
Abstract
Unconsidered microorganisms, such as Pseudomonas aeruginosa, may be often recovered from food samples. This study aimed to characterize seven P. aeruginosa recovered from traditional market chicken meat in Lima, Peru. Antimicrobial susceptibility to 18 antimicrobial agents as well as the presence of amino acid changes in fluoroquinolone targets, 10 mediated colistin resistance (mcr) genes and integrons were analyzed. Clonal relationships were determined through pulsed-field gel electrophoresis (PFGE). Serotype by agglutination, multilocus sequence typing and the presence of 14 virulence factors (VFs) were established. Two isolates were multidrug-resistant, all being fluoroquinolone-resistant and exhibited the amino acid changes GyrA T83I and ParC S87L. No mcr gene was detected in the colistin-resistant isolates. The isolates showed identical PFGE patterns, and the selected P6 isolate belonged to the serotype O:4 and the sequence type 1800, and presented 12 VFs (all but exoU and exlA). The present study highlights the presence of multidrug and virulent P. aeruginosa in market chicken meat, and suggests cross-contamination during meat manipulation.
Keywords
Introduction
Pseudomonas spp. is a ubiquitous genus often isolated from water and soil samples, as well as member of human, animal or plant microbiomes, with several species being able to cause human diseases (Estepa et al., 2014; Horna et al., 2019a; Ruiz-Roldán et al., 2020; Silby et al., 2011). Among the pathogenic Pseudomonas spp., Pseudomonas aeruginosa is by far the most relevant. P. aeruginosa usually acts as an opportunistic pathogen, often being isolated from sensitive patients, such as those hospitalized in burn units or intensive care units (Flores-Paredes et al., 2021; Horna et al., 2019a). Infections related to P. aeruginosa are severe and difficult to treat because of the high levels of antimicrobial resistance often exhibited by this microorganism (Flores-Paredes et al., 2021; Horcajada et al., 2019; Horna et al., 2019b), with emerging resistance to last resort antibacterial agents, such as carbapenems and colistin (Gómez et al., 2023; Xiao et al., 2022). In addition, P. aeruginosa possess an extensive armamentarium of virulence factors (VFs), including toxins, siderophores, and adhesins, among others, which contributes to enhancing its pathogenic power (Horna et al., 2019a; Li et al., 2024; Ruiz-Roldán et al., 2020; Strateva and Mitov, 2011). Among these, ExoU and ExoS, effectors of the so-called Type 3 secretion system, ranks among the most relevant VFs, with ExoU exhibiting a potent cytotoxic effect and ExoS being involved in apoptosis process and disruption of the actin cytoskeleton (Horna and Ruiz, 2021). Furthermore, P. aeruginosa has the ability to live in benthonic communities (Horna et al., 2019b), conforming difficult to eradicate biofilms of important clinical significance when present in clinical devices (Tuon et al., 2022). Overall, these traits lead to bad outcomes and high lethality levels (Oliveira de Matos et al., 2018). A series of circulating P. aeruginosa sequence type (ST) have been classified as “high risk clones,” as for instance ST111, ST175 or ST235, because its large dissemination, virulence power, and high levels of antimicrobial resistance (Horcajada et al., 2019; Juan et al., 2017). Among these, isolates qualifying as hypervirulent are of special concern (Horcajada et al., 2019; Li et al., 2024). In fact, the relevance of P. aeruginosa as a human pathogen is highlighted by its inclusion among the so-called “ESKAPE” group (Rice, 2008).
The food chain is a route of microorganism dispersion (Ruiz-Roldán et al., 2018). The presence of human pathogenic microorganisms, such as Listeria monocytogenes, Salmonella enterica or diarrheagenic Escherichia coli in food samples is well-established and considered as classical foodborne pathogens (Enciso-Martínez et al., 2022; Martínez-Puchol et al., 2021; McLauchlin et al., 2020). However, other pathogens such as Acinetobacter spp. or Pseudomonas spp., although described, is often overlooked (Marí-Almirall et al., 2019; Ruiz-Roldán et al., 2021).
