
Abstract
PrfA is a key virulence regulator for Listeria monocytogenes (Lm) responding to host environment. Here we report that the natural mutation in PrfAK10N/T151A enhanced the pathogenicity of hypervirulent serotype 4h L. monocytogenes. We characterized the phylogenetic tree of PrfA, and found that PrfAK10N/T151A prevalently distributed in all serotype 4h isolates. Remarkably, the growth rate of serotype 4h strain Lm XYSN was significantly slower than EGD-e, in contrast, the substitution mutant Lm PrfAN10K/A151T increased the growth rate of L. monocytogenes. Notably, PrfAK10N/T151A upregulated the expression of multiple virulent genes of Lm XYSN cultured in brain-heart infusion medium, and increased the invasion ability in HeLa and Caco-2 cells. Importantly, the PrfAK10N/T151A mutation significantly enhanced the colonization and survival of Lm XYSN in vivo. Therefore, our findings indicate that the natural mutation of PrfAK10N/T151A enhances the PrfA activity, resulting in the upregulation of various virulence genes and contributing to virulence and pathogenesis of Lm XYSN, which contributes to the hypervirulence of serotype 4h isolates.
Introduction
Listeria monocytogenes (Lm) is a ubiquitous Gram-positive pathogen, which can survive as a saprophyte in soil and can also become intracellular bacteria by entering the host through contaminated food (Xayarath et al., 2015). During the transition from a free-living saprophytic state to a pathogenic parasitic lifestyle, the master regulator PrfA controls the expression of major virulence genes associated with L. monocytogenes infection of various nonphagocytic cells and macrophage cells, such as virulence genes located in Listeria virulence island-1 and internalin family genes(de las Heras et al., 2011). Among the virulence genes regulated by PrfA, bsh and bilE enhance L. monocytogenes bile salt tolerance in the gastrointestinal tract (Dussurget et al., 2002; Sleator et al., 2005). Additionally, inlA, inlB, and hly are involved in invading multiple host cells, hly, plcA, plcB, mpl, and prsA2 are required for disrupting phagocytic membranes, hpt and prsA2 are related to Listeria replication in cytoplasm, and actA is associated with Listeria cell-to-cell spread (Scortti et al., 2007). Overall, PrfA’s transcriptional activation contributes significantly to L. monocytogenes survival within the host and its ability to cause infection and disease.
The activation of PrfA requires to bind with the cofactor glutathione (GSH) (Hall et al., 2016; Reniere et al., 2015), thus the helix-turn-helix motif is stabilized in a conformation compatible with DNA-binding, thereby promoting the stably binding with PrfA box sequences in the promoter of virulence genes (Hall et al., 2016). This interaction facilitates the activation of these genes, leading to their increased expression during infection. The constitutive activation of PrfA have been identified in various serotypes of L. monocytogenes strains (referred to as PrfA* mutants) (Hansen et al., 2020; Miner et al., 2008; Ripio et al., 1997; Shetron-Rama et al., 2003; Vega et al., 2004; Wong and Freitag, 2004; Xayarath et al., 2011), consequently, the virulence genes regulated by PrfA are highly expressed, resulting in hyperinvasive and hypervirulent PrfA* mutants in mouse.
PrfA is a highly conserved protein in L. monocytogenes, with naturally occurring mutations resulting in its constitutive activation being reported only in EGD (PrfAG145S, C229Y) and P14 (PrfAG145S) strains (Bécavin et al., 2014; Vasanthakrishnan et al., 2015). Here, we identify two amino acid changes of PrfA in all serotype 4h strains, which exhibit virulence levels of 200–400 times higher than EGD-e when administered via intragastrical infection, suggesting a high susceptibility of these strains to mice through gastrointestinal infection (Yin et al., 2019). This study characterizes the novel spontaneous mutations (K10N, T151A) in PrfA of hypervirulent Lm XYSN, which enhance Lm XYSN gut colonization and survival within the host.
