
Abstract
The purpose of this study was to reveal the antibacterial activity and mechanism of Polygonatum sibiricum extract (PSE) against Bacillus cereus and further analyze the application of PSE in pasteurized milk (PM). The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values and growth curve analysis were used to evaluate the antibacterial activity of PSE against B. cereus. The changes in contents of intracellular adenosine 5′-triphosphate (ATP) and reactive oxygen species (ROS), activities of β-galactosidase, adenosine triphosphatase (ATPase) and alkaline phosphatase (AKP), cell membrane potential, protein and nucleic acid leakage, and cell morphology were used to reveal the antibacterial mechanism. The effects of PSE on viable count and sensory evaluation of PM during storage were analyzed. The results showed that the MIC and MBC values of PSE against B. cereus were 2 and 4 mg/mL, respectively. Growth curve analysis showed that PSE with a concentration of 2 MIC could completely inhibit the growth of B. cereus. After treatments with PSE, the levels of intracellular ATP and ROS, and activities of β-galactosidase, ATPase and AKP of B. cereus were significantly reduced (p < 0.05). Cell membrane was depolarized, amounts of protein and nucleic acid leakage were significantly increased (p < 0.05), and cell morphology was destroyed. Furthermore, PSE significantly reduced the viable count of B. cereus in PM and improved the sensory quality of PM during storage (p < 0.05). Together, our findings suggested that PSE had the desired effect as a natural preservative applied in PM.
Introduction
B
Pasteurized milk (PM) is one of the common dairy products, and the mild sterilization method allows it to retain most of nutrients and active substances in raw milk (Liem et al., 2016). However, pasteurization cannot completely inactivate B. cereus, so the contamination rate of B. cereus in PM has been high (Chang et al., 2021a; Fei et al., 2019b). Unlike other products, PM is a short shelf life food, and chemical preservatives cannot be added to it, resulting in no ideal strategy to inactivate B. cereus in PM (Kumari and Sarkar, 2016).
Polygonatum sibiricum belongs to the genus Polygonatum and is widely distributed north of the Yangtze River in China (Chen et al., 2021). In general, P. sibiricum refers to the rhizome of P. sibiricum, which is a traditional medicinal herb and widely used as functional food (Liao et al., 2023). The current studies showed that P. sibiricum had the effects of regulating blood sugar, reducing blood lipids, improving immunity and antivirus (Liu et al., 2022; Wan et al., 2023). What's more, Zhao et al. (2019) found that P. sibiricum extract (PSE) contained a variety of ingredients with antibacterial activity and believed that it had the potential as an antimicrobial agent. Meanwhile, P. sibiricum has a sweet taste and a yellowish color, which did not adversely affect the sensory quality of food (Wan et al., 2023). Therefore, PSE should be an ideal natural preservative that can be added to PM.
Therefore, the aim of this study was to reveal the antibacterial activity and mechanism of PSE against B. cereus isolates and to evaluate the bactericidal effect of PSE on B. cereus in PM, thus providing a scientific basis for its application as a natural preservative in real food systems.
Materials and Methods
PSE material and bacteria strains
The PSE was provided by Xi'an Ruihe Bioengineering Technology Co., Ltd. (Xi'an, China). According to the product introduction, PSE was extracted from the rhizome of P. sibiricum and consisted of 60% polysaccharides, 18% steroid saponins, 10% flavonoids, 4% alkaloids, and 8% other substances. The image of PSE is shown in Supplementary Figure S1. The PM was produced by Nanyang Tri-color Pigeon Dairy Co., Ltd. (Nanyang, China) and transported to laboratory with cold chain on the same day, which met the quality requirements of PM in national standard as used in China GB 19645-2010. No pathogenic bacteria and spores were detected in the PM, and the shelf life was 7 d.
A total of six B. cereus strains isolated from PM were used in this experiment, namely, B. cereus YNP3, GZP3, GZP7, YNP11, GXP5, and GXP9, respectively (Chang et al., 2021a). All isolates were used to evaluate the antibacterial activity of PSE, while only B. cereus YNP3 was used to study the antibacterial mechanism and application. B. cereus stock culture were streaked on mannitol-egg yolk polymyxin (MYP) agar and subsequently cultured at 30°C for 24 h. A single typical pink colony was selected and inoculated into brain heart infusion (BHI) broth, followed by an incubation at 30°C for 24 h to obtain B. cereus cultures for future experiments.
