
Abstract
Clostridioides difficile and its endospores possess the characteristics of a foodborne pathogen and have been detected at several stages in the food chain. In the presence of an imbalance in host intestinal ecology, C. difficile can proliferate and cause intestinal infections. Multiple food source factors can substantially alter the host’s gut ecosystem, including the consumption of baijiu. However, it remains to be known whether the gut ecological changes induced by the consumption of baijiu increase the risk of C. difficile invasion and infection. In this study, C. difficile cells were exposed to two commercially available baijiu to evaluate the effect of baijiu on C. difficile cells and to verify through a mouse model. The results showed that baijiu effectively inhibited the growth and biofilm production of C. difficile, downregulated the expression levels of tcdA and tcdB virulence genes but upregulated the expression level of spore-producing genes Spo0A, enhanced the spore production, as well as increased C. difficile cell adhesion to Caco-2 cells. The mouse model showed that the intake of baijiu promoted the invasion and infection of C. difficile spores, causing damage to the cecum tissue, accompanied by an increase in the gut lipid carrier protein-2 (Lcn-2) and TcdA toxin protein levels. Simultaneously, cholic acid was elevated, whereas deoxycholic acid was decreased. This study is the first to find a possible link between baijiu intake and C. difficile spore invasion and infection.
Introduction
Clostridioides difficile is a pathogenic bacterium commonly found in the human gut, capable of producing two toxin proteins with synergistic pathogenic effects, destroying intestinal epithelial cells and thus inducing intestinal infections and tissue damage, with clinical symptoms ranging from mild to moderate diarrhea, severe diarrhea, pseudomembranous colitis, toxic megacolon, and eventually, death (Smits et al., 2016). The main cause of C. difficile infection (CDI) stems from its highly environmentally resistant endospores, which can remain dormant for long periods under disturbed gut ecology (e.g., antibiotic therapy) or specific dietary interventions, and these spores germinate under suitable nutritional conditions to form virulent strains, resulting in recurrent infections (Hazleton et al., 2022, Pruss et al., 2022, Yakabe et al., 2022, Zeng et al., 2022). Previously, CDI outbreaks were reported in antibiotic abusers, long-term hospitalized populations, and immunocompromised middle-aged and elderly populations, and C. difficile was classified as a clinical pathogenic microbe (Gonzales-Luna et al., 2023). Notably, the recent detection of C. difficile in numerous foods with the same ribotypes (RTs) associated with human infections, coupled with the increasing rate of community-acquired infections, suggests that C. difficile also possesses the characteristics of a foodborne pathogen (Candel-Pérez et al., 2019, Tkalec et al., 2019, Bolton and Marcos 2023, Marcos et al., 2023). Tkalec et al. (2019) collected 12 home-grown vegetables in Slovenia and found a C. difficile detection rate of 18.2% in vegetables, with the highest rate in potatoes (28%), followed by ginger (6.7%) and leafy vegetables (9.4%). Upward of 40 different RTs of C. difficile, including 6 highly virulent strains, were present in vegetables, shellfish, and pork, whereas 9 RTs (002, 003, 012, 014, 027, 029, 070, 078, and 126) have been isolated from patients with confirmed community-associated CDI (Bolton and Marcos, 2023). C. difficile-associated food contamination is mainly from its spores, which originates from sewage treatment plants, shellfish in seafood, horse and pig manure, organic fertilizer from traditional crops, slaughterhouses, and retail meat products (mainly beef and pork as well as poultry) (Candel-Pérez et al., 2019). Managing or reducing the rate of C. difficile spore infections is difficult because these spore transmission routes involve multiple links in the food chain.
The association between diet and CDI was not fully understood. Although C. difficile spores are present in the food chain, there is no published evidence that they are capable of causing infections in humans. In fact, the normal gut ecology of the human body can effectively act against the invasion of C. difficile and the germination of its spores (Yang and Yang, 2019). However, once this homeostasis was disturbed, it was likely to provide an opportunity for the invasion of C. difficile spores. Some studies have shown that baijiu has substantial effects on gut ecology, mainly in the form of altered microbiome and metabolome (Fang et al., 2019). Baijiu is a world-famous distilled spirit, which occupies a dominant position in China’s liquor market. Its main components are ethanol (EtOH) and water (98–99% of the total), in addition to a large variety of trace organic compounds such as acids, esters, alcohols, and aldehydes (1–2% of the total) (Liu and Sun, 2018). It is unclear whether this change induced by baijiu creates an opportunity for invasion and infection by conditionally pathogenic microbes, such as C. difficile. We were considering that C. difficile and its endospores are already present in several links of the food chain. Therefore, we hypothesized that the gut disturbance caused by baijiu may increase the risk of such foodborne pathogenic bacteria infecting the host gut. In this study, we used two different commercially available baijiu (both with 40.8%) to treat C. difficile strains, to investigate the effects of baijiu on the biological properties of C. difficile and to verify the association between baijiu and C. difficile spore germination and infection through in vitro and in vivo experiments, to preliminarily assess the potential risks involved.