Materials and Methods
Bacterial strains, cell lines, and animals
The bacteria and plasmids used in this study are listed in Supplementary Table S1. All L. monocytogenes strains were cultured in brain-heart infusion (BHI; Becton Dickenson, USA). Escherichia coli DH5α were cultured in Luria-Bertani media (LB; Sangon, China). HeLa cells and Caco-2 cells were obtained from the American Type Culture Collection, and cultured at 37°C under 5% CO2 in DMEM (Gibco, USA) supplemented with 10% FBS (MeisenCTCC, China). Six-week-old female C57BL/6 mice were purchased from Vital River Laboratory Animal Technology Co., Ltd (Beijing, China).
Construction PrfA substitution mutants
The PrfA substitution mutants of Lm XYSN was generated using a homologous recombination strategy. The oligonucleotide primer sequences used in this process can be found in Supplementary Table S2. Fragments containing prfAN10K/A151T or prfAN10K/G145S/A151T mutations were ligated to the pAUL-A shuttle vector and then introduced into Lm XYSN competent cells by electroporation. After the plasmids were removed, the Lm PrfAN10K/A151T and Lm PrfAN10K/G145S/A151T substitution mutants were identified by sequencing.
Bioinformatics analysis
Amino acid sequences of 47 types of PrfA in L. monocytogenes were downloaded from National Center for Biotechnology Information (https://www.ncbi.nlm.nih.gov/genome/annotation_prok/evidence/NBR001116/), and representative strains of each type were randomly selected based on PrfA protein sequences, totaling 52,034 isolates. The PrfA protein sequences of 47 representative strains were aligned using BioEdit and imported into MEGA11. Based on the alignment, we created the neighbor-joining phylogram, including 1000 bootstrap replicates. PrfA sequences of Lm XYSN and other L. monocytogenes strains were compared by ClustalX. The PrfAK10N/T151A and PrfAN10K/A151T were submitted to SWISS-MODEL server (https://www.swissmodel.expasy.org/) for automatic modeling to predict their three-dimensional structures.
Growth curve analysis
Bacteria were cultured at 37°C and adjusted to an initial optical density (OD)600 value of 0.05 using the BHI medium. Three replicates of each strain were added to separate wells of a 96-well cell culture plate, with each well containing 200 μL of the bacterial suspension. The plate was then placed in a Spark multifunction microplate reader (TECAN, Switzerland) and incubated at 37°C with shaking. The OD600 value of the bacteria cultures was measured at 30 min interval.
Quantitative real-time polymerase chain reaction
Overnight cultures of bacteria were inoculated into fresh BHI medium and grown to an OD600 value of 0.8. Total RNA from bacterial cultures was extracted using the RNAprep Pure Cell/Bacteria Kit (TIANGEN, China). Note that 1 µg total RNA was reverse transcribed into cDNA through the HiScript® IIl RT SuperMix for quantitative polymerase chain reaction (qPCR) with gDNA Eraser (Vazyme, China). Primers used for quantitative real-time PCR (qRT-PCR) are listed in Supplementary Table S3. qRTPCR was performed using AceQ Universal SYBR qPCR Master Mix (Vazyme, China) on the QuantStudioTM 6 Flex real-time PCR system (Bio-Rad). gyrB was selected as the internal reference gene, and the relative changes in expression were calculated by the 2-ΔΔCt method (Livak and Schmittgen, 2001).
Western blot analysis
Overnight cultures were diluted 100-fold into 100 mL of fresh BHI medium in Erlenmeyer flasks and grown to an OD600 value of 0.8 after 4∼4.5 h. Intracellular proteins were obtained through ultrasonic disruption, while the method for extracting secreted proteins was performed as described by Yin et al. (Yin et al., 2019). The PrfA antibodies were kindly provided by Dr. Pascale Cossart, while the LLO and GAPDH antibodies were prepared in our laboratory. Secondary antibodies used were HRP-labeled goat anti-mouse IgG and HRP-labeled goat anti-rabbit IgG (Immunoway, America).
