhibit Lm growth, we removed the filter membranes from the filters that filtered the fecal slurry and washed the remaining bacteria trapped on the filter membranes with sterile PBS. Lm co cultured with the bacteria washed from the filter membranes and counted after 3 and 6 h, respectively (Fig. 4D, E). Lm growth was significantly inhibited by bacteria on three types of filter membranes compared to the BHI medium. Overall, our results suggest that there were specific bacterial strains that could inhibit Lm growth, and these bacteria were larger than 1 μm in size.
Identification of the order Clostridiales associated with inhibition from Lm growth
To identify intestinal commensal species that provide resistance to colonization against Lm, the filtrate was subjected to 16S rRNA gene sequencing to determine microbiota composition. The bacterial community at the phylum (Fig. 5A) and genus (Fig. 5B) levels showed that there were obvious differences in the composition of 3/5 μm filtrate and 1/0.45 μm filtrate. Similar to this, cluster analysis showed that the 5 μm filtrate and 3 μm filtrate clustered together, but those of the 1/0.45 μm filtrate located on different branches (Fig. 5D). LEfSe analysis demonstrated that the order Clostridiales in Firmicutes was relatively abundant in the 3/5 μm filtrate (Fig. 5C), including Ruminiclostridium, Butyricicoccus, Lachnoclostridium, Roseburia, Coprooccus, Lachnospiraeae_NK4A136_GROUP, Blautia, and Mucispirillum. (Fig. 5E). These results suggest that the order Clostridiales may play a potential role in inhibiting Lm growth.

Sequencing of 16S rRNA gene from the filtrate filtered with different membranes.

Discussion
Lm replicates in the intestinal villi and is eliminated in the gut lumen and disseminates to the spleen and liver (Zhang et al., 2017). Many studies have focused on the L. monocytogenes colonization and invasion of the SI, but recent studies have shown that the LI, and not the SI, was the main portal for the dissemination of Lm (Becattini et al., 2017). Our results suggest that microbiota can inhibit Lm growth, especially in the LI, and may provide a target to find functional bacteria to treat Lm infection.
Compared to the SI, the contents of the LI have a more significant inhibitory effect on Lm growth. Several studies, including ours, have demonstrated that the LI has greater species richness and diversity compared to the SI (Booijink et al., 2010). These provide more resistance to colonization and nutritional competition. Furthermore, we found that microbes in feces provide the same resistance as those in the intestine, which confirmed our previous study on treating Lm-infected mice with fecal microbiota transplantation (Guo, 2023).
It has been suggested that the gut microbiota may play an important role in Lm infection and can promote resistance to colonization by pathogenic species (Green and Brown, 2016; Sassone-Corsi and Raffatellu, 2015). Zachar inoculated SPF and Germ-free mice with different doses of Lm by gavage, the results suggested that SPF mice never showed obvious signs of disease with doses as high as 5 × 107 CFU bacteria; in contrast, all mice without germs developed diarrhea and mottled hair and lost weight with the 100 CFU dose of bacteria, and mortality occurred on the fifth postinfection day (Zachar and Savage, 1979). Lactobacilli and Bifidobacteria have been shown to reduce the expansion and spread of orally inoculated Listeria in vivo and prevent Listeria invasion of epithelial cells in vitro (Archambaud et al., 2012).
In addition, the microbiota provides resistance to direct colonization against pathogens. Our results suggest that there were specific bacterial strains that could inhibit Lm growth, and these bacteria were larger than 1 μm in size (Fig. 4). A total of 16S sequencing analysis showed that bacteria can inhibit the growth of Lm existed in Clostridium (Fig. 5). Similar to our results, Becattini assembled a consortium of four Clostridiales species that efficiently protect mice from orally administered Lm (Becattini et al., 2017). There are a large number of commensal species in the gut microbiota to maintain gut health and help the host resist pathogens, which can be developed as a new next generation of probiotics for human health (Satokari, 2019).
Our result also shows that the microbial metabolites filtered by the 0.22 μm micropore filter had no inhibitory effect on Lm growth. However, that was just the result of in vitro experiments, and the role of intestinal metabolites in the host is more complex. Intestinal microbial products can stimulate the host's immune response and indirectly resist pathogenic bacteria (Libertucci and Young, 2019).
Footnotes
Authors' Contributions
L.G.: Writing—reviewing and editing. X.Y.: Data curation and writing—original draft preparation. Q.L.: Conceptualization and methodology.
Declaration of Interest Statement
All persons who meet authorship criteria are listed as authors, and all authors certify that they have participated sufficiently in the work to take public responsibility for the content, including participation in the concept, design, analysis, writing, or revision of the article. Furthermore, each author certifies that this material or similar material has not been and will not be submitted to or published in any other publication.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Doctoral Research Initiation Fund of Zaozhuang University (grant no. 1020722) and the Science and Technology Project of Zaozhuang (grant no. 2021GX07).