Regarding food chain, a recent study developed in Australia, focused on the detection of extended-spectrum beta-lactamase-producing microorganisms from chicken and pork meats, highlighted the presence of different species of Pseudomonas resistant to 3rd generation cephalosporins, including P. aeruginosa (McLellan et al., 2018). Meanwhile a study designed to determine the presence of P. aeruginosa in marketed pork and chicken meat in China, resulted in the isolation of 109 P. aeruginosa with overall low levels of antibiotic resistance (Wu et al., 2023). Antibiotic resistance levels of meat-recovered P. aeruginosa varies among studies.
In this scenario, the aim of this study was to characterize VFs, antibiotic resistance, and the clonal relationships of P. aeruginosa isolated from market meat in Lima, Peru.
Materials and Methods
Samples and isolation method
In November 2021, during the course of a study designed to determine the presence of Enterobacterales in marketed meat (14 chicken and 7 beef samples) (Guibert et al., 2023), seven bacterial colonies suspected of being Pseudomonas spp. were recovered. All samples were randomly acquired on traditional markets from Southern Lima, sterile transported to the laboratory and processed within 1−3 h after collection (Guibert et al., 2023).
Bacterial identification
DNA extraction of the microorganisms suspected to be P. aeruginosa was performed by boiling (100°C, 10 min) a bacterial suspension in sterile water. Thereafter, the 16S rRNA was amplified following established procedures (Salazar de Vegas et al., 2006), gel recovered (Omega Bio Tek, Norcross, USA) and sequenced (Macrogen, Seoul, South Korea). The proteins were extracted using ethanol-formic acid as per the manufacturer’s guidelines, in order to confirm the identification by matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) using a MALDI Biotyper® (Bruker Daltonics GmbH & Co. KG, Bremen, Germany). The results were analyzed following the MBT Compass Library DB-6903 (V.6) from May 2016.
Antimicrobial susceptibility
The susceptibility to 17 antibacterial agents, including ciprofloxacin (5 µg), levofloxacin (5 µg), gentamicin (10 µg), amikacin (30 µg), tobramycin (10 µg), netilmicin (30 µg), piperacillin (100 µg), ceftazidime (30 µg), cefepime (30 µg), cefiderocol (30 µg), piperacillin–tazobactam (100/10 μg), ceftolozane–tazobactam (30/10 μg), ceftazidime–avibactam (30/20 µg), aztreonam (30 µg), imipenem (10 µg), meropenem (10 µg/mL), and doripenem (10 µg), was established by disk diffusion in accordance with Clinical & Laboratory Standards Institute (CLSI) guidelines (CLSI, 2022). Meanwhile, susceptibility to colistin was first screened by colistin agar spot, and isolates growing on 2 µg/mL of colistin were subsequently evaluated by microdilution (CLSI, 2022; Naomi-Matsuoka et al., 2020). Multidrug resistance was defined following the criteria of Magiorakos et al. (2012).
Induction of chromosomal AmpC
The induction of the chromosomal AmpC was determined by the ceftazidime-imipenem antagonism test as previously described (Cantarelli et al., 2007). A flattening of the ceftazidime halo was considered as a positive result for AmpC production.