Determination of minimum inhibitory concentration and MBC
The minimum inhibitory concentration (MIC) and MBC values of PSE against B. cereus were determined, according to the method reported by Fei et al. (2018), with minor modifications. The PSE treatment concentrations were 16, 8, 4, 2, 1, 0.5, and 0.25 mg/mL, respectively. BHI broth without B. cereus was used as a negative control, while B. cereus culture without PSE was used as a positive control.
Growth curve
B. cereus YNP3 was added to BHI medium containing PSE with 0 MIC (Control), 0.5 MIC, 1 MIC, 1.5 MIC, and 2 MIC, respectively, and the final concentration was adjusted to 107 colony-forming unit (CFU)/mL, then cultured at 30°C for 24 h. Every 2 h, the optical density (OD) of samples at 600 nm was measured using a spectrophotometer (Shanghai Precision Scientific Instrument Co., Ltd., Shanghai, China) to plot the growth curve.
Measurement of intracellular adenosine 5′-triphosphate
The intracellular adenosine 5′-triphosphate (ATP) level was measured as described previously (Sanchez et al., 2010). Briefly, 107 CFU/mL of B. cereus YNP3 suspensions were treated with 0 MIC (Control), 1 MIC, and 2 MIC of PSE at 30°C for 30 min, respectively. Then, the intracellular ATP of sample was determined using an ATP assay kit (Shanghai Beyotime Biotechnology Co., Ltd., Shanghai, China) as described in the instructions.
Determination of intracellular reactive oxygen species
The reactive oxygen species (ROS) was measured based on previous study (Lin et al., 2021). Briefly, B. cereus YNP3 pellets were added to 2,7-dichlorofluorescent diacetate (10 μM), and the concentration was adjusted to ∼107 CFU/mL, followed by incubation at 30°C for 20 min. Then, B. cereus pellets were collected and washed twice with phosphate buffer saline (PBS) and resuspended in PBS. Finally, B. cereus was treated with 0 MIC (Control), 1 MIC, and 2 MIC of PSE for 30 min, respectively, and then their fluorescence intensities were measured using a multifunction microplate reader (Spark, Tecan Austria GmbH, Austria) at excitation wave length of 488 nm and emission wavelength of 525 nm.
Determination of membrane potential
The membrane potential of B. cereus YNP3 was revealed using the method of Guo et al. (2019). Briefly, 107 CFU/mL of B. cereus suspensions were treated with 0 MIC (Control), 1 MIC, and 2 MIC of PSE for 30 min, respectively, followed by the addition of bis-(1, 3-dibutylbarbituriis -(1, 3-dibutylbarbituric acid) trimethine oxonol fluorescent probe into bacterial suspension. After incubation at 30°C for 30 min, the fluorescence intensities of all samples were determined at excitation wavelength of 492 nm and emission wavelength of 515 nm.
Determination of β-galactosidase, adenosine triphosphatase, and alkaline phosphatase activities
The β-galactosidase, adenosine triphosphatase (ATPase), and alkaline phosphatase (AKP) activities of B. cereus YNP3 were determined, according to the report of Lin et al. (2019). First, about 107 CFU/mL of B. cereus were exposed to 0 MIC (Control), 1 MIC and 2 MIC of PSE at 30°C for 3 h. Then, β-galactosidase, ATPase, and AKP in the cells were extracted and measured using their respective assay kits (Shanghai Enzyme-linked Biotechnology Co., Ltd., Shanghai, China) according to the method given in the instructions.
Determination of efflux of protein and nucleic acid
According to the method provided by Chang et al. (2021b), the amount of protein and nucleic acid released from B. cereus YNP3 were determined. Briefly, ∼107 CFU/mL of B. cereus suspensions were treated with 0 MIC (Control), 1 MIC, and 2 MIC of PSE at 30°C for 12 h, respectively. Every 3 h, the treated samples were centrifuged to obtain supernatants. The amounts of protein in supernatants were determined using the Coolmath bright blue method, while the levels of nucleic acid in supernatants were measured by ultraviolet spectrophotometry.