Invasion and replication assays in HeLa and Caco-2 cells
HeLa or Caco-2 cells were cultured in 24-well plates at 37°C with 5% CO2 for 12∼16 h. Overnight cultures of bacteria were inoculated into fresh BHI medium and grown until reaching an OD600 value of 0.8. Monolayers of cells were infected with L. monocytogenes at a bacterium/cell ratio of 20 for 1 h. Following, the cells were washed twice with PBS and further incubated either for 15 min (invasion assay) or 2 h (replication assay) with DMEM medium containing 50 μg/mL gentamicin sulfate. After which, the cells were lysed with 0.2% Triton X-100 buffer for 9 min to release the bacteria. The cell lysate was serially diluted and spread onto BHI plates to determine the number of bacteria present.
Virulence in mouse model
Overnight bacterial cultures were diluted into fresh media and grown at 37°C until reaching an OD600 value of 0.8. The bacteria were washed twice with PBS and adjusted the OD600 of the bacteria to 0.78 (about 1 × 109 colony-forming unit [CFU]/mL). Orogastric virulence assays were performed with 6-week-old female C57BL/6 mice (n = 6/group) using a dose of 3 × 106 CFU of L. monocytogenes. After infection for 72 h, the livers, spleens, ileum, and colon of mice were harvested to determine bacterial loads of L. monocytogenes.
Results
PrfA of serotype 4h isolates contains unique spontaneous mutations
The phylogenetic tree was constructed with the amino acid sequences of PrfA protein, the entire 52,034 isolates were grouped into six clusters, 95.3% isolates (49,588/52,034) belonged to cluster I, among which 98.8% isolates had 100% homology with PrfA protein of EGD-e, suggesting that PrfA of most L. monocytogenes are highly conserved (Fig. 1). Interestingly, the cluster VI consisted of isolates belonging to lineage III and IV, which exhibited 2, 3, or 4 amino acids differences compared to PrfA of EGD-e. Considering PrfA’s role in regulating various virulence proteins, we hypothesize that the PrfA mutations may be linked to the hypovirulence observed in cluster VI strains.

Phylogenetic trees of 47 PrfA proteins. The tree was constructed using the neighbor-joining method, including 1000 bootstrap replicates. Number of strains represents the number of strains containing the corresponding type of PrfA mutation. **** represents the reference PrfA of EGD-e, ± aa represents the number of amino acids that are more or less than the reference PrfA, the full name of E005-2 strain is CDPHFDLB-F15M04169-E005-2.
Unlike other types of PrfA characterized with one amino acid mutation, two natural amino acid mutations K10N/T151A were found in serotype 4h isolates belonging to cluster I, at amino acid sites 10 and 151, respectively (Fig. 2A). The PrfAK10N/T151A and PrfAN10K/A151T were submitted to SWISS-MODEL for automatic modeling to obtain the spatial structures, and analyzed the structure of the two proteins by PyMOL. Two mutated sites in PrfA could be found in the three-dimensional structure directly (Fig. 2B and 2C). One mutated site was in the αA (amino acid resides 3–14) and the other was in the αD (amino acid resides 138–154). Mutations present in the αA (amino acid resides 3–14) reduce the complexity of the side chain and its interaction with T-14, and the mutation present in αD reduce the interaction with L-147 due to the reduction of the side chain.