Fluoroquinolone resistance mechanisms
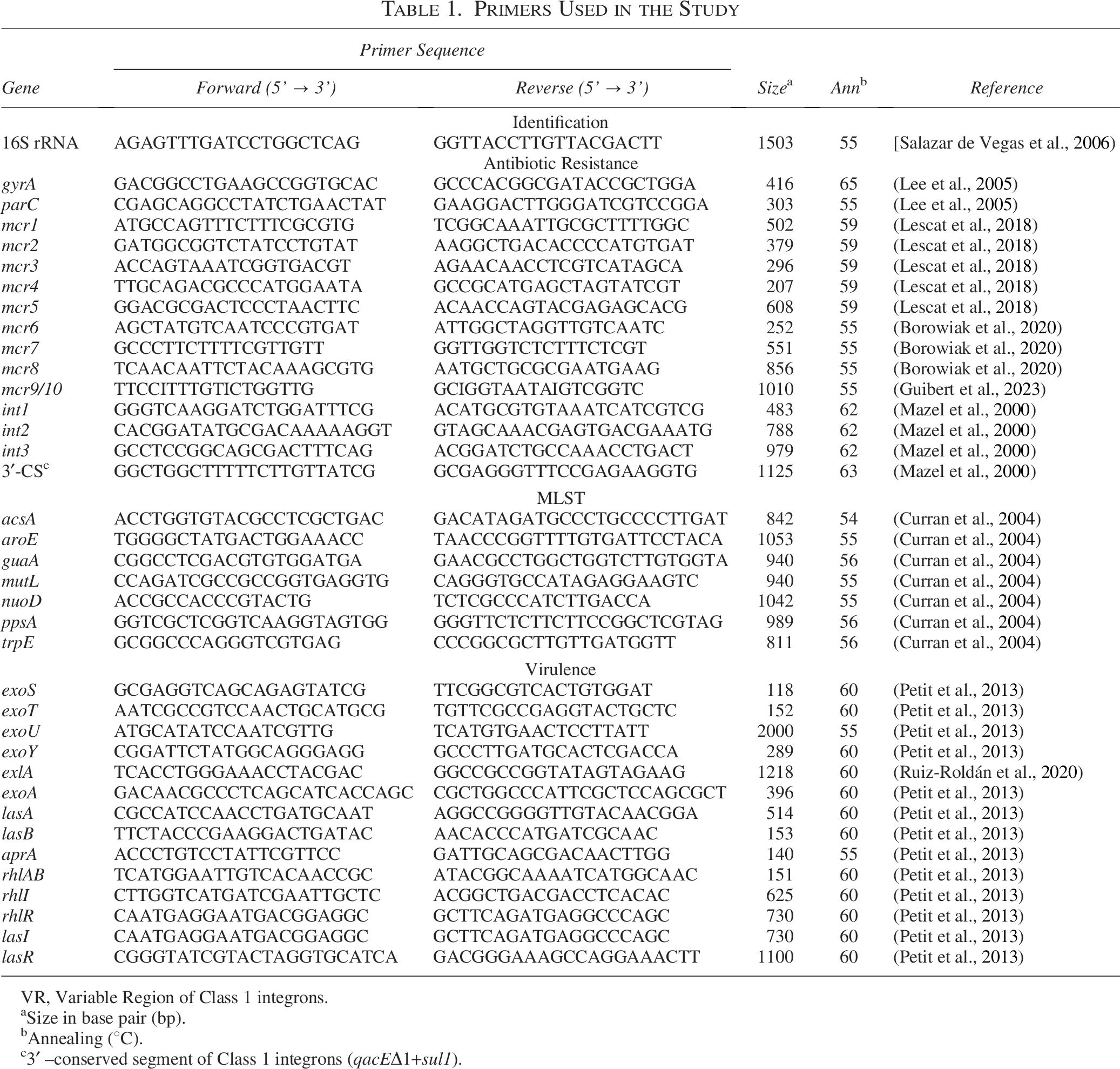
The gyrA and parC genes were analyzed by PCR (Lee et al., 2005) and sequencing to determine the alterations in the quinolone-resistance determining regions within GyrA and ParC proteins (Table 1).
Primers Used in the Study
VR, Variable Region of Class 1 integrons.
Size in base pair (bp).
Annealing (ºC).
3′ –conserved segment of Class 1 integrons (qacEΔ1+sul1).
Mcr variants
The presence of 10 mobile colistin resistance (mcr) genes (mcr-1 to mcr-10) was established following previously published multiplex protocols. Thus, mcr-1 to mcr-5 were amplified following the methodology described by Lescat et al. (2018), while mcr-6 to mcr-10 were amplified following the Borowiak et al. (2020) methodology (Table 1). In both cases, the methodology was slightly modified, with amplifications being made gene by gene, instead of in a multiplex PCR, and 40 pmol (mcr-1 to mcr-5) or 20 pmol (mcr-6 to mcr-10) were used.
Presence of integrons
The presence of genes encoding type 1, 2, and 3 integrases, as well as the class 1 integrons 3′–conserved segment (qacEΔ1+sul1) was established by PCR (Table 1) (Mazel et al., 2000).