Transmission electron microscopy observations
Transmission electron microscopy (TEM) was performed as described by Li et al. (2016). Briefly, after treatments with 0 MIC (Control), 1 MIC, and 2 MIC of PSE at 30°C for 3 h, the B. cereus YNP3 suspensions (∼107 CFU/mL) were centrifuged to obtain cell pellets. These pellets were fixed in 2.5% of glutaraldehyde for 12 h and then washed three times with PBS. Further, the pellets were fixed in 1% of osmium tetroxide for another 2 h and washed three times with PBS. The samples were dehydrated with different concentrations (50%, 70%, 90%, and 100%) of ethanol for 10 min, then washed with acetone and embedded with Epon Lx-112 (Ladd Research, Williston, VT). Finally, the cells were double stained with uranyl acetate and lead citrate and observed under a TEM (Hitachi, Tokyo, Japan).
Determination of viable number of B. cereus YNP3 in PMPM
B. cereus YNP3 pellets were added to fresh PM containing 0 MIC (Control), 1 MIC, and 2 MIC of PSE, and their concentrations were adjusted to ∼103 CFU/mL. All samples were stored at 4°C for 7 d, and the viable bacterial counts in samples were enumerated on days 0, 1, 3, 5, and 7 (Wang et al., 2022a).
Sensory evaluation
Sensory evaluation of PM was assessed according to the study of Yong et al. (2015). The PSE was added to fresh PM, and the concentrations were adjusted to 0 MIC (Control) and 1 MIC, respectively, then all samples were stored at 4°C for 7 d. On days 0, 3, and 7, the samples were subjected to sensory evaluations by 20 untrained volunteers. The sensory attributes of samples included smell, color, tissue state, taste, and boiling state. The 9-point hedonic scale was used to indicate how much the volunteers liked each sensory attribute. The scales of “1” meant extremely disliked, and “9” meant extremely liked.
Statistical analysis
All experiments were carried out independently three times. Results are expressed as mean ± standard deviation. One-way analysis of variance (ANOVA) in SPSS 20.0 software (SPSS, Inc., Chicago, IL, USA) was used for data analysis. Duncan's multiple range test was used to determine the significant differences (p < 0.05) between treatments.
Results and Discussion
Antibacterial activity of PSE against B. cereus
Table 1 showed that MICs and minimum bactericidal concentrations of PSE against six strains of B. cereus isolated from PM were 2 and 4 mg/mL, respectively. In previous studies, Moringa oleifera seeds (IMOS), olive oil polyphenol extract (OOPE), Zanthoxylum schinifolium essential oil, and Robusta coffee extracts (RCE) against B. cereus were less than or equal to the MIC of PSE (Fei et al., 2019a; Tritripmongkol et al., 2022; Wang et al., 2022a; Zhao et al., 2022). In contrast, the inhibitory effects of chlorogenic acid, trigonostemone, and beetroot extract on B. cereus were weaker than that of PSE (Gong et al., 2023; Sangdee et al., 2021; Tian et al., 2022). It can be seen from above that the antibacterial effect of PSE against B. cereus is above the medium level. Meanwhile, compared with the control group, the growth curves of PSE-treated B. cereus YNP3 were significantly inhibited (p < 0.05) (Fig. 1). When the PSE concentration was 2 MIC, the growth of B. cereus was completely inhibited.