Materials and Methods
Baijiu, bacterial strains, cells
Two brands of commercially available baijiu, named sample 1 (S1) and sample 2 (S2), all with an alcohol concentration of 40.8%, were purchased from the local market, whereas the same concentration of EtOH was used as a control. All baijiu and alcohol groups were filtered through 0.22 μm sterile filters before the start of the experiment. C. difficile ATCC 43255 and Caco-2 cells were purchased from the American Type Culture Collection (ATCC) (Manassas, VA, USA). C. difficile was incubated in the brain–heart infusion (BHI) broth (hopebio Ltd., Qingdao, China) at 37°C in an anaerobic workstation (85% nitrogen, 10% hydrogen, 5% carbon dioxide; Electrotek AW 400TG Two Gas Version, United Kingdom). C. difficile moxalactam norfloxacin agar (CDMN) selective medium (Oxoid Ltd., Basingstoke, UK) was used for live C. difficile enumeration. Caco-2 cells were cultured using Dulbecco’s modified Eagle’s medium (DMEM) (KeyGEN Biotech Ltd., Nanjing, China) with 20% fetal bovine serum (Gibco, Thermo Fisher Scientific, USA) and 1% antibiotics (penicillin, streptomycin 100×).
Effect of baijiu on the growth of C. difficile and determination of minimum inhibitory concentration
Regarding the method of Yang and Yang (2018), C. difficile was incubated in fresh BHI broth until the end of the log phase (OD590 approximately 1.2), from which 20 µL (containing 3 × 104 cells) was added to a 96-well plate, and then baijiu (40.8%) or EtOH (40.8%) in each well (twofold dilution method with fresh BHI broth) and adjusted to a final volume of 200 µL with fresh BHI broth, resulting in a final concentration of baijiu or EtOH of the specified value, respectively. The 96-well plates were subsequently incubated at 37°C, anaerobically for 24 h, and then placed in a microplate reader (PE & EnSpire 2300) to determine the OD590 values, which determined the inhibitory effect of baijiu and EtOH on C. difficile and the minimum inhibitory concentration (MIC). Normal growth of C. difficile (without baijiu or EtOH) was used as a control.
Effect of baijiu on the biofilm production of C. difficile
The bacterial solution from the previous step was further incubated for 48 h. Each well was removed and washed twice with sterile PBS (pH 7.4, Sangon Biotech, Shanghai) and dried at 37°C for 15 min. After that, 200 µL of crystalline violet solution (0.2%) was added to each well for staining and placed at 37°C for 30 min. The wells were then washed twice with sterile PBS to remove the crystalline violet solution and dried at 37°C for 15 min. Add 200 µL of EtOH–acetone (80%/20%) solution and measure OD570. Fresh BHI broth is used as a decolorization group (to remove background interference). Normal growth of C. difficile (without baijiu or EtOH) was used as a control.
Effect of baijiu on the expression levels of tcdA, tcdB, and Spo0A in C. difficile
With some modifications based on the method of Yang and Yang (2018), after centrifugation (6000 rpm) of the C. difficile bacterial solution (50 mL, 3 × 105 colony-forming unit [CFU]/mL), the cell pellets were collected and resuspended in fresh BHI broth in quintuplicate with 10 mL of S1, S2, EtOH, and BHI, respectively, after which all samples were incubated in an anaerobic workstation at 37°C for 24 h and then centrifuged to collect the cell pellets. The C. difficile RNA was extracted using the RNA-prep Pure Kit for bacteria (TianGen Biotech, Beijing, China) according to the manufacturer’s instructions. Contaminating genomic DNA was removed with DNase (DNA-free kit; TianGen Biotech, Beijing, China), and the final total RNA and purity were detected by UV absorption and agarose gel electrophoresis, respectively. The gene expression levels of tcdA, tcdB, and Spo0A were detected by real-time quantitative polymerase chain reaction (qPCR) with specific primers, referring to the method of Aldape et al. (2015). One microgram of cDNA was synthesized from the total RNA using the All-in-One First-Strand cDNA Synthesis SuperMix (TransGen Biotech, Beijing, China) and diluted 1:20 with sterile water, amplified using TransStart Top Green qPCR SuperMix (TransGen Biotech) in a Mastercycler ep realplex system (Eppendorf, Hamburg, Germany). C. difficile 16S rRNA was used as an internal reference, and Top Green qPCR SuperMix (TransGen Biotech) was used with the following reaction conditions: 30 s at 94°C, followed by 40 cycles (including 5 s at 94°C, 15 s at 55°C, and 10 s at 72°C with annealing). The relative expression of the genes was determined using the 2−ΔΔCt method (Aldape et al., 2015). The mean Ct of 16S rRNA (internal reference) was subtracted from the mean Ct of samples for tcdA, tcdB, and Spo0A genes (ΔCt). The ΔCt of the untreated control at 6 h was subtracted from the mean ΔCt of each experimental group (ΔΔCt).