Clonal relationships
Clonal relationships were established through pulsed-field gel electrophoresis (PFGE) using the CHEF-DRIII system (Bio-Rad Laboratories Inc., Hercules, USA) as previously described (Estepa et al., 2014). A dendrogram was constructed using the GelJv2.0 software (https://sourceforge.net/projects/gelj/) (Heras et al., 2015). Clustering was established following the Unweighted Pair Group Method with Arithmetic Mean considering a 1% tolerance in band positions. The DICE coefficient was used to establish clonal relationships. An identity ≥90% was considered to classify isolates as indistinguishable, and ≥80% as belonging to the same epidemiological clonal group (Guiral et al., 2018).
Multilocus sequence typing
The ST was established following standard procedures (Table 1) (Curran et al., 2004).
Serotyping
Serotyping was performed by slide agglutination using O:1 to O:16 monovalent antisera according to the manufacturer's recommendations (BIORAD, Marnes-la-Coquette, France).
Presence of VFs
The presence of the exoS, exoT, exoU, and exoY genes encoding Type 3 Secretion System effectors, as well as exoA, exlA, lasA, lasB, aprA, rhlAB, rhlI, rhlR, lasI, and lasR were determined by PCR following previously established procedures (Table 1) (Petit et al., 2013; Ruiz-Roldán et al., 2020).
Results
Seven bacterial colonies suspected of being Pseudomonas spp. were recovered from the meat samples of three chickens, each from a different retail stall in the same market (Mercado El Sol, Villa El Salvador, Southern Lima). In all cases, analysis of the 16S rRNA showed the presence of P. aeruginosa, which was thereafter, confirmed as P. aeruginosa by MALDI-TOF.
The seven isolates presented slight differences in the antimicrobial resistance profiles, with all being resistant to fluoroquinolones (ciprofloxacin and levofloxacin), and two isolates were intermediate resistant to aztreonam and another one to netilmicin. Of note, three isolates were resistant to colistin. Overall, two isolates were multidrug-resistant (MDR) (Table 2).
Antimicrobial Susceptibility of Pseudomonas aeruginosa.
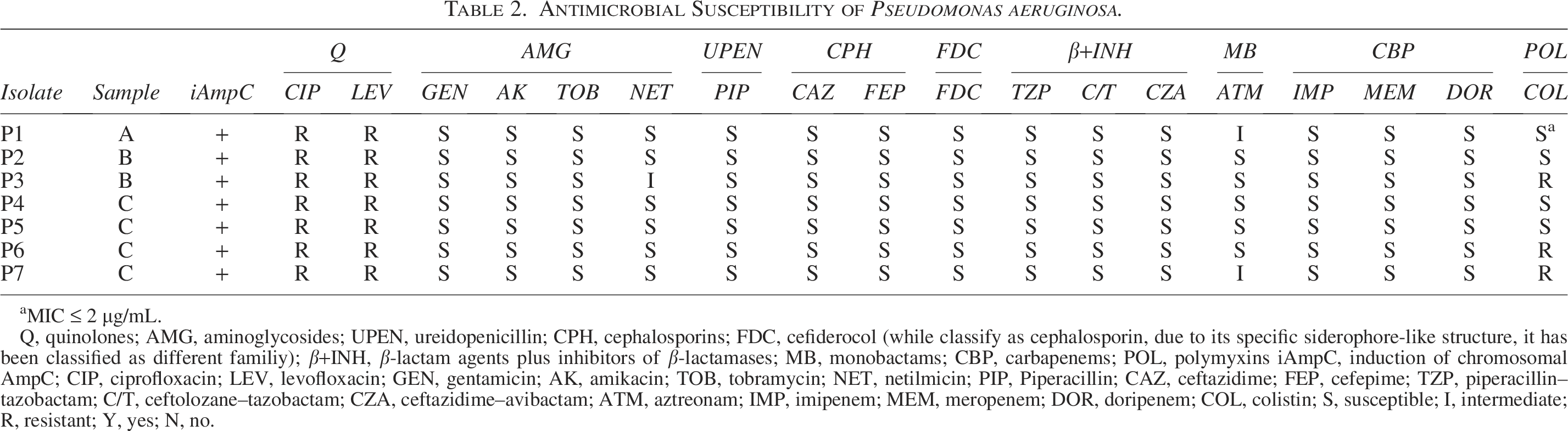
MIC ≤ 2 µg/mL.