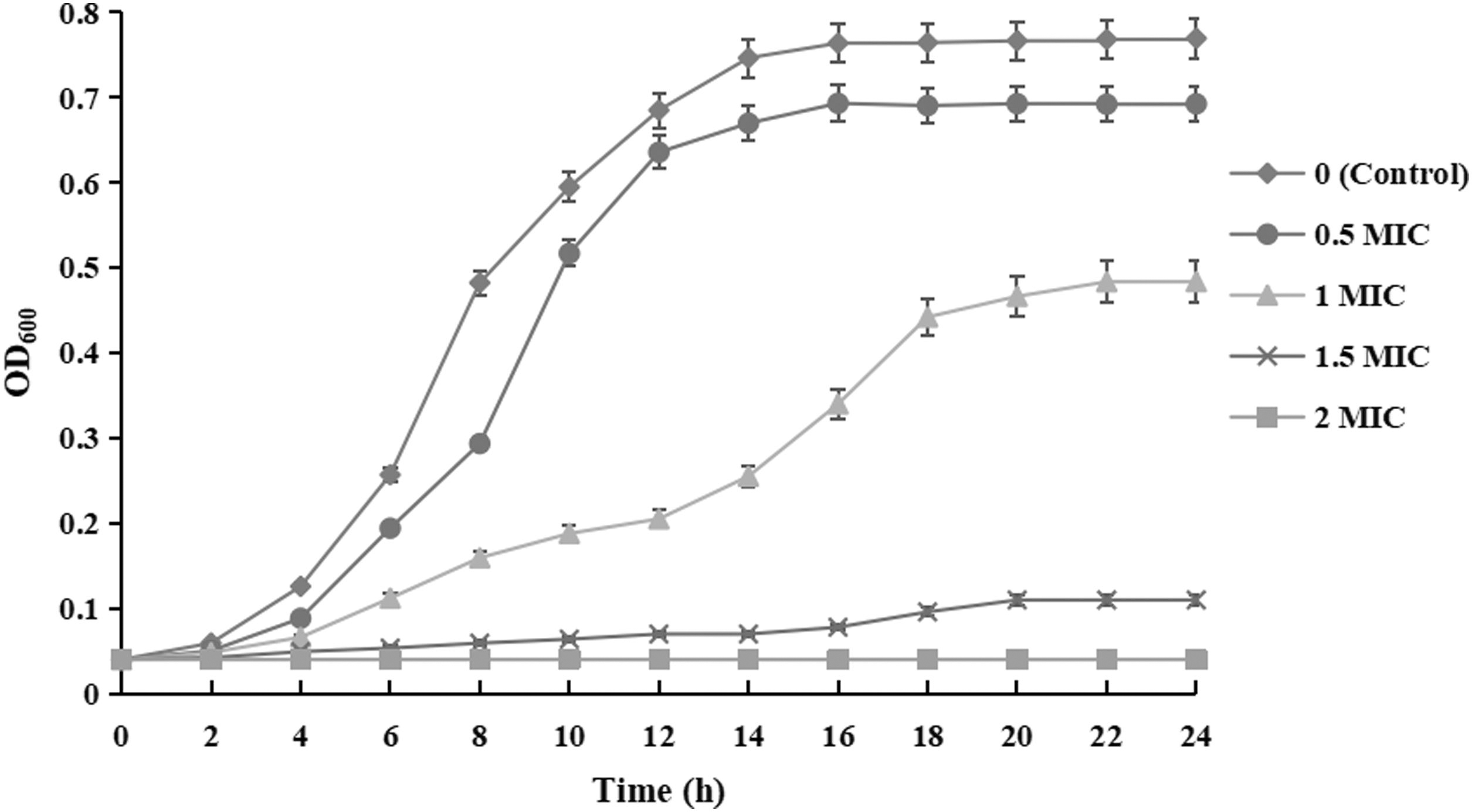
Effect of PSE on the growth curve of Bacillus cereus YNP3. PSE, Polygonatum sibiricum extract.
Minimum Inhibitory Concentration and Minimum Bactericidal Concentration Values of Polygonatum Sibiricum Extract Against Six of Bacillus cereus Strains Isolated from Pasteurized Milk
MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration.