Effect of baijiu on the spore production of C. difficile
With some modifications based on the methods of Aldape et al. (2015), 20 mL of fresh C. difficile bacterial solution (5 × 108 CFU/mL) was mixed with 20 mL of S1, S2, EtOH, and sterile water and incubated overnight at 37°C in an anaerobic workstation and centrifuged to collect the cell pellets, washed twice with PBS, and adjusted OD590 to 0.8. Ten milliliters of samples (3 × 105 CFU/mL) under each treatment were mixed with 10 mL of EtOH (100%) at room temperature and shaken (3000 rpm for 1 h) to kill vegetative cells; after that, samples were centrifuged at 12,000 rpm for 5 min, and the cell pellets were collected and washed twice with PBS. In the last wash, the pellets were resuspended in PBS, and spore numbers were obtained by the gradient dilution method after coating the BHI plates and incubating them in an anaerobic workstation at 37°C for 72 h.
The cytotoxicity effect of baijiu on the Caco-2 cells
After Caco-2 cells grew to 80% in 12-well plates, they were washed twice with PBS, digested with trypsin, centrifuged at 1000 rpm for 4 min, and resuspended in 1 mL of DMEM broth. Cells were counted by a cell counter and diluted to 200,000 cells/mL with DMEM. One milliliter of cell solution (200,000 cells) was taken per well for plate spreading and incubated at 37°C with 5% CO2 for 72 h. After a twofold dilution of baijiu and EtOH with DMEM broth, 100 μL of the diluted baijiu samples was added to each well, and the positive control (PC) group was replaced with 100 μL of DMEM. After incubation for 4 h at 37°C and 5% CO2, the supernatant of each well was discarded, and 100 μL of Cell Counting Kit-8 (MedChemExpress, New Jersey, USA) solution diluted 10 times with DMEM was added to each well, incubated for 30 min at 37°C and 5% CO2, and OD450 was measured in a microplate reader.
Adhesion test of C. difficile treated with baijiu to the Caco-2 cells
Twenty milliliters of baijiu was filtered through a sterile 0.22 μm membrane and mixed with 20 mL of C. difficile bacterial solution (3 × 105 CFU/mL), and the supernatant was removed by centrifugation after 12 h of anaerobic incubation at 37°C. After that, the cell pellets were resuspended with DMEM, and the OD590 under each treatment was adjusted to a similar value (OD590 = 0.2). When Caco-2 cells grew to a density of 50%, the DMEM broth was gently discarded and washed three times with 1 mL of PBS. Add 500 μL of C. difficile bacterial solution (3 × 105 CFU/mL) and 500 μL of DMEM without antibiotics to each well and set up three replicate wells for each group. The plate was coated with the bacterial solution at 0 h and served as a control group. The above bacterial solution was mixed with the cells and incubated at 37°C under anaerobic conditions for 1 h. The supernatant was discarded after 1 h. The unadhered bacteria were removed by washing one time slowly with PBS. Add 200 μL of trypsin under digestion for 3–5 min, then add 600 μL of DMEM without antibiotics, blow to collect cells and bacteria at 6000 rpm, centrifuge for 3 min, discard the supernatant, and resuspend the precipitate with 1 mL of BHI for dilution smear plate counting. The preadherent bacteria (the very first 1 mL of OD590 = 0.2 bacterial solution) was used as a control. Adherence rate (%) = number of C. difficile after adherence/number of C. difficile before adherence × 100%.
Animal experiment
A total of 55 specific-pathogen-free (SPF) C57BL/6 male mice (7 weeks old) were purchased from Shanghai SLAC Laboratory Animal, Co., Ltd. (Shanghai, China) and allowed to acclimatize for one week before modeling. Mice were housed in individual cages under identical conditions (21°C, fed a standard chow), and all mice were identified as negative for C. difficile through a C. difficile TOXA/B IITM kit (Tech Lab, Kraft Drive Blacksburg, VA, USA) before the start of the experiment. A suspension of C. difficile spores (5 × 106 CFU/mL) was prepared in advance of infection according to the method of Bublitz et al. (2023). A total of five groups were set up. Among them, the NC group (n = 5) was grown normally without any treatment; the C. difficile group (n = 10) was without any treatment except for the challenge of C. difficile spores; the S1 group (n = 10), S2 group (n = 10), and EtOH group (n = 10) were gavaged with baijiu or EtOH (50 μL) and subsequently challenged with C. difficile spores (200 μL) at the designated time points. Mice were monitored for survival and body weight throughout the experiment and euthanized in the near-death state or if they lost more than 20% of their body weight from baseline. All experimental procedures involving mice were approved by the Animal Ethics and Welfare Committee of Nanjing Normal University (IACUC-20230253). During the experiment, once the mice appeared to die, their cecum tissues were immediately collected and preserved in a 10% formalin solution. In addition, mouse fecal samples were collected at each of the four designated time points and stored in sterile tubes at −80°C. The fecal collection time points were as follows: the day before baijiu gavage at the end of the acclimation period (time 1, T1), the day before infection with C. difficile spores (time 2, T2), the third day after infection with C. difficile spores (time 3, T3), and the sixth day after infection with C. difficile spores (time 4, T4). At the end of the last sampling point (T4), after one week of continued observation, the remaining mice were euthanized, and their cecum tissue was collected. The fecal samples were collected and were immediately counted for C. difficile viable bacteria using the CDMN plate (Oxoid, Basingstoke, Hants, UK). All these plates were incubated in the anaerobic workstation at 37°C for 5 days. Meanwhile, the fecal samples were immediately measured for TcdA toxin, lipid carrier protein-2 (Lcn-2), cholic acid (CA), and deoxycholic acid (DCA) levels after being properly stored and brought back to the laboratory.