Q, quinolones; AMG, aminoglycosides; UPEN, ureidopenicillin; CPH, cephalosporins; FDC, cefiderocol (while classify as cephalosporin, due to its specific siderophore-like structure, it has been classified as different familiy); β+INH, β-lactam agents plus inhibitors of β-lactamases; MB, monobactams; CBP, carbapenems; POL, polymyxins iAmpC, induction of chromosomal AmpC; CIP, ciprofloxacin; LEV, levofloxacin; GEN, gentamicin; AK, amikacin; TOB, tobramycin; NET, netilmicin; PIP, Piperacillin; CAZ, ceftazidime; FEP, cefepime; TZP, piperacillin–tazobactam; C/T, ceftolozane–tazobactam; CZA, ceftazidime–avibactam; ATM, aztreonam; IMP, imipenem; MEM, meropenem; DOR, doripenem; COL, colistin; S, susceptible; I, intermediate; R, resistant; Y, yes; N, no.
Quinolone resistance was related to the presence of the amino acid substitutions T83I and S87L at GyrA and ParC, respectively. Meanwhile, no mcr gene was confirmed in colistin-resistant isolates. All isolates produced the inducible chromosomal AmpC. None of the seven isolates was positive for integrase genes or the presence of qacEΔ1+sul1 region.
Determination of the clonal relationships of the seven isolates showed that they belonged to the same clone (Fig. 1), and isolate P6 was thereby selected for the determination of serotype, multilocus sequence typing (MLST) as well as the presence of VFs. The results showed that the isolate was classified as serotype O:4, belonged to the ST1800 and harbored all the virulence genes sought, with the exception of exoU and exlA.

Dendrogram of similarity of the seven P. aeruginosa isolates.
Discussion
The food chain is an open door for microorganisms to have access to humans. Therefore, food safety is key in preventing the development of a series of diseases, mostly affecting the gastrointestinal tract. Salmonella spp., Campylobacter spp. or diarrheagenic E. coli are classified among the most common pathogenic microorganisms isolated from marketed meat (Benites et al., 2022; Martínez-Puchol et al., 2021; Wang et al., 2017). Nevertheless, other pathogenic microorganisms may also be isolated from food samples, including typical nosocomial pathogens, such as Acinetobacter spp. and P. aeruginosa (Marí-Almirall et al., 2019; Ruiz-Roldán et al., 2021). In Peru, most of the studies aimed at the characterization of microorganisms such as P. aeruginosa are usually focused on clinical isolates (Flores-Paredes et al., 2021; Horna et al., 2019b). While Pseudomonas spp. have previously been isolated from food samples in Peru (Ruiz-Roldán et al., 2018), to the best of our knowledge this is the first molecular and epidemiological analysis of P. aeruginosa isolated from food samples in Peru.
In the present study, all the isolates studied showed resistance to fluoroquinolones (ciprofloxacin and levofloxacin), and three to colistin. Studies of other microorganisms isolated from chicken meat in the country describe a similar scenario, with nonsusceptibility to quinolones reaching >80% among E. coli or 100% among Salmonella enterica serovar Infantis (Guibert et al., 2023; Martínez-Puchol et al., 2021; Ruiz-Roldán et al., 2018). Nevertheless, different to our isolates, Peruvian food-recovered microorganisms tend to show high levels of antimicrobial resistance, being common the presence of MDR isolates (Guibert et al., 2023; Martínez-Puchol et al., 2021; Ruiz-Roldán et al., 2018), and often including isolates resistant to colistin (Guibert et al., 2023). Regarding detected mutation points in gyrA and parC, they have been largely described in P. aeruginosa isolates (López et al., 2022), as well as in the equivalent position of a large series of microorganisms, being by far the most common cause of quinolone resistance (Ruiz, 2003).
Two isolates showed resistance to polymyxins, but no mcr gene was detected. This finding suggests the presence of mutations in chromosomal genes involved in the synthesis of lipid A (Huang et al., 2020). In fact, analysis of other food-recovered microorganisms, while detected the presence of mcr-1, showed high frequency of non mcr colistin-resistant isolates (Guibert et al., 2023).