Decreased intracellular ATP and ROS levels
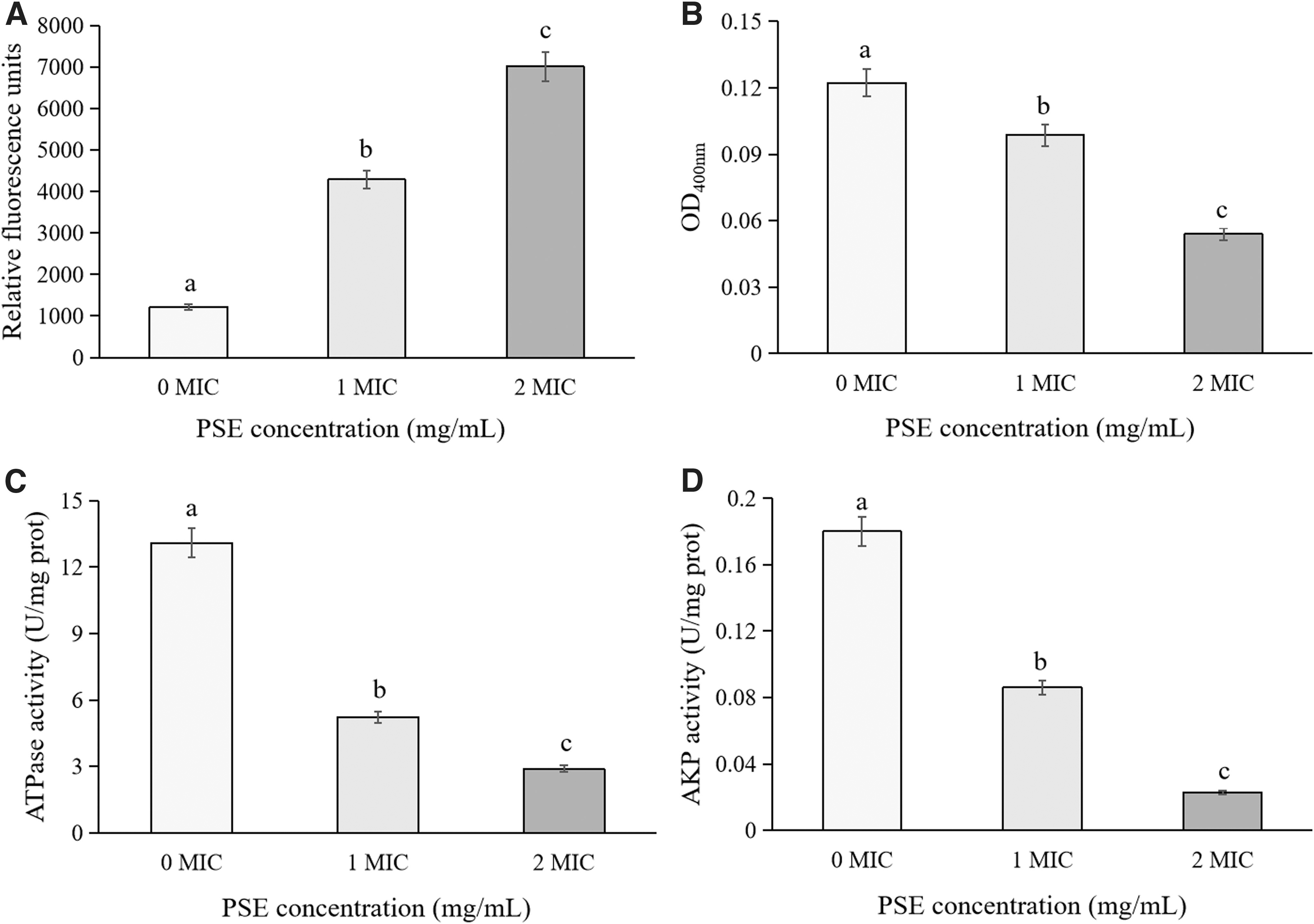
Figure 2A showed that compared with the untreated cells, the intracellular ATP contents of B. cereus YNP3 treated with 1 MIC and 2 MIC of PSE decreased significantly (p < 0.05). Similarly, a significant reduction in intracellular ATP content was found in B. cereus treated with OOPE and IMOE (Fei et al., 2019a; Wang et al., 2022a). Further, previous studies showed that the reasons for the decrease of intracellular ATP content included the loss of ATP pool and proton dynamic components, as well as the leakage of ATP due to the increase of membrane permeability (Fei et al., 2018; Gill and Holley, 2004; Kacprzyk et al., 2007).