Histological analyses
Mouse cecum tissues were preserved in 10% formalin solution and fixed at 4°C for 24 h. Afterward they were paraffin-fixed and cut into 5 µm sections with a microtome (Leica EM UC7, Leica, Munich, Germany). Finally, all samples were stained with hematoxylin-eosin (H&E) and then observed under a light microscope (Olympus, Mod. U-LH100HG).
Determination of fecal Lcn-2 in feces by ELISA
One gram of stool sample was resuspended in PBS containing 0.1% Tween 20 to a final concentration of 100 mg/mL and vortexed for 20 min to obtain a homogeneous stool suspension. Centrifuge at 14,000 × g and 4°C for 10 min, collect the supernatant, and store at −20°C until analysis. The content of Lcn-2 in the supernatant was determined using the Duoset Mouse Lcn-2-ELISA kit (R&D Systems, Minneapolis, Minnesota, USA) using the colorimetric peroxidase substrate tetramethylbenzidine and OD450 was measured.
Determination of the C. difficile spores and toxin level and bile acid content in feces
Stool samples were collected, homogenized, and resuspended in PBS in duplicate. One copy was diluted in gradient and coated on CDMN for spore counting, whereas the TcdA toxin content in the stool was detected using the Mouse CDT-A ELISA Kit (RenjieBio, Shanghai, China). The other was centrifuged, and the supernatant was collected, diluted, and measured for CA and DCA levels using the Mouse CA/DCA ELISA Kit (Enzyme-linked Biotechnology, Shanghai, China).
Statistical analysis
All the data obtained in this study were presented as mean ± standard deviation. Statistical analysis was performed using analysis of variance using Minitab Statistical Software (version 20) (Minitab Inc., State College, PA, USA). A p value of <0.05 was considered statistically significant.
Results and Discussion
Effect of baijiu on the growth and biofilm production of C. difficile
The original concentration of each baijiu was used as a starting point for serial dilutions, and subsequently, C. difficile cells were treated with the different concentrations of baijiu obtained to observe changes in growth and biofilm production. S1, S2, and EtOH gradually weakened the inhibitory effect on the growth of C. difficile with increasing dilution (Fig. 1). S1 antagonized C. difficile at an MIC of 40.8%, S2 at 2.55%, and EtOH at 10.2%. Meanwhile, there was no significant correlation between the variation of C. difficile biofilm yield and baijiu concentration, where the MIC of S1 antagonizing C. difficile biofilm was 0.1594%, and 0.0797% for S2 and EtOH. Actually, baijiu contains alcohol as the main ingredient that is capable of killing bacteria. Kang et al. (2022) found that baijiu showed a strong antibacterial activity against Escherichia coli and Staphylococcus aureus and inactivated their mature biofilm, which was similar to the results that emerged in the present study, indicating baijiu against the growth and biofilm formation of some Gram-positive bacteria, such as C. difficile. However, it is essential to note that harsh survival conditions (e.g., alcohol) can force C. difficile to produce spores, setting the stage for recurrent infections (Smits et al., 2016).

Effect of baijiu on the growth and biofilm production of Clostridioides difficile. Tukey test for multiple comparisons was used between treatments of different concentrations in the same group, and different letters represent significant differences, p < 0.05. Control represents normal growth of C. difficile without any treatment. OD590nm for growth and OD570nm for biofilm.
Effect of baijiu on the expression levels of tcdA, tcdB, Spo0A, and spore production of C. difficile
Baijiu and EtOH downregulated the expression levels of tcdA and tcdB but increased the expression level of Spo0A in C. difficile cells, with the most prominent effect under S2 treatment, followed by EtOH and S1. Whereas, a viable bacterial count (evolved from spores) further indicated that the numbers of C. difficile spores in the baijiu and EtOH groups remained at high levels (Fig. 2). It was found that some natural active products and probiotic fermentation products can also inhibit the expression of these key genes (Yang and Yang, 2019). Notably, the expression levels of tcdA, tcdB, and Spo0A were uniformly downregulated in the vast majority of studies that inhibited C. difficile (Yang and Yang, 2019). The expression levels of the tcdA and tcdB genes in C. difficile directly control the production of the toxin proteins TcdA and TcdB, which can synergistically cause infection and destruction of intestinal epithelial cells, whereas the expression level of Spo0A is directly correlated with biofilm production and spore germination in C. difficile (Yang and Yang, 2019). In contrast, here we found that the expression level of Spo0A was opposite to that of tcdA and tcdB, suggesting that baijiu and EtOH differ slightly in repressing the expression of key genes. Alcoholic substances did not appear to have a pronounced inhibitory effect on the spore-associated gene. Strong et al. treated C. difficile spores with 70% EtOH for 20 min and found that the spore survival rate remained as high as 90% (Strong et al., 2014). Alcohol-related hand sanitizers and antiseptics widely used in health care centers and hospitals are not effective in removing spores produced by pathogens such as C. difficile and Bacillus anthracis (Nerandzic et al., 2015). This also makes alcohol-based fungicides ineffective in preventing the recurrent infections caused by C. difficile. Alcohol-related foods can substantially alter host gut ecology, including changes in the gut microbiome and metabolome, and such changes in gut ecology may present opportunities for germination, colonization, and infection of spore-producing microbes. From this point of view, C. difficile spores in the food chain may have a high invasion rate and infection rate for the baijiu-drinking population.