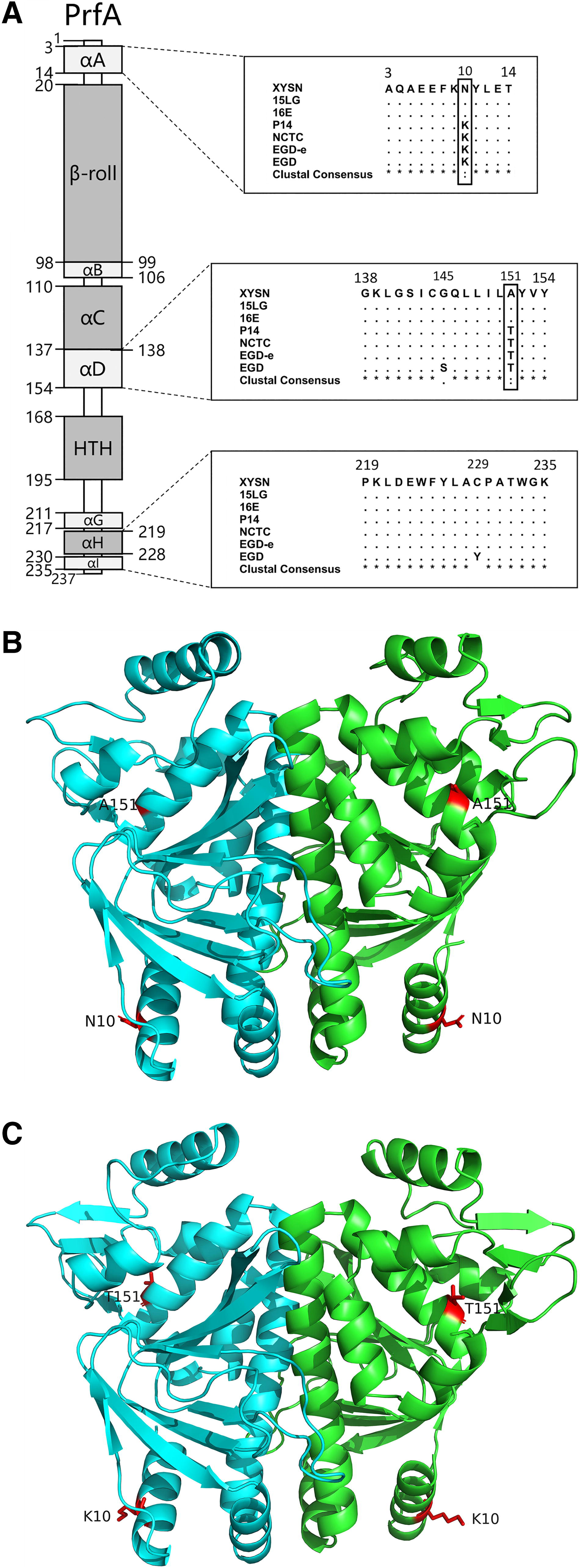
Bioinformatics analysis of PrfA protein.
The PrfAK10N/T151A mutation affects the bacterial growth
It has been reported that overexpression of the PrfA regulator leads to growth defects of L. monocytogenes in BHI medium (Bécavin et al., 2014; Vasanthakrishnan et al., 2015). To evaluate the effect of PrfAK10N/T151A mutation in regulating Lm XYSN survival in vitro, the growth ability of Lm XYSN, Lm PrfAN10K/A151T, Lm PrfAN10K/G145S/A151T, and EGD-e under the BHI medium was monitored at 37°C. The growth rates of the other strains were significantly slower compared with EGD-e. The strain Lm PrfAN10K/A151T exhibited a significantly enhanced growth rate compared with the wild-type strain Lm XYSN. Conversely, Lm PrfAN10K/G145S/A151T demonstrated a significantly reduced growth rate than Lm XYSN (Fig. 3A). The results indicate that the PrfAK10N/T151A mutation in serotype 4h L. monocytogenes affected the growth capacity of strains, potentially due to its enhanced activity.

Growth curve and expression analysis of PrfA-dependent genes in Listeria monocytogenes (Lm).
The PrfAK10N/T151A mutation causes virulence factor overexpression in L. monocytogenes
To determine whether spontaneous mutation of PrfAK10N/T151A in Lm XYSN affected the expression of PrfA-dependent virulence genes, the transcriptional expressions of prfA, actA, hly, plcA, and mpl were determined in BHI medium. The results showed that the expression levels of PrfA-dependent genes in substitution mutant strain Lm PrfAN10K/A151T were significantly downregulated than those in Lm XYSN. However, Lm PrfAN10K/G145S/A151T showed significantly higher expression levels of these genes compared with Lm XYSN (Fig. 3B). Consistent with the transcriptional expression, Lm PrfAN10K/A151T displayed higher cytoplasmic PrfA protein levels and lower LLO protein secretion levels compared with Lm XYSN. Furthermore, Lm PrfAN10K/G145S/A151T exhibited a significant increase in both PrfA and LLO protein levels when compared with Lm XYSN (Fig. 3C–E). Based on the significant differences in expression levels of PrfA-dependent genes and LLO protein between Lm XYSN and Lm PrfAN10K/A151T, we propose that the PrfAK10N/T151A mutation can increase the activity of PrfA, thus activating the upregulation of downstream virulence genes.