Changes in
Figure 2B illustrated that compared with the untreated strains, the intracellular ROS contents of PSE-treated B. cereus were significantly reduced (p < 0.05), and similar results were also found in B. cereus exposed to IMOE and beetroot extract (Gong et al., 2023; Wang et al., 2022a). Moreover, Barzegar and Moosavi-Movahedi (2011) believed that the decrease of intracellular ROS content of bacteria was related to the strong free radical scavenging ability of natural extracts in the polar intracellular environment. In contrast, a significant increase in intracellular ROS was found in Cronobacter sakazakii treated with Houttuynia cordata Thunb. crude extract and Yersinia enterocolitica treated with citral (Chang et al., 2023; Kang et al., 2022). Existing studies showed that some antibacterial substances, especially polyphenols and antibiotics, could increase the content of bacterial intracellular ROS by destroying the ROS defense system and eventually lead to cell apoptosis (Bhattacharya et al., 2018; Grant et al., 2012; Lee et al., 2020).
Damaged cell membrane
Figure 3A showed that compared with the control group, the significantly increased fluorescence intensity of PSE-treated B. cereus YNP3 indicated that PSE can depolarize the cell membrane (p < 0.05). Similarly, B. cereus treated with OOPE, beetroot extract, and thymoquinone also showed membrane depolarization, indicating that the treatments of these substances reduced the amount of negative charge in tested cells (Fei et al., 2019a, 2019b; Gong et al., 2023; Wang et al., 2021). In contrast, thymol caused depolarization of the B. cereus cell membrane, which was related to changes in intracellular pH and leakage of potassium ions (Wang et al., 2022b). Both depolarization and hyperpolarization indicated that the cell membrane potential was far away from the resting potential and the cell membrane has been damaged (Chen et al., 2023).
Effects of PSE on the
Further, β-galactosidase has been reported to be involved in cell membrane permeability, and ATPase is an important ion transporter in the cell membrane (Liang et al., 2020; Song et al., 2016). AKP existed between cell membrane and cell wall, and the decrease of AKP activity could indirectly reflect the increase of cell membrane permeability of tested bacteria (Wang et al., 2020). In this study, a significant decrease in activities of β-galactosidase, ATPase, and AKP was found in the cell membrane of B. cereus treated by PSE compared with that of untreated cells (p < 0.05) (Fig. 3B–D). These results were consistent with previous reports (Diao et al., 2018; Wang et al., 2022a), suggesting that PSE seriously damaged the cell membrane of B. cereus.
Leakage of protein and nucleic acids
Figure 4A showed that compared with the control group, protein leakages of both 1 MIC and 2 MIC treated B. cereus YNP3 were significantly increased throughout the treatment period (p < 0.05). Meanwhile, Figure 4B illustrated that the amounts of nucleic acid leakage of PSE-treated B. cereus were significantly higher than that of untreated strains, and the amount of leakage also increases significantly with increasing PSE concentration (p < 0.05). Similarly, previous studies have also found a large amount of protein and nucleic acid leakage in B. cereus treated with IMOE, C. sakazakii treated with Chrysanthemum buds crude extract, and Staphylococcus aureus treated with Rosa roxburghii Tratt pomace crude extract, and suggested that this phenomenon was related to the damage at cell membrane level of tested bacteria under the action of natural extract (Chang et al., 2021b; Chen et al., 2023; Wang et al., 2022a).