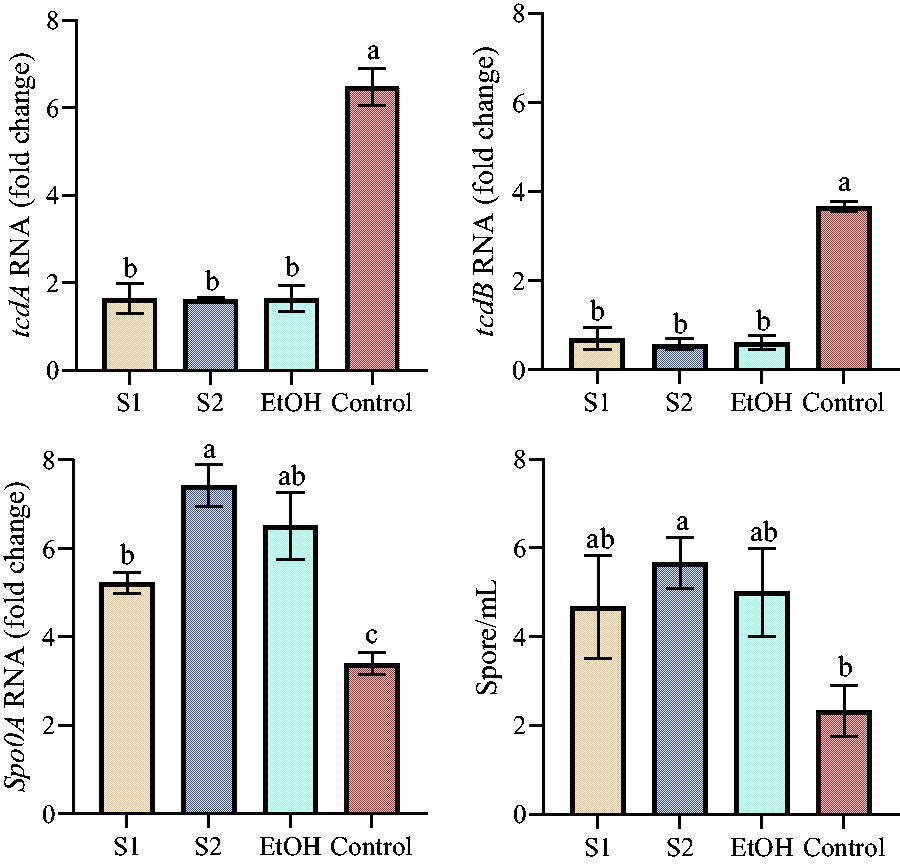
Effect of baijiu on the regulation of tcdA/B and Spo0A expression levels of Clostridioides difficile and spore production. Tukey test for multiple comparisons was used between different groups, and different letters represent significant differences, p < 0.05. Control represents normal growth of C. difficile without any treatment.
The cytotoxicity effect of baijiu on the Caco-2 cells
The cytotoxicity effect of baijiu and EtOH on the Caco-2 cells gradually decreases with increasing dilutions (except for the 10.2% concentration) (Fig. 3). Notably, the cytotoxicity effect of S1, S2, and EtOH reached 100% for Caco-2 cells at 10.2%, and after exceeding this concentration, the killing power decreased substantially, and the survival rate of Caco-2 cells was close to 90%. Wang et al. (2014) treated Caco-2 cells with different concentrations (1%, 2.5%, 5%, 7.5%, and 10%) of alcohol for 3 h and found that Caco-2 cell viability was not altered at alcohol concentrations below 5%. In the concentration range below 10%, alcohol does not increase the release of cytoplasmic enzyme lactate dehydrogenase (Wang et al., 2014); at the same time, alcohol concentrations above 5% can cause cell shedding, and at 10% EtOH concentration, the cell shedding rate exceeds 50% (Wang et al., 2014). Miller et al. (2000) found that 10% EtOH caused microtubule breakage and reduced cell survival in Caco-2 cells, and the lowest EtOH concentration that caused this microtubule disruption was 2.5%. Asai et al. (2003) also found that low concentrations of EtOH (10%) induced apoptosis in Caco-2 cells. Banan et al. (2007) found that EtOH destabilizes the Caco-2 cytoskeleton and disrupts its barrier integrity by activating the NF-κB pathway, resulting in a significant reduction in the proportion of cells with a normal actin backbone. Taken together, baijiu at concentrations below 10% produced less effect on Caco-2 cells.