The PrfAK10N/T151A mutation promotes L. monocytogenes invasion ability
The infection capacities of Lm XYSN, Lm PrfAN10K/A151T, Lm PrfAN10K/G145S/A151T, and EGD-e strains were determined using the human cervical carcinoma epithelial cell line HeLa and human colon epithelial cell line Caco-2. The result indicated that the substitution mutant strain Lm PrfAN10K/A151T exhibited significantly reduced invasion and replication abilities in both HeLa and Caco-2 cell lines than Lm XYSN. In contrast, the Lm PrfAN10K/G145S/A151T exhibits significantly enhanced invasion capability in HeLa cells and both invasion and replication capability in Caco-2 cells (Fig. 4A–D). This suggests that the spontaneous mutation of PrfAK10N/T151A in Lm XYSN upregulated the expression of LIPI-1 genes, thus enhanced the infectivity of the bacteria at the cellular level.

Invasion and replication capacities of Listeria monocytogenes (Lm) XYSN, Lm PrfAN10K/A151T, Lm PrfAN10K/G145S/A151T, and EGD-e to
PrfAK10N/T151A mutation contributes to Listerial intestinal colonization and virulence
The colonization ability of Lm XYSN, Lm PrfAN10K/A151T, Lm PrfAN10K/G145S/A151T, and EGD-e in the liver, spleen, ileum, and colon of C57BL/6 mice was measured after inoculation for 72 h. Compared with PrfAN10K/A151T, the bacterial loads in the ileum and colon of mice significantly increased postinfection with Lm XYSN (ileum: 4388-fold; colon: 76-fold) and Lm PrfAN10K/G145S/A151T (ileum: 9065-fold; colon:1125-fold; liver: 4-fold; spleen: 56-fold). Additionally, mice inoculated with EGD-e showed no bacterial colonization in any organ (Fig. 5A–D). These results suggest that the spontaneous mutation of PrfAK10N/T151A enhanced the bacterial colonization ability in the mouse gut, thus enhancing the virulence of the bacteria.

Virulence of Listeria monocytogenes (Lm) XYSN, Lm PrfAN10K/A151T, Lm PrfAN10K/G145S/A151T, and EGD-e. Bacterial loads in
Discussion
Previous studies have demonstrated that activation of PrfA converted L. monocytogenes from an environmental bacterium into a pathogen capable of replicating and spreading within host cells (Freitag et al., 2009; Toledo-Arana et al., 2009). Additionally, the constitutive activation of PrfA in the PrfA* strain has been found to have diverse effects on bacterial phenotype and virulence. The mutation sites of most PrfA* mutants were located in the αD helix, such as L140F, G145S, Y154C, G155S, and others. Among these mutations, the G145S mutation is a spontaneous mutation that results in constitutive expression of virulence genes products (Bécavin et al., 2014; Vasanthakrishnan et al., 2015). In our study, we identified two spontaneous mutations, K10N and T151A, present in all serotype 4h Listeria isolates, located in the αA and αD helices, respectively. The functional significance of the αA helix was not yet fully understood, while the αD helix was the mutation position of many PrfA* mutants. Our findings demonstrated that the natural unique mutation in PrfAK10N/T151A enhances PrfA transcriptional activity, contributing to the hypervirulence of the 4h serotype L. monocytogenes strain XYSN.