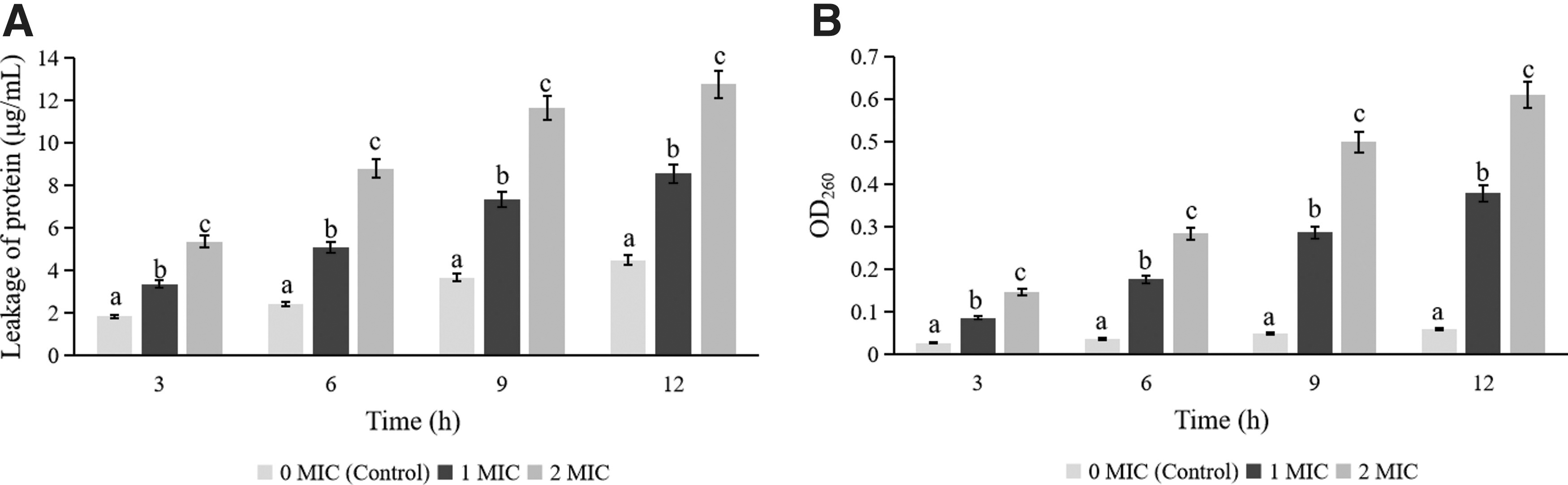
The amount of
Changes in cell morphology
As shown in Figure 5, compared to the untreated cells (Fig. 5A), B. cereus treated with 1 MIC of PSE showed a significant reduction in cell fluid and a slight depression (Fig. 5B). When the action concentration of PSE was 2 MIC, the cell fluid of tested bacterial cells was further reduced, and the cell depression phenomenon was more serious, resulting in cell deformation (Fig. 5C). Fei et al. (2019a, 2019b; 2020) found a similar phenomenon in B. cereus treated with OOPE and C. sakazakii treated with Amaranthus tricolor crude extract and suggested that the decrease of cell fluid was caused by the leakage of cell contents including ATP, proteins, and DNA after improvement of membrane permeability, which was also consistent with the findings in this study (Figs. 2A and 4). However, not all natural extracts cause cell fluid leakage from tested cells, for example, C. sakazakii and Y. enterocolitica did not experience significant cell fluid leakage after treatment with citral (Kang et al., 2022; Shi et al., 2016a).

TEM images of Bacillus cereus YNP3
The application of PSE in PM
Table 2 showed that during the 7-d storage period, the viable counts of B. cereus in PM without PSE increased from 3.12 to 4.02 Log CFU/mL. Compared with the control group, the viable counts of B. cereus in PM treated with 1 MIC and 2 MIC of PSE decreased significantly (p < 0.05), and after 7 d of storage, the viable bacterial counts of both treated groups were reduced from 3.09 to 2.28 Log CFU/mL and from 3.15 to 1.06 Log CFU/mL, respectively. Similarly, IMOS has been reported to reduce the levels of B. cereus and C. sakazakii in goat milk (Wang et al., 2022a), and Larimichthys crocea whey acidic protein-derived peptide has been found to effectively inhibit S. aureus in milk (Yang et al., 2020).
Viable Bacterial Counts of Bacillus cereus YNP3 (Log Colony-Forming Unit/mL) Exposed to Pasteurized Milk with Polygonatum Sibiricum Extract Concentrations of 0 Minimum Inhibitory Concentration (MIC), 1 MIC, and 2 MIC at 4°C for 0, 1, 3, 5, and 7 D, Respectively
All values were represented by mean ± standard deviation (n = 3), and different lowercase letters in each row indicated significant differences between different treatment groups (p < 0.05).
MIC, minimum inhibitory concentration.
Figure 6 showed that the scores of smell, color, tissue state, taste, and boiling state of PM in the MIC group were significantly higher than those in the control group (p < 0.05). Previous studies have shown that protease and lipase produced by some microorganisms will decompose the protein and fat in milk, reducing the nutritional value and flavor of milk (Billeaud, 2021; Nemeckova et al., 2009). Vargas et al. (2019) found that excessive bacteria in milk increased the acidity of milk, thus reducing the stability of milk system. According to the above viewpoints, the role of PSE in maintaining sensory quality of PM should be related to its strong antibacterial ability. In addition, due to the small addition amount (0.4 g/bag), adding PSE will only increase the cost of PM per bag by 0.13 yuan, which is acceptable to consumers.