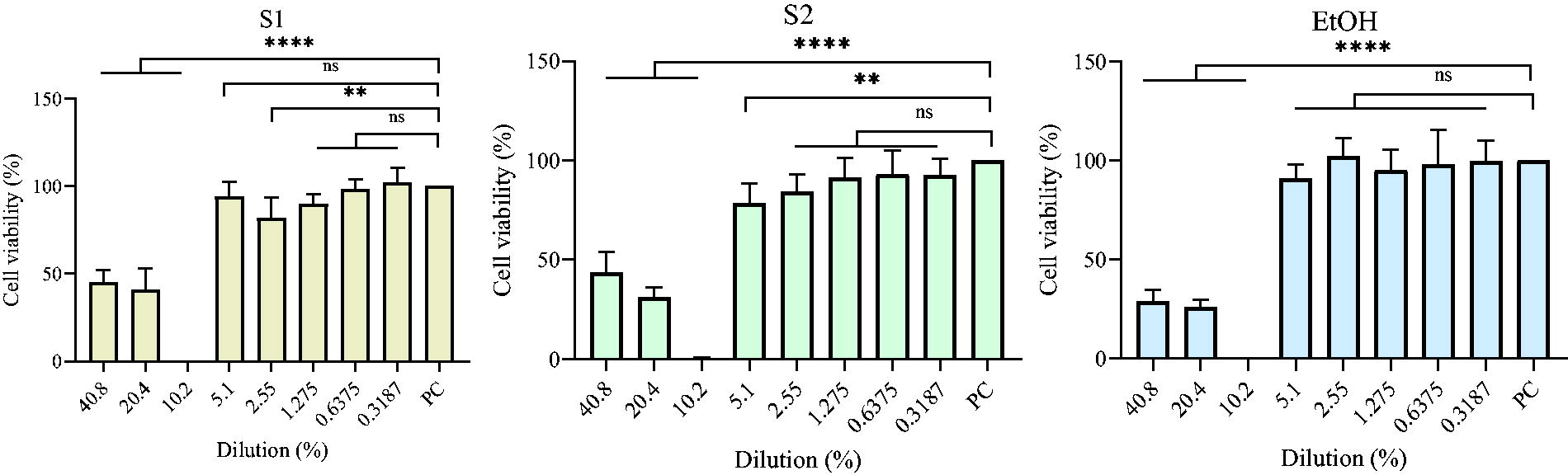
The cytotoxicity effect of baijiu on Caco-2 cells. Ordinary one-way ANOVA was used between treatments of different concentrations in the same group. ns, no significant difference; **p < 0.01; ****p < 0.0001. PC represents positive control, that is, normally growing Caco-2 cells. ANOVA, analysis of variance.
Adhesion of C. difficile to the Caco-2 cells after baijiu treatment
Compared with the normal growth of C. difficile, the ability to adhere to the Caco-2 cells was increased in C. difficile treated with baijiu and EtOH (Fig. 4). This similar phenomenon was also found in some kinds of honey, such as Manuka honey, pine honey, and pollen honey (bee bread), at subinhibitory concentrations, increasing the adhesion of C. difficile to HT-29 and CCD 841 cell lines (Wultańska et al., 2022). C. difficile adhesion is essential for its colonization of the host intestine and a prerequisite for further proliferation, infection, and toxin secretion (Anwar and Vedantam, 2022). It has been shown that the expression level of the Spo0A gene is closely related to the colonization ability of C. difficile (Baloh and Sorg, 2021, Lee et al., 2021). Based on the results obtained earlier in this study, we speculated that the elevated expression level of Spo0A in response to baijiu is likely to be one of the reasons for the promotion of C. difficile adhesion. It was noteworthy that the adhesion rates of S1 and S2 treatment groups were dramatically higher than that of EtOH, implying that there may be other trace substances in baijiu that can promote adhesion. Nevertheless, alcohol in baijiu remained the main contributor to this phenomenon. Taken together, baijiu S1 and S2 increased C. difficile adhesion to the Caco-2 cells, which may, in turn, promote C. difficile colonization in the host gut and set the stage for further infection.

Adhesion of Clostridioides difficile organisms to Caco-2 cells. Tukey test for multiple comparisons was used between different groups, and different letters represent significant differences, p < 0.05. Control represents normal growth of C. difficile without any treatment.