The isolation of MDR/XDR microorganisms for non human sources has been largely described worldwide, with the large use of antimicrobial agents in veterinary/agriculture together the presence of antibiotic residues or antibiotic-resistant microorganisms in water and solid waste being proposed among the main underlying reasons (Ruiz, 2021).
Unexpectedly, the seven isolates showed identical PFGE patterns. While the option that the selected meat samples had a common origin from the same farm, or that a P. aeruginosa clone was disseminated in the farms of the area cannot be ruled out, the most probable scenario is cross contamination of samples during meat processing. This finding is also supported by the fact that all the samples originated from the same market.
One isolate, isolate P6, which was selected for further analysis, was classified as serotype O:4, and belonged to ST1800. To the best of our knowledge, this ST has been reported four times in the P. aeruginosa MLST database. (https://pubmlst.org/bigsdb?db=pubmlst_paeruginosa_isolates&page=profiles): one (isolate PL28) as a cause of blood infection in Colombia in 2012, two others (isolates AZPAE14925 and AZPAE14913) in 2008 as a cause of intraabdominal infection in Brazil and urinary tract infection in Croatia, respectively, and the remaining isolate (isolate BWH055) had a nonspecified clinical origin, and was of an unreported geographical origin (Kos et al., 2015; Vanegas et al., 2014; https://www.ncbi.nlm.nih.gov/pathogens/isolates/). These data, highlighting the virulence potential of the present isolates, despite neither exoU nor the exlA gene were detected. This finding is in agreement with that described for the 3 P. aeruginosa ST1800 isolates (AZPAE14913, AZPAE14925, and BWH055) present in GenBank (https://www.ncbi.nlm.nih.gov/nuccore/), which possessed the exoS, exoT, and exoY genes. Similarly, the two previous P. aeruginosa ST1800 isolates (AZPAE14913 and AZPAE14925), from which data about serotype were recorded, belonged to the serotype O:4 (Nageeb et al., 2021). Furthermore, recently, Li et al. (2024) described seven extensively resistant hypervirulent P. aeruginosa belonging to serotype ST1800, all presenting the serotype O:4, as a cause of human infections. As in the present study, these isolates possessed three of the four type 3 secretion system effectors (ExoS, ExoY, and ExoT), but no data of other VFs was disclosed. While specific MDR high risk clones, such as ST235, have been associated with poor clinical outcome in sepsis infections, a common scenario is an inverse association between virulence and resistance (Juan et al., 2017). Despite no studies to establish hypervirulence have been developed, the isolation from marketed food samples of MDR P. aeruginosa carrying a large virulence armamentarium and belonging to a ST which have been qualified as hypervirulent, is of concern.
In summary, MDR P. aeruginosa have been isolated from marketed chicken meat, with the present data strongly suggesting cross contamination during meat manipulation. The present results highlight the risk of acquisition of harmful microorganisms able to cause severe infections through the food chain.
Authors’ Contributions
Y.S., M.J.P., and J.R.: Conceptualization. F.G., K.E., C.A.A., B.R.-B., R.O-L., M.L., C.T.-B., and M.J.P.: Investigation. Y.S. and J.R.: Formal analysis. J.R.: Writing—original draft. Y.S., M.J.P., J.R., F.G., K.E., C.A.A., B.R.-B., R.O-L., M.L., and C.T.-B.: Writing—reviewing and editing.
Footnotes
Acknowledgment
The authors would like to thank Donna Pringle from idiomatic correction.
Funding Information
This study was supported by the Fondo Nacional de Desarrollo Científico, Tecnológico y de Innovación Tecnológica (FONDECYT–Perú) and Universidad Cientifica del Sur within the “Proyecto de Mejoramiento y Ampliación de los Servicios del Sistema Nacional de Ciencia, Tecnología e Innovación Tecnológica” [contract: 08-2019-FONDECYT-BM-INC-INV”]. This study was developed within the frame of the net 221RT0113 from the Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo.
Ethic Statement
The study was approved by the Institutional Review Board of the Universidad Científica del Sur (Code: 053-2019-PRO99). No data of the salespersons or retail stalls was collected, and only generic adscription of samples to local markets was recorded.
Disclosure Statement
No competing financial interests exist.