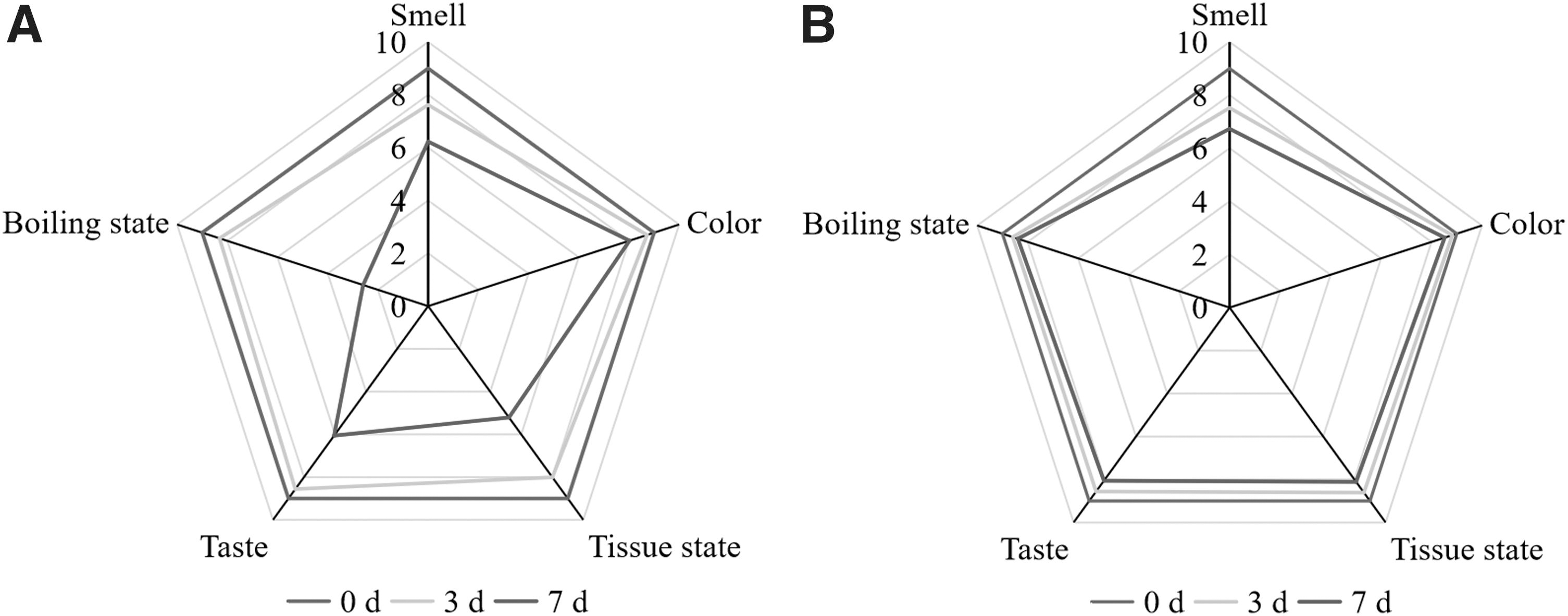
The effect of PSE sensory evaluation of PM during storage.
Conclusion
In summary, PSE can effectively inhibit B. cereus in PM. However, considering the physiological function and sweet taste of PSE, the necessary animal experiments and dose optimization should be carried out before PSE can be applied to the industrial production of PM.
Footnotes
Authors' Contributions
P.F. and Y.C.: conceptualization, methodology, formal analysis, software, funding acquisition, supervision, and writing—original draft; Z.S., X.L., P.J., Y.M., G.D., C.W., M.B., and Y.L.: methodology, formal analysis, and software; H.F. and X.C.: conceptualization, data curation, and supervision.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the doctoral scientific research launch fund project of Nanyang Institute of Technology (NGBJ-2023-08), Interdisciplinary Sciences Project, Nanyang Institute of Technology (520079), Guizhou Provincial Key Technology R&D Program [2023] (No. 464). and Natural Science Foundation of Guizhou Province (Qiankehe foundation-ZK [2022] General 008).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.