Survival rate and histopathological analysis of mice
The baijiu and EtOH groups did not cause mortality in mice, and death occurred mainly after infection with C. difficile spores. In the experiment, significant decreases in the relative body weight of mice occurred mainly in the first and second gavage of baijiu and after infection with C. difficile spores. After being challenged with C. difficile spores, mortality occurred in the S1, S2, EtOH, and C. difficile groups, in addition to the NC group. The final survival rate of mice was, in descending order, NC (100%) > C. difficile (90%) > S1 (70%) > S2 (60%) = EtOH (60%) (Fig. 5B). At the end of the acclimation period, all mice were gavaged with baijiu for the first time, and the body weight of mice in S1, S2, and EtOH groups started to decrease, but after three days, the body weight rebounded to the level before gavage, after that, the second gavage with baijiu was performed, and the body weight of mice decreased again, but rebounded after three days (Fig. 5C). Although mice in the S1, S2 and EtOH groups were able to return to their predrinking levels in terms of body weight, the changes induced by baijiu increased the germination and susceptibility of C. difficile spores, which eventually caused severe infection and increased mortality. Compared with the C. difficile group, which showed one death only after five days of spore infection, this indicates that the normal intestinal ecology untreated with baijiu or alcohol can provide good resistance to the invasion and infection of C. difficile. The normal gut microbiota structure is always in a dynamic balance, that is, harmful microbes, beneficial microbes, and commensal microbes are in mutual control (Yang et al., 2022). Beneficial microbes counteract the growth of harmful members by competing for nutrients, occupying ecological sites, and secreting antimicrobial-like substances. In addition, changes in intestinal nutrients, especially bile acid metabolites, can markedly affect the germination and colonization of C. difficile spores and infection (Yang and Yang, 2019). Significantly higher rates of CDI in people with gut ecological disruption or low gut immunity are caused by treatment with antibiotics or other drugs (Yang and Yang, 2019). The presence of C. difficile spores in the food chain does not generally cause host infection, which may be attributed to the normal ecological environment in the host’s gut that can effectively contain the invasion of C. difficile spores. Although the alcohol contained in baijiu can inhibit the growth of several harmful microbes, it brings a disturbed gut ecology (Kang et al., 2022). Therefore, together with the results of the previous assays, we hypothesize that C. difficile spores present in the food chain are likely to germinate in the presence of baijiu and thus cause infection. More than 3 days after infection, the cecum of mice in S1, S2, and EtOH groups showed more severe edema, destruction of intestinal villi, an influx of polymorphonuclear cells into the lamina propria, crypt erosion, loss and proliferation of goblet cells compared with the dead mice in the C. difficile group (Fig. 5D). At time 4 (T4), the cecum tissue of euthanized mice in C. difficile, S1 recovered to the level of NC group, but S2 and EtOH groups still had slight edema.
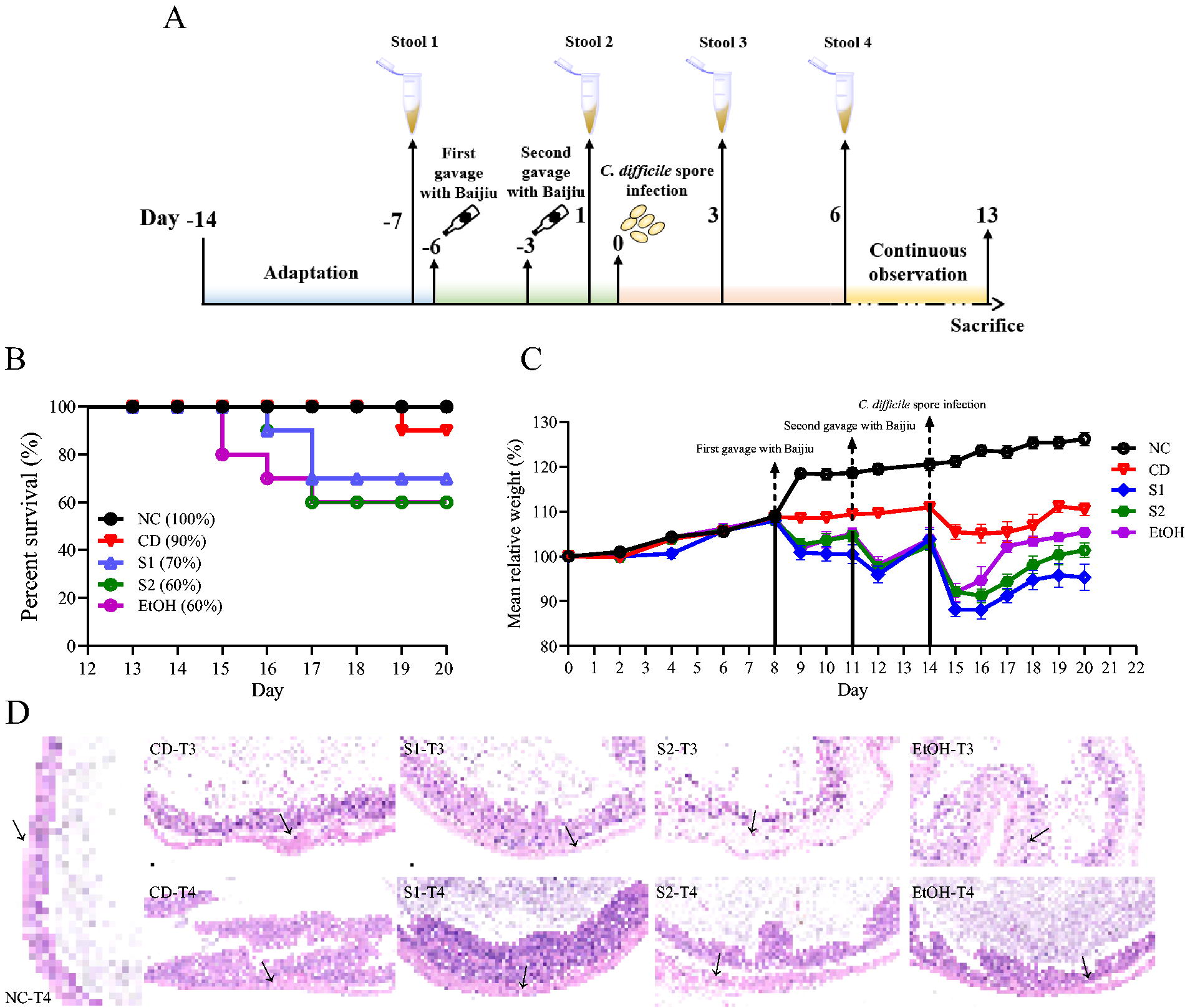
Clostridioides difficile spore infection in a mouse model.