PrfA typically exhibits lower activity in vitro, leading to weak regulation of downstream virulence genes. However, upon entering host cells in vivo, PrfA undergoes allosteric activation through its binding to the cofactor GSH, resulting in higher expression of downstream genes (Chatterjee et al., 2006; Joseph et al., 2006; Moors et al., 1999; Reniere et al., 2015, 2016; Shetron-Rama et al., 2002; Toledo-Arana et al., 2009). Interestingly, Vasanthakrishnan et al. found that the PrfA* G145S mutation results in elevated expression of virulence genes, imposing a huge burden on L. monocytogenes outside the host and limiting its survival and competitiveness (Vasanthakrishnan et al., 2015). In this study, we observed that Lm XYSN grew at a slower rate than the substitution mutant strain Lm PrfAN10K/A151T, while the Lm PrfAN10K/G145S/A151T showed an even greater reduction in growth rate. This suggests that the PrfAK10N/T151A mutation may enhance the activity of the PrfA protein, thereby affecting bacterial growth. Furthermore, we discovered that the spontaneous PrfAK10N/T151A mutation upregulated the expression of downstream virulence genes (actA, hly, plcA, mpl) in vitro, and enhanced the secretion of LLO protein in culture supernatants, yet did not significantly affect PrfA protein expression. Therefore, these findings indicate that the spontaneous mutation of PrfAK10N/T151A leads to enhanced transcriptional activity of PrfA and imposes a significant growth cost on bacteria in vitro.
PrfA is the key virulence factor of L. monocytogenes, globally regulating the expression of virulent and virulence-associated genes within the host. The PrfA* mutants locked the protein into a constitutively active state, enhancing the affinity of PrfA binding to the promoter of virulence-regulating genes, ultimately leading to various physiological changes of Listeria (Vega et al., 2004). The PrfA* mutants (L140F, G145S, A218G, P219S) were highly invasive to tissue culture cells, increased bacterial fitness within infected host cells, and highly pathogenic in mouse infection models (Bruno and Freitag, 2010; Hansen et al., 2020; Xayarath et al., 2011). In this study, the hypervirulent strain Lm XYSN carrying a spontaneous PrfAK10N/T151A mutation activates the expression of virulence genes, significantly enhancing the invasion capacity to HeLa and Caco-2 cell lines, and promoting the colonization ability in the ileum and colon of mice, indicating that the PrfAK10N/T151A spontaneous mutation enhanced Listerial virulence. It is worth mentioning that PrfAK10N/T151A is uniquely distributed in serotype 4h L. monocytogenes strains, we propose that the spontaneous mutation contributes to Lm XYSN break through the intestinal barriers, facilitating the hypervirulence of serotype 4h isolates (Yin et al., 2019). However, further investigation is needed to decipher the crystal structure of PrfAK10N/T151A, thus fully understand the mechanism of enhancing the regulation activity and the virulence of 4h L. monocytogenes strains.
Conclusions
L. monocytogenes is a highly variable species that is classified into 47 types based on amino acid diversity of major virulence regulator PrfA. Serotype 4h L. monocytogenes have unique PrfAK10N/T151A mutations, resulting in bacterial cells growing slower, but significantly upregulated multiple virulence genes expression. PrfAK10N/T151A mutations strikingly promoted bacteria invasion ability to HeLa and Caco-2 cells, and enhanced the virulence of L. monocytogenes in vivo. Altogether, the spontaneous mutation of PrfAK10N/T151A facilitates the infection and pathogenicity of serotype 4h strains.
Authors’ Contributions
Z.W., Y.Y., and X.J. designed the experiments. J.L. and X.C. performed the experiments and analyzed the results. Z.W. and J.L. participated in the bioinformatics analysis. L.W. and R.W. were involved in Western blot analysis. Y.W. and F.M. participated in cell experiments. J.L. and Y.Y. wrote the article. All authors contributed to the article and approved the submitted version.
Footnotes
Acknowledgments
The authors are grateful to Dr. Xiqing Wang for technical assistance with protein structure analysis. The authors wish to acknowledge the generous gift of the shuttle vector pAUL-A by Prof. Trinad Chakraborty and the PrfA antibody by Dr. Pascale Cossart.
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 32161133025, 32373012), Jiangsu Agricultural Science and Technology Independent Innovation Funds (CX(21)1004), Key Research and Development Program of Jiangsu (BE2021331), and the Priority Academic Development Program of Jiangsu Higher Education Institutions (PAPD).
Data Availability
The data used to support the findings of this study are available from the corresponding author upon request.
Ethics Statement
Animal experiments were conducted by following per under guidelines laid down for the welfare and ethics of experimental animals. All animals were kept at the animal biosafety facilities according to the procedures approved by the Institutional Animal Ethics Committee of Yangzhou University.
Disclosure Statement
All authors declare no conflicts of interest.