Toxin level, C. difficile spore quantity, Lcn-2, and bile acid content in mice feces
After three days of infection with C. difficile spores, the TcdA level and the number of C. difficile spores in the feces increased rapidly, with significant differences between C. difficile, S1, S2, and EtOH. The C. difficile group reared normally without any treatment before infection showed better resistance after infection, mainly in terms of relatively low TcdA level and C. difficile spore count (Fig. 6A and B), indicating that the natural intestinal barrier was able to contain the viability of C. difficile spores better. After infection, the fecal TcdA levels in S1, S2, and EtOH exceeded 200 ng/g. S1, S2, and EtOH maintained high levels of TcdA and C. difficile spores even at the end of the assay. The intake of baijiu and EtOH directly led to elevated Lcn-2 levels (significant difference compared with predrinking), and this indicator was further exacerbated after C. difficile spore infection, reaching levels above 60 ng/g (except for NC). Lcn-2 is an innate immune protein and a key iron-regulating protein that not only stabilizes the host iron pool in host nutritional immunity but also contributes to the reduction of systemic, cellular, and mucosal blood iron levels during inflammation (Xiao et al., 2017). Previous studies have shown that Lcn-2 plays a protective role primarily in infectious and inflammatory bowel diseases, but has a dual role in neurodegenerative diseases, metabolic syndrome, renal diseases, dermatologic conditions, and cancers (Xiao et al., 2017). Changes in Lcn-2 content can reflect the level of intestinal inflammation (Daniel et al., 2023). At T4, Lcn-2 levels began to fall back but did not reach the level at T2 in the C. difficile, S1, S2, and EtOH groups, with the greatest decrease in the C. difficile group (Fig. 6C), indicating that intestinal inflammation was alleviated. In conjunction with the changes in Lcn-2 levels at different time points, it appeared that baijiu and EtOH caused disturbances in the intestinal tract of mice that contributed to an inflammatory intestinal ecology, and this level did not return to the level at T1 even three days after the cessation of baijiu intake. Previous studies have shown that the type and concentration of bile acids in the gut can directly affect the metabolic activity of C. difficile, with primary bile acids (e.g., CA) promoting CDI and spore germination and secondary bile acids (e.g., DCA) having the opposite effect (Yang and Yang, 2019). In this study, CA levels in the gut of mice in the baijiu and EtOH groups increased rapidly and were accompanied by a rapid decrease in DCA levels, a trend that was further exacerbated by the invasion and infection of C. difficile spores in both bile acid levels (Fig. 6D). At the endpoint of the experiment, DCA levels did not return to preinfection levels in S1, S2, and EtOH, except for C. difficile, where DCA levels returned to preinfection levels (Fig. 6E). These data suggested that the disturbance of bile acid metabolism in the gut ecology of mice induced by baijiu is persistent, and this secondary bile acid environment represented by low levels of DCA provides the conditions for C. difficile spore germination and infection.

Fecal Clostridioides difficile toxin content and quantity, Lcn-2, bile acid content in mice.
Conclusion
In this study, two kinds of commercially available baijiu with the same alcoholic concentration were used to treat C. difficile cells, and the effects of baijiu on C. difficile growth, biofilm production, spore production, virulence gene, and sporulation gene expression were investigated separately, and it was found that alcohol in baijiu upregulated the expression level of C. difficile sporulation gene Spo0A, enhanced the spore production of C. difficile, and increased the adhesion of C. difficile to Caco-2 cells. It was further confirmed by a mouse model that the gut changes induced by baijiu were favorable for the invasion, germination, and infection of C. difficile spores, and this process was accompanied by the increase of gut Lcn-2 and CA levels and the decrease of DCA. This study suggests that there may be a link between baijiu and C. difficile spore invasion and infection. Individuals with sensitive intestinal functions, such as the elderly and those with primary intestinal disorders (e.g., IBD), should be cautious when consuming baijiu.
Footnotes
Authors’ Contributions
W.R. and J.Y.: Conceptualization, investigation, writing—original draft, and writing—review and editing. X.C. and C.S.: Conceptualization and investigation. S.Z. and X.L.: Investigation and software. W.R. and S.Z.: Methodology and visualization. X.C.: Methodology and data curation. J.Y.: supervision, funding acquisition, and writing—review and editing.
Data Availability
Data will be made available on request.
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 